条石鲷鳔器官早期发育的组织学观察
2010-09-24肖志忠刘清华赵春彦徐世宏马道远
何 滔, 肖志忠 刘清华 李 军 赵春彦, 徐世宏 马道远
(1. 中国科学院 海洋研究所, 山东 青岛 266071; 2. 中国科学院 研究生院, 北京 100049)
条石鲷鳔器官早期发育的组织学观察
何 滔1,2, 肖志忠1, 刘清华1, 李 军1, 赵春彦1,2, 徐世宏1, 马道远1
(1. 中国科学院 海洋研究所, 山东 青岛 266071; 2. 中国科学院 研究生院, 北京 100049)
应用石蜡切片技术和显微测量法, 对条石鲷(Oplegnathus fasciatus)鳔的早期发生、分化和形成过程进行了连续观察。结果显示: 条石鲷为闭鳔鱼类, 在水温为 23.5℃±0.5℃的条件下, 鳔的形成过程可分为鳔原基出现、鳔管形成、鳔腔充气、鳔管退化及气腺形成4个时期。1日龄(孵化后第2天)仔鱼,在食道与肠的交界处分化出鳔原基; 4日龄, 鳔管原基出现; 5日龄, 气腺原基出现; 6日龄, 迷网出现; 8日龄, 鳔管形成; 10日龄, 鳔开始充气; 17日龄, 鳔充气完毕, 鳔管开始退化; 20日龄左右, 鳔管从幽门处完全脱离; 25日龄, 气腺迷网发育完全; 30日龄, 鳔管完全消失, 鳔的发育基本完善。在条石鲷的苗种培育过程中, 鳔器官的发育影响到条石鲷仔鱼的成活率。
条石鲷(Oplegnathus fasciatus); 早期发育; 鳔; 组织学
鳔可以调节鱼体比重、感受大气压力的变化、在某些鱼类中还可以作为呼吸和发声器官。气腺、迷网(即毛细血管网)及鳔管是鳔的主要组成结构,控制着鳔室内气体的生成和排放。根据充气方式及鳔管有无, 可将有鳔鱼分为闭鳔类和管鳔类。鳔在鱼类早期发育中影响着仔稚鱼的成活率[1,2]。
条石鲷(Oplegnathus fasciatus), 属鲈形目(Perciformes)、石鲷科(Oplegnathidae)、石鲷属(Oplegnathus), 主要分布于中国黄海、东海、台湾沿海, 日本北海道以南及韩国以南沿海海域。其肉质极为细致, 口感好, 是一种具有较高食用价值和观赏价值的优良地方海产鱼类[3]。迄今, 在国外人们对条石鲷营养和病理方面[4~9]进行了一些研究; 而在国内则报道了一些条石鲷早期发育的过程[10~16]。王建鑫等[13]对条石鲷成体消化道进行了组织学观察, 但对条石鲷整个早期发育过程的组织学研究还未见报道。在本实验中, 作者从组织学水平对条石鲷鳔的早期发育过程进行了观察, 分析了鳔的发生、分化与死亡高峰的关系, 为死亡高峰的发生机制提供了理论依据。
1 材料与方法
实验所用条石鲷鱼苗于2009年5月~9月取自山东省青岛市金沙滩水产开发有限公司, 采用腹部挤压法分别获得成熟的卵子和精液, 干法人工授精获取受精卵。孵出后的鱼苗培育条件为水温 23.5℃±0.5 ℃, 海水盐度 33±0.5, 日换水量为 1/2, 不间断充气。3~14日龄仔鱼投喂轮虫, 11~40日龄增加投喂卤虫(Artemianauplii)无节幼体, 从25日龄开始给予鱼麋及配合饲料。其中轮虫(Brachionus plicatilis)和卤虫无节幼体投喂前进行营养强化(康克 A, 青岛森淼公司)。
从初孵仔鱼至 100日龄每天连续取样, 取样样本数为40尾。MS-222麻醉, 10尾用于测量生长; 另30尾样品用Bouin’s液室温固定24 h, 70%乙醇保存备用。常规梯度乙醇脱水, 二甲苯透明, 石蜡包埋,分别进行横、纵和水平3个方向的连续切片, 切片厚度 5~8 µm, H.E.染色, 中性树胶封片, Nikon(YS100)显微镜观察和照相。用Photoshop软件进行图像的优化处理, 并以台微尺和目微尺校准图像。
2 结果
2.1 鳔原基出现
2.1.1 初孵仔鱼
未见鳔原基出现(图1-1)。
2.1.2 1日龄仔鱼
全长3.41 mm±0.08 mm, 鳔原基在食道和肠的交界处出现, 为一由间质细胞组成的圆形细胞团, 该细胞团直径为 0.02 mm±0.005mm, 其中部可见微小空腔(图1-2), 其形态与消化管上皮细胞无明显差别。
2.1.3 2日龄仔鱼
全长3.53 mm±0.08 mm, 鳔原基膨大, 间质细胞团向体前、后伸展, 形状由圆形变为椭圆形, 长径0.04 mm±0.01 mm, 短径 0.03 mm±0.01 mm, 但分化并不显著, 细胞团中央的空腔更加明显(图 1-3), 周边的间质细胞清晰可辨, 其外层为结缔组织。
2.1.4 3日龄仔鱼
鳔原基继续膨大、伸长, 长径0.06 mm±0.01 mm,短径 0.04 mm±0.01 mm, 间质细胞团中央空腔增大形成鳔腔, 或称鳔室。在鳔内壁可观察到立方状上皮细胞的排列分布, 其形态与胃原基内壁的矮柱状上皮细胞极其相似。鳔原基周边有较薄的结缔组织, 最外层包裹一层膜状物(鳔膜)将形成鳔的浆膜层。鳔的背部开始出现黑色素细胞(图1-4)。
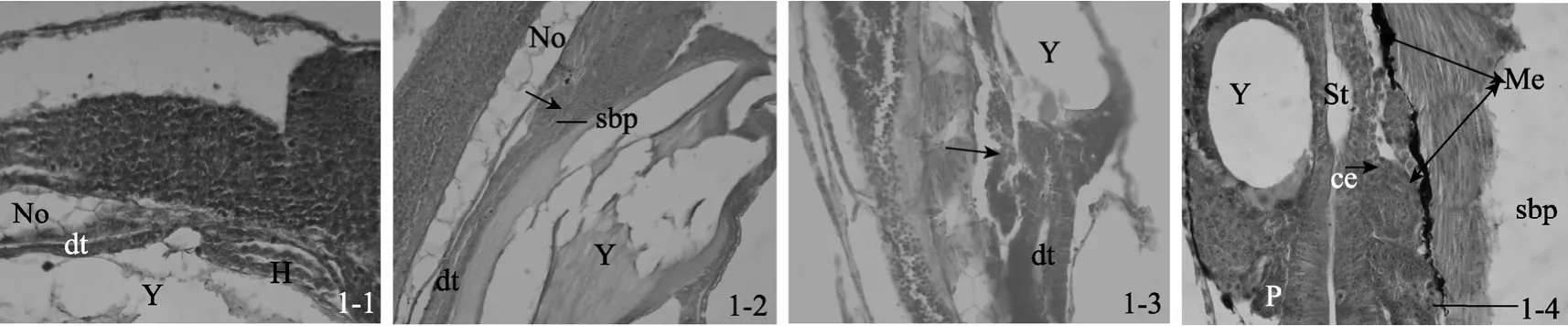
图1 鳔原基出现Fig. 1 Swim bladder primordium
2.2 鳔管形成
2.2.1 4日龄仔鱼
全长 3.56 mm±0.07 mm, 鳔内层有的间质细胞开始分化为背部上皮细胞, 即经扁平化而成的单层扁平细胞。鳔腹部的上皮细胞增厚为2~3层, 上皮细胞层的外层依次为结缔组织层和平滑肌纤维层, 结缔组织与上皮细胞层之间分布有微血管及神经。条石鲷的鳔不分室(即单一鳔室), 在鳔体后部偏左的位置出现鳔管原基, 但其尚未与消化管连通。鳔随仔鱼的生长而逐渐伸长, 鳔腔也随之扩大(图 2-1), 鳔长径0.07 mm±0.01 mm, 短径0.05 mm±0.01 mm。
2.2.2 5日龄仔鱼
仔鱼的摄食量继续增大, 鳔尚未充气。背部上皮细胞继续扁平化, 变薄; 腹部上皮细胞继续增厚, 形成 3~4层; 腹部前下方的多层扁平细胞开始变为立方状的腺细胞, 该处为以后气腺的区域, 细胞间分布有许多的微血管网, 鳔壁外侧为结缔组织和肌纤维层。鳔管从鳔室后部偏左的位置延伸至胃的幽门处(图2-2), 但管腔尚未打通。鳔长径0.08 mm±0.02 mm, 短径 0.06 mm±0.02 mm。
2.2.3 6日龄仔鱼
全长 3.84 mm±0.13 mm, 鳔体背部的黑色素细胞增多, 鳔腔背部的单层扁平上皮细胞特化为膜状,腹前部增厚的上皮细胞褶曲成 3~4层的多层结构,呈波浪型, 其中的立方状腺细胞群与微血管网交织在一起, 形成鳔的气腺组织, 并行使分泌、充气的生理功能(图2-3)。鳔长径0.09 mm±0.02 mm, 短径0.05 mm。
2.2.4 8日龄仔鱼
全长4.19 mm±0.16 mm, 鳔腔继续伸展拉长, 呈长椭圆形, 后端略尖细, 前端圆钝, 鳔长径 0.17 mm±0.04 mm, 短径0.09 mm±0.01 mm。鳔背部的黑色素细胞向着腹部延伸。鳔管管腔形成,并连通鳔室和幽门, 其管壁结构呈多元化, 靠近鳔室的部分由单层扁平上皮、结缔组织、肌肉层、外层被膜组成, 而靠近胃体的部分则与胃壁的结构相似, 由单层柱状上皮、结缔组织、肌肉层、外层被膜组成。此时, 在鳔腹前部的气腺区域呈现复杂的多层盘曲结构(立方状的腺细胞群与微血管网), 几乎占满整个鳔腔。气腺发育完善, 迷网形成, 毛细血管中可见少量血细胞(图 2-4)。

图2 鳔管形成Fig. 2 Development of the pneumatic duct
2.3 鳔腔充气
2.3.1 10日龄仔鱼
全长4.77 mm±0.29 mm, 鳔开始充气, 呈椭圆形,鳔长径0.30 mm±0.05 mm, 短径0.13 mm±0.02 mm。此时仔鱼的游泳能力增强, 摄食积极, 食量增加, 仔鱼的运动呈集群和趋光性。鳔腹部的气腺立方上皮细胞增厚为2~3层, 腺细胞核较大, 染色时着色较深,核位于细胞基部(图3-1)。

图3 鳔腔充气Fig. 3 Swim bladder inflation
2.3.2 14日龄仔鱼
全长6.18 mm±0.48 mm, 鳔长径0.36 mm±0.06 mm, 短径0.15 mm±0.03 mm, 鳔的腹部开始有黑色素细胞的分布, 迷网血管及血细胞数量增多。鳔的充气继续进行, 而一些鳔未充气的仔鱼, 其鳔腔中充满大量褶曲的上皮细胞(图3-2), 则很快死亡。
2.4 鳔管退化及气腺形成
2.4.1 17日龄仔鱼
全长7.19 mm±0.81 mm, 随着鳔的充气, 鳔体积进一步膨大, 鳔长径 0.38 mm±0.07 mm, 短径 0.17 mm±0.03 mm。此时多数仔鱼的鳔中已充满气体。鳔管开始退化, 管腔填充关闭, 鳔管首先从胃幽门处脱离开来(图4-1)。整个鳔体均有黑色素细胞的分布,色素密集, 气腺及迷网血管中可见大量血细胞, 鳔受到迷走神经和交感神经腹腔神经节的双重调节,在迷网微血管内碳酸苷酶和乳酸的参与下, O2和CO2通过血细胞的对流交换进入气腺, 并由气腺进入鳔腔。
2.4.2 20日龄仔鱼
全长9.49 mm±1.18 mm, 仔鱼的鳔体继续增大,鳔长径0.41 mm±0.07 mm, 短径0.18 mm±0.05 mm。其鳔腹外部的黑色素细胞逐渐消失, 背部的黑色素细胞分布则更加浓密。气腺继续增厚, 迷网发达。鳔管已完全从幽门处脱离, 但在鳔室后部还有所残余(图 4-2)。
2.4.3 25日龄稚鱼
全长12.34 mm±1.19 mm, 鳔长径0.50 mm±0.12 mm, 短径0.26 mm±0.06 mm。鳔体背部的黑色素细胞更加浓密, 气腺由6~7层立方细胞组成, 通过毛细血管与迷网相连, 迷网中有大量毛细血管, 相互交错呈网状结构(图 4-3)。至此, 气腺与迷网基本发育完善, 气体通过气腺和迷网的相互调节来控制鳔腔的大小, 并决定稚鱼的沉浮或停滞。
2.4.4 30日龄稚鱼
全长19.45 mm±1.61 mm, 鳔管完全消失, 鳔发育成熟。鳔不分室, 由鳔腔向外可分为鳔腔-内壁上皮细胞层-结缔组织(疏松层和致密层)-环状肌肉层(气腺、迷网)-外层被膜(图 4-4)。此后, 鳔随着稚鱼的生长而沿鱼体轴方向拉长伸展。
2.4.5 40日龄幼鱼
全长28.16 mm±4.09 mm, 鳔发育成熟, 位于消化道背部与脊椎骨下方之间, 鳔外侧具一层较薄的银白色腹膜。

图4 鳔管退化Fig. 4 Regression of the pneumatic duct
3 讨论
3.1 鱼类鳔早期发育的比较
本实验的观察显示, 条石鲷属于闭鳔类, 1日龄(孵化后第 2天)即出现鳔原基, 这与大菱鲆(Scophthatmus maximus)[17]、短盖巨脂鲤(Colossoma brachyopomum)[18]、南方鲶(Silurus meridionalis Chen)、大鳍 (Mystus macropterus)[19]鳔原基出现的时间相同, 而真鲷(Pagrosomus major)[20]、条纹婢(Latris lineata)[1]在孵化后第 1天(初孵仔鱼)即出现鳔原基, 半滑舌鳎(Cynoglossus semilaevis)[21]则在孵化后第3天(2日龄)出现鳔原基。
到10日龄, 条石鲷仔鱼的鳔开始充气, 到17日龄充气完毕, 充气间隔为 7 d; 真鲷[20]从孵化后第 9天开始充气, 到孵化后第 12天左右充气完成, 充气间隔为 3天; 短盖巨脂鲤[18]从 4~5日龄开始充气,到11日龄充气完毕, 充气间隔为6~7天; 大菱鲆[17]从孵化后第5天开始充气, 到第19天充气完毕, 充气间隔则达14 d。
条石鲷仔鱼的鳔管开口于胃幽门处, 与其相似的有太平洋油鲽(Solea solea)[22], 其鳔管开口于胃后端背部; 而南方鲶、大鳍[19]、真鲷[20]、大菱鲆[17]仔鱼的鳔管则开口于胃贲门处或胃的前端。
从 17日龄开始, 条石鲷仔鱼的鳔管开始退化,到30日鳔管消失。真鲷[20]仔鱼的鳔管在孵化后第12天就已退化; 大菱鲆[17]仔鱼的鳔管则在19日龄左右开始封闭; 30日龄鳔管完全消失标志着条石鲷鳔器官发育的基本完成, 这与真鲷[20]的基本一致。
3.2 鳔器官对仔鱼生长发育的影响
本实验中, 观察发现 1日龄仔鱼在水体中呈倒垂状或与水层成 45°斜垂在水体表层, 这可能正与此时鳔原基的出现相关。因为卵黄囊和油球逐渐被吸收, 而此时仔鱼的鳔发育并不完善, 尚不能实施其生理功能, 所以使得条石鲷仔鱼在水域中呈停滞状态并倒悬。蔡泽平等[23]在初孵真鲷仔鱼中也发现类似鱼体的斜悬现象, 但未做深入的分析。总之, 此阶段仔鱼的倒悬原因需进一步的实验论证。
鳔的充气对于仔稚鱼的摄食具有重要的意义,因为仔鱼在摄食的时候, 鳔充气良好较充气不足的消耗的能量较少, 故摄食能力较高, 存活率也会相应地较高。一般来说, 鳔的充气通常是在卵黄即将吸收完毕和外源性摄食开始的时候进行[2]。条石鲷仔鱼在5日龄的时候, 卵黄囊和油球已完全被吸收, 而鳔在10日龄才开始充气, 之间相隔5 d; 真鲷仔鱼的卵黄则在6日龄的时候被完全吸收[23], 在8日龄鳔就开始充气[20], 之间只相隔2 d; 而半滑舌鳎[21]和大菱鲆[17]的仔鱼则分别在4日龄、5日龄卵黄囊完全消失的同时开始鳔的充气; 相比较而言, 条石鲷从卵黄吸收完全到鳔开始充气的时间间隔较长, 故这段时间的存活率相应地较低。在生产育苗中作者也发现在12~18日龄左右, 条石鲷仔鱼出现了大量死亡的现象, 这可能正与条石鲷鳔开始充气的时间相对较晚有关, 仔鱼摄食能力的相对较低导致了其大量死亡。同时组织学的观察也表明, 从10日龄鳔开始充气到17日龄充气完毕, 这段时间正是条石鲷仔鱼鳔充气的关键时期, 所以这段时间鳔的不正常充气也可能是仔鱼大量死亡的原因。
3.3 外界因子的影响
温度[24]、光照强度、光照周期[25]、盐度、水体充气速率[26]、水体浑浊度[27]等环境因子以及高度不饱和脂肪酸[28]、甲状腺素[29]等营养因子影响着鳔的充气。Trotter等[1,24,25,30]对条纹婢的系列研究发现,外界因子如温度、光照对鳔的起始充气率和充气后存活率有显著影响, 通过对温度、光照的调控可以明显地提高鳔的起始充气率以及充气后存活率。
本实验中, 作者将温度、光照、pH等环境因子保持了恒定, 但在条石鲷仔鱼充气阶段(12~18日龄左右)出现了仔鱼大量死亡的现象, 这很可能就与此时的外界因子未达到条石鲷仔鱼所需的最佳水平有关, 当然也可能与鱼体自身的某些遗传特性相关。在今后的实验中, 应在不同的环境、营养因子水平研究鳔的发育状况, 以期通过对外界因子的人为控制来提高鳔发育的完善性, 并提高仔稚鱼的成活率。
[1] Trotter A J, Pankhurst P M, Battaglene S C,et al.Morphology of swim bladder development in hatchery-reared striped trumpeterLatris lineata[J]. Journal of Applied Ichthyology, 2004, 20: 395-401.
[2] 门强, 雷霁霖, 武云飞. 鳔器官的发育对人工培育育苗的影响[J]. 海洋水产研究, 2003, 24(1): 81-84.
[3] 朱元鼎. 福建鱼类志(下卷)[M]. 福州: 福建科学技术出版社, 1985. 254-255.
[4] Nam Y K, Cho Y S, Choi B N,et al. Alteration of antioxidant enzymes at the mRNA level during short-term starvation of rockbreamOplegnathus fasciatus[J].Fisheries Science, 2005, 71: 1 385-1 387.
[5] Cho Y S, Choi B N, Kim K H,et al. Differential expression of Cu Zn superoxide dismutase mRNA during exposures to heavy metals in rockbreamOplegnathusfasciatus[J]. Aquaculture, 2006, 253: 667-679.
[6] Tachibana K, Yagi M, Hara K,et al. Effects of feeding of β-carotene-supplemented rotifers on survival and lymphocyte proliferation reaction of fish larvae Japanese parrotfish (Oplegnathus fasciatus) and Spotted parrotfish(Oplegnathus punctatus)preliminary trials[J].Hydrobiologia, 1997, 358: 313-316.
[7] Wang X J, Kim K W, Bai S C,et al. Effects of the different levels of dietary vitamin C on growth and tissue ascorbic acid changes in parrot fish (Oplegnathus fasciatus)[J]. Aquaculture, 2003, 215: 203-211.
[8] Jung S J, Oh M J. Iridovirus-like infection associated with high mortalities of striped beakperch,Oplegnathus fasciatus(Temminck et Schlegel), in southern coastal areas of the Korean peninsula[J]. Journal of Fish Diseases, 2000, 23: 223-226.
[9] Choi S K, Kwon S R, Nam Y K,et al. Organ distribution of red sea bream iridovirus(RSIV) DNA in asymptomatic yearling and fingerling rock bream(Oplegnathus fasciatus) and effects of water temperature on transition of RSIV into acute phase[J]. Aquaculture, 2006, 256: 23-26.
[10] 常抗美, 毛建平, 吴剑锋, 等. 条石鲷胚胎及仔稚鱼的发育[J]. 上海水产大学学报, 2005, 14(4): 401-405.
[11] 辛俭, 薛利建, 毛国民, 等. 条石鲷的胚胎发育观察[J].浙江海洋学院学报(自然科学版), 2005, 24(1):31-36.
[12] 刘朝阳, 孙晓庆. 条石鲷的生物学特性及增养殖前景[J]. 齐鲁渔业, 2006, 23(7): 6.
[13] 王建鑫, 石戈, 李鹏, 等. 条石鲷消化道的形态学和组织学[J]. 水产学报, 2006, 30(5): 618-626.
[14] 全汉锋, 肖治中. 条石鲷人工繁育技术研究[J]. 台湾海峡, 2007, 26(2): 295-300.
[15] 柳学周, 徐永江, 王妍妍, 等. 条石鲷的早期生长发育特征[J]. 动物学报, 2008, 54(2): 332-341.
[16] 肖志忠, 郑炯, 于道德, 等. 条石鲷早期发育的形态特征[J]. 海洋科学, 2008, 32(3): 25-30.
[17] 门强, 雷霁霖. 大菱鲆鳔器官发育的形态学与组织学特征[J]. 中国水产科学, 2003, 10(2): 111-114.
[18] 郭恩棉, 张艳萍, 王鑫. 短盖巨脂鲤鳃、伪鳃和鳔胚后发育学研究[J]. 海洋湖沼通报, 2005, 3: 31-37.
[19] 龙祥平, 王德寿, 田怀军. 南方鲶、大鳍 鳔发育的组织学研究[J]. 水产学报, 1993, 17(4): 340-343.
[20] 马爱军, 雷霁霖, 马英杰, 等. 真鲷仔稚鱼鳔发育与分化研究[J]. 青岛海洋大学学报, 1998, 28(4):593-598.
[21] 蔡文超, 柳学周, 马学坤, 等. 半滑舌鳎早期发育阶段鳔和冠状幼鳍的生长发育规律研究[J]. 海洋水产研究, 2006, 27(2): 94-98.
[22] Boulhic M, Gabaudan J. Histological study of the organogenesis of the digestive system and swim bladder of the Dover sole,Solea solea(Linnaeus 1758)[J].Aquaculture, 1992, 102: 373-396.
[23] 蔡泽平, 蔡国雄, 何伟宏. 真鲷早期发育观察[J]. 南海研究与开发, 1990, 3: 43-48.
[24] Trotter A J, Pankhurst P M, Morehead D T,et al. Effects of temperature on initial swim bladder inflation and related development in cultured striped trumpeter(Latris lineata)larvae[J]. Aquaculture, 2003, 221:141-156.
[25] Trotter A J, Battaglene S C, Pankhurst P M,et al.Effects of photoperiod and light intensity on initial swim bladder inflation, growth and post-inflation viability in cultured striped trumpeter (Latris lineata) larvae[J].Aquaculture, 2003, 224: 141-158.
[26] Battaglene S C, Talbot R B. Effect of salinity and aeration on survival of and initial swim bladder inflation in larval Australian bass[J]. Prog Fish-Cult, 1993, 55:35-39.
[27] Rieger P W, Summerfelt R C. The influence of turbidity on larval walleye,Stizostedion vitreum, behavior and development in tank culture[J]. Aquaculture, 1997,159: 19-32.
[28] Chikara Kitajima, Takeshi Watanabe, Yasuo Tsukashimaet al. Lordotic deformation and abnormal development of swimbladder in some hatchery-reared marine physoclistous fish in Japan [J]. World Aquaculture Society, 1994, 25(1): 64-77.
[29] 赵维信, 贾江. 鳙、团头鲂和短盖巨脂鲤早期发育阶段甲状腺素含量变化研究[J]. 水产学报, 1997, 21(2):120-127.
[30] Trotter A J, Pankhurst P M, Hart P R,et al.Swim bladder malformation in hatchery-reared striped trumpeterLatris lineate(Latridae)[J]. Aquaculture, 2001, 198:41-54.
Received: Jun., 23, 2010
Key words:Japanese parrotfish; early development; swim bladder; histology
Abstract:Based on the histological observations under optical microscope, we found that the Japanese parrotfish belonged to physoclistous fish. The development of swim bladder in Japanese parrotfish included four stages: generation of anlage, formation of pneumatic duct, initial inflation, degeneration of the pneumatic duct, and formation of gas gland. At 23.5 ± 0.5 ℃, the primordial swim bladder was first observed at 1 DAH. The primordium of pneumatic duct, gas gland and rete mirabile were present at 4, 5, and 6 DAH, respectively. The initial inflation occurred at 10 DAH, and finished at 17 DAH. The pneumatic duct began to degenerate from pylorus at 17 DAH and disappeared entirely at 30 DAH. The swim bladder was functional from 30 DAH and onward.
(本文编辑:谭雪静)
Histological observation of swim bladder during early development stage of Japanese parrotfish(Oplegnathus fasciatus Temminck et Schlegel 1844)
HE Tao1,2, XIAO Zhi-zhong1, LIU Qing-hua1, LI Jun1, ZHAO Chun-yan1,2, XU Shi-hong1, MA Dao-yuan1
(1. Institute of Oceanology, the Chinese Academy of Sciences, Qingdao 266071, China; 2. The Graduate School of the Chinese Academy of Sciences, Beijing 100049, China)
Q954.61
A
1000-3096(2010)12-0008-06
2010-06-23;
2010-07-22
中国科学院创新性前沿项目(KSCX2-YW-N-47-08); 国家农业科技成果转化资金项目(05EFN2166000453); 青岛市科技发展计划项目(05-1-HY-79)
何滔(1985-), 男, 重庆万州人, 博士研究生, 主要从事养殖生物学研究; 李军, 通信作者, 研究员, E-mail: junli@ms.qdio.ac.cn
