温度及土壤含水量对冬小麦生长习性及返青率的影响
2010-09-20孟健男李卓夫
于 晶,孟健男,曾 俨,苍 晶*,李卓夫
(1.东北农业大学生命科学学院,哈尔滨 150030;2.东北农业大学农学院,哈尔滨 150030)
低温对植物的伤害表现为冷害和冻害两种类型,冷害是0℃以上低温对植物的伤害,包括直接破坏细胞原生质活性的直接伤害和引起细胞代谢失常的间接伤害,冻害是0℃以下低温使植物组织内结冰而引起的伤害,包括细胞内结冰和细胞间隙结冰伤害[1]。不同植物种类、同种植物的不同品种对低温的抗性不同,在可承受的低温下植物体内会发生许多生理生化变化[2-3]及诱导抗寒基因或抗冻蛋白的形成来适应低温逆境[4-7],但随着低温的不断下降及低温时间的延长,植物便会死亡,因此,通过植物受低温胁迫后恢复生长的能力(返青率)可以直接来判断其抗寒能力[8]。
2007年,冬小麦品种东农冬麦1号在黑龙江省高寒地区育成、并通过了品种审定,该品种抗寒性极强,可以在黑龙江省等寒地(最低温度-30℃)存活,但是,由于黑龙江省不仅冬季严寒、风大,而且降雪量年际间不平衡,若雪少则易造成冬季麦苗裸露田间,被风吹干而死亡,如果遇秋冬和返青期土壤含水量不适宜(过高或过低),就会在很大程度上影响返青率。为确保东农冬麦1号能够安全越冬,有必要对其土壤含水量和低温双重逆境胁迫进行综合深入研究。目前对东农冬麦1号的研究主要集中在对低温的抗性上[9-11],对不同胁迫之间的相互关系以及交叉适应的机制还所知甚少。本研究通过比较不同抗寒品种东农冬麦1号、济麦22(对照)品种在不同土壤含水量及低温逆境胁迫下返青率的情况,探讨植物对不同胁迫交叉适应的可能机制,确定春季田间最适返青的土壤含水量,为指导大田冬小麦封冻前及春季返青前浇灌提供理论依据。
1 材料与方法
1.1 试验材料与种植
供试冬小麦材料为东农冬麦1号(D)和济麦22(J),由东北农业大学小麦育种室提供。种子精选后于2008年9月9日室外盆栽播种,盆栽用土取自东北农业大学实验田,黑土。播深5 cm,株距1 cm,坐水种植,施基肥量与正常大田常规种植一致。盆栽材料3种土壤含水量(30%~35%,25%~30%,20%~25%)处理,每个处理10盆。3 d测定一次土壤(分蘖节附近)含水量,使土壤水分一直保持在上述3个含量范围内。
1.2 试验方法
1.2.1 幼苗外部形态及生长习性
越冬期不同温度下观察幼苗外部形态,并对试验材料的幼苗生长习性进行目测评定。
1.2.2 返青率调查
分别记录小麦盆栽材料越冬期温度稳定5 d降至-10~-15℃、-15~-20℃、-20~-25℃和-25~-30℃后,在昼夜温度15℃/10℃恢复培养及次年春季缓慢升温时的盆栽返青率,并以同时期大田种植材料的返青率为参照。
1.2.3 数据记录及统计分析
采用Excel 2003完成数据处理,DPS 3.01软件进行方差分析和相关性分析。
2 结果与分析
2.1 麦苗外部形态及生长习性
小麦抗寒性与麦苗的生长状态有关。匍匐型(叶片全部匍匐于地面)是抗寒性强的类型;半匍匐型(叶片尖端与地面接触,叶身离开地面)具有中等抗寒性;半直立型(叶片全部离开地面,但不完全直立)具有较弱的抗寒性;直立型(叶片全部直立)是抗寒性弱的类型。两个冬小麦品种越冬期生长状态有很大的差异。东农冬麦1号叶片全部匍匐于地面,济麦22叶片尖部着地,叶身直立(见图版Ⅰ)。两个品种均2~3个分蘖,在低温-10~-15℃时,叶片均呈深绿色,两者直观上无差异;在低温-15~-20℃时东农冬麦1号叶片仅叶尖部略黄,而济麦22叶片明显黄化;在低温-20~-25℃及-25~-30℃时东农冬麦1号仅叶片尖端干枯,而济麦22叶片几乎全部黄枯。
2.2 返青率
在气温连续5 d通过-10~-15℃、-15~-20℃、-20~-25℃及-25~-30℃时,分别将室外盆栽材料转移至室内(15℃/10℃)恢复培养,调查返青情况,并于次年春季调查缓慢升温的盆栽及大田返青率(见表1)。恢复生长15 d后的麦苗状态见图版Ⅱ~Ⅳ。

图版Ⅰ 越冬期东农冬麦1号(D)与济麦22(J)麦苗生长状态PlateⅠ Wheat growth state of Dongnongdongmai1 and Jimai22 in wintering
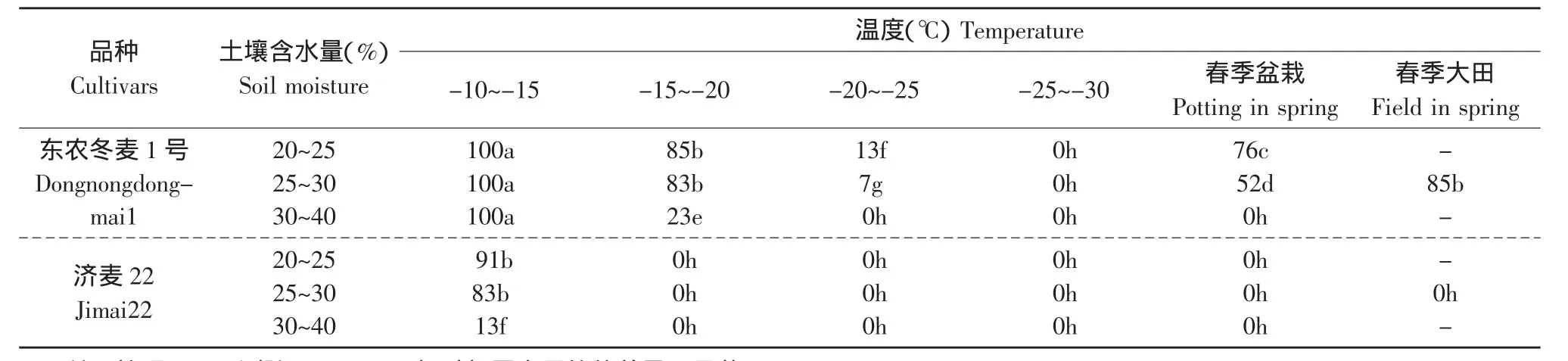
表1 东农冬麦1号与济麦22的返青率Table1 Overwinter survival rate of Dongnongdongmai1 and Jimai22

图版Ⅱ 东农冬麦1号(D)与济麦22(J)恢复生长15 d的麦苗状态(-10~-15℃)PlateⅡ Wheat state of Dongnongdongmai1 and Jimai22 after recovery growth 15 d(-10~-15℃)

图版 Ⅲ 东农冬麦1号(D)与济麦22(J)恢复生长15 d的麦苗状态(-15~-20℃)PlateⅢ Wheat state of Dongnongdongmai1 and Jimai22 after recovery growth 15 d(-15~-20℃)

图版 Ⅳ 东农冬麦1号(D)与济麦22(J)恢复生长15 d的麦苗状态(-20~-25℃及-25~-30℃)PlateⅣ Wheat state of Dongnongdongmai1 and Jimai22 after recovery growth 15 d(-20~-25℃and-25~-30℃)
-10~-15℃低温后的麦苗在昼夜15℃/10℃温度下恢复生长15 d时,两个材料均已返青,不同的土壤含水量麦苗返青情况有很大差异。土壤含水量高,返青苗长势弱,济麦22烂根极为严重,存活率低;土壤含水量正常或偏低时,返青麦苗长势好,根壮叶茂(见图版V-a)。从整体来看,东农冬麦1号的长势在3种土壤含水量下均好于济麦22(见图版Ⅱ)。-15~-20℃低温后的麦苗在15℃/10℃温度下恢复生长15 d时,不论土壤含水量高低,济麦22均烂根,无一返青,而东农冬麦1号在含水量适中及偏低时能够返青,且低含水量处理的返青率高于适中含水量处理(见图版Ⅲ),其根系生长旺盛(见图版V-b),但地上部分长势比-10~-15℃降温下生长的麦苗慢。
-20~-25℃及-25~-30℃低温后的麦苗在15℃/10℃温度下恢复生长15 d时东农冬麦1号和济麦22均未返青(见图版Ⅳ),且各含水量处理的济麦22均烂根,而东农冬麦1号无烂根现象(见图版V-c),约30 d后东农冬麦1号开始返青。麦苗越过冬季后,在次年春季缓慢升温条件下,济麦22均未返青,东农冬麦1号正常返青。试验表明,小麦越冬期经同样低温后,经缓慢升温过程的返青率要显著高于突然增至高温的返青率;随着温度的降低,返青时间逐渐延长,返青率下降。综上所述,无论土壤含水量在什么范围内,济麦22越冬的极限温度都在-15℃左右,而东农冬麦1号在-30℃左右的越冬条件下仍可以正常返青。相关性分析表明,温度与返青率呈显著正相关,相关系数为0.91;土壤含水量与返青率呈负相关,相关系数为-0.18。

图版V 不同温度及不同含水量下东农冬麦1号(D)与济麦22(J)的比较PlateV Comparison of Dongnongdongmai1 and Jimai22 in different temperature and different soil moisture
3 讨论与结论
小麦能否安全越冬主要取决于品种(即基因型)的抗寒性,但也受外界环境及栽培措施等其他因素影响。对于同一小麦品种来说,抗寒性与其播期、播量及播深有关,还与光照、温度、土质和墒情、肥料的施用种类及多少有关[12]。小麦品种的抗寒性可通过直接目测越冬期小麦形态而初步测定,即抗寒性强的品种植株匍匐于地面生长,抗寒性弱的品种植株直立于地面生长;也可以通过第二年返青情况直接判定,即经过严冬后存活的小麦比例多少。本试验通过外部形态及返青率同时证实了东农冬麦1号的抗寒性极强,济麦22的抗寒性中等,两种方法互相验证,结果非常可靠。土壤含水量影响小麦的返青率。目前,对麦田含水量与小麦抗寒性关系的研究尚不系统。本试验通过盆栽控制土壤含水量的方法初步证明:土壤水分超过30%会导致返青期的小麦严重烂根;土壤含水量中等(25%左右)或偏低(20%左右),有利于小麦返青,根系生长旺盛;但土壤水分偏低时,麦苗长势弱于土壤含水量中等时的情况;但土壤含水量过低,则麦苗会旱死。至于土壤水分含量低到什么程度会导致小麦不能返青,还需进一步研究。此外,小麦在越冬期经历的温度不同对返青的影响也很大。本试验通过土壤含水量与温度的联合分析表明,在不同土壤含水量下,小麦越冬后的返青率均随温度的不断降低而逐渐下降,且返青时间相应延长。关于小麦返青的土壤临界水分、气温以及它们之间的关系正在进一步探讨。
[1]李合生.现代植物生理学[M].2版.北京:高等教育出版社,2006.
[2]Gana J A,Sutton F,Kenefick D G.cDNA structure and expression patterns of a low-temperature-specific wheat gene tacr7[J].Plant Mol Biol,1997,34:643-650.
[3]Minami A,Nagao M,Ikegami K,et al.Cold acclimation in bryophytes:low-temperature-induced freezing tolerance in Physcomitrella patens is associated with increases in expression levels of stress-related genes but not with increase in level of endogenous abscisic acid[J].Planta,2005,220:414-423.
[4]Cabane M,Calvet P,Vincens P,et al.Characterization of chillingacclimation-related proteins in soybean and identification of one as amember of the heat shock protein HSP70 family[J].Planta,1993,190(3):346-353.
[5]Guy C L,Haskell D.Detection of polypeptides associated with the cold acclimation process in spinach[J].Electophoresis,1988,9(11):787-796.
[6]Meza-Basso L,Alberdi M,Raynal M,et al.Changes in protein synthesis in rapeseed(Brassica napus)seedling during a low temperature treatment[J].Plant Physiol,1986,82(3):733-738.
[7]李晓毓.低温胁迫下菊花蛋白质组的双向电泳分析及质谱鉴定[D].贵阳:贵州大学,2006.
[8]王静,魏小红,龙瑞军.植物抗寒机制的研究方法与进展[J].甘肃科技纵横,2004,33(6):72-73.
[9]王晓楠,付连双,李卓夫,等.低温驯化及封冻后不同抗寒性小麦品种的形态建成及生理基础分析[J].作物学报,2009,35(7):1313-1319.
[10]于晶,张林,苍晶,等.不同抗寒性冬小麦分蘖节低温诱导蛋白比较研究[J].应用生态学报,2009,20(5):1092-1098.
[11]于晶,张林,崔红,等.高寒地区冬小麦东农冬麦1号越冬前的生理生化特性[J].作物学报,2008,34(11):2019-2025.
[12]于晶,张林,苍晶,等.冬小麦抗寒性研究进展[J].东北农业大学学报,2008,39(11):123-127.
