中国海五种群三疣梭子蟹的形态差异分析
2010-09-05董志国李晓英阎斌伦高焕吴旭干成永旭孙效文
董志国,李晓英,阎斌伦,高焕,吴旭干,成永旭,孙效文
(1. 淮海工学院 江苏省海洋生物技术重点实验室,江苏 连云港 222005;2. 上海海洋大学水产与生命学院,上海 201306;3.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
中国海五种群三疣梭子蟹的形态差异分析
董志国1,2,3,李晓英1,阎斌伦1,高焕1,吴旭干2,成永旭2,孙效文3
(1. 淮海工学院 江苏省海洋生物技术重点实验室,江苏 连云港 222005;2. 上海海洋大学水产与生命学院,上海 201306;3.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
运用三种多元分析方法,即聚类分析、主成分分析和判别分析,通过测量三疣梭子蟹23个形态学性状参数,对中国海大连海区、东营海区、连云港海区、舟山海区和湛江海区5个种群三疣梭子蟹的形态差异进行了比较研究。聚类分析结果表明,连云港种群和东营种群形态最为相似(形态距离0.07),大连种群与东营种群、大连种群(dl)与舟山种群(zs)的趋异程度逐渐增加,形态距离分别为0.08和0.09,而湛江种群的趋异程度最大,与其余4种群的形态距离达0.15以上。主成分分析构建了3个主成分,其贡献率:主成分1为27.39%,主成分2为15.23%,主成分3为10.27%,累积贡献率为52.89%。在第1主成分中,比例性状ICM/CL、BCW/CL、PWC/CL和SW/CL的影响比较大。判别分析结果表明,5个海区种群之间的形态差异极显著(P<0.01),通过建立5个地理种群的判别函数,结果判别准确率P1为63.64%~94.44%, P2为67.74%~92.86%,综合判别率为82.24%,因此,该判别方程具有较高的判别效果。
中国海;三疣梭子蟹;形态差异;多元分析
三疣梭子蟹(Portunus trituberculatus),俗称海蟹或大蟹,属于甲壳纲,十足目,梭子蟹科。广泛分布在中国南北沿海,是中国大型海洋经济蟹类,是沿海重要的渔业资源。生活在中国近海不同海区的三疣梭子蟹由于生活环境的不尽相同,水文因素的差异以及各海区地理位置的不同造成三疣梭子蟹分属于各个不同的种群。近年来,国内外有关三疣梭子蟹的增养殖生态[1-3]、繁殖发育[4-6]和资源恢复和人工放流[7-10]等方面的研究较多,而对三疣梭子蟹在分子水平上的研究较少,仅见郭天慧等(2004)对三疣梭子蟹线粒体DNA 16S rRNA和COI基因片段的初步研究[11]以及日本东京大学Mitsugu M. Yamauchi 等(2003)对线粒体基因组的研究[12]。而关于三疣梭子蟹群体遗传方面的研究国内外均少有研究报道,仅见高宝全等(2007)对我国北方四个三疣梭子蟹的群体研究[13],特别是中国主要海区三疣梭子蟹的种群遗传结构和种质资源状况却仍有待深入研究。本文运用形态学分析方法,以聚类分析、主成分分析、判别分析三种多元分析方法,对我国四大海区5个不同地理种群的三疣梭子蟹进行综合分析,找出差异较大的形态特征,并建立鉴别方法,为三疣梭子蟹种质资源的保护和利用,以及生产实践提供科学依据。
1 材料和方法
1.1 实验材料
试验材料均取自天然海区,分别是大连海区(dl)、东营海区(dy)、连云港海区(lyg)、舟山海区(zs)和湛江海区(zj)这5个海区的雌性三疣梭子蟹,各种群取样时间、地点和数目见表1,所得到的样本快速低温运送到实验室并在第一时间对其进行实验数据的测量。
1.2 测量方法
采用游标卡尺、分规、直尺等测量工具,精确到0.2 mm,测量方法参照黎中宝(2004)[14]和Clive P.Keenan(1998)[15]。选取三疣梭子蟹背甲、前额裂片、腹甲、步足和螯肢部位,测量参数为23个可量性状,共测得3 496个形态学数据。

表1 三疣梭子蟹样本生物学数据Tab. 1 Samples information of P. trituberculatus
1.3 数据处理
使用STASTISTICA 99’edition软件进行数据处理,采用聚类分析,主成分分析和判别分析3种多元分析方法对5个地理种群进行形态差异分析。
1.3.1 聚类分析 为消除蟹体规格大小对参数值的影响,参照Brzeski(1988)[16]先将每只蟹的所有参数分别除以背甲长(CL)予以校正,得出22个形态学比例性状,再分别求出各组样本每个参数校正值的平均值,用平均校正值进行聚类分析,采用欧氏距离的最短距离系统聚类法(张尧庭和方开泰,1982)[17]。
1.3.2 主成分分析 为将多个指标作为小数指标的统计方法,所有数值处理同聚类分析方法先进行校正处理,从22个比例性状中通过计算机程序得出3个综合性指标,即互不关联的3个主成分。
1.3.3 判别分析 对所有样本进行逐个判别。判别准确率的计算公式为[18]:
判别准确率P1(%)=判别正确的数目/实测数目×100
判别准确率P2(%)=判别正确的数目/判别数目×100
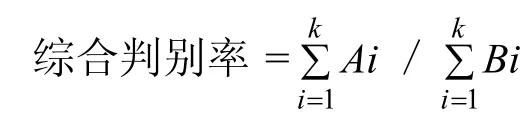
其中,Ai为第i个种群判别正确的梭子蟹数目,Bi为第i个种群实际判别的梭子蟹数目,k为种群数。
2 结果分析
2.1 聚类分析
图1为5个地理群体三疣梭子蟹所有样本的22个比例性状的形态聚类图,结果显示5个群体可以分为二组,一组为大连、东营、连云港、舟山群体,而另一组为湛江群体。连云港种群(lyg)与东营种群(dy)形态距离最小,为0.07,形态最为接近,大连种群(dl)与连云港种群(zs)和舟山种群这两种群的趋异程度逐渐增加,形态距离为0.09,舟山与连云港和东营的形态距离均为0.11,而湛江种群(zj)与上述4个群体的形态距离最大,均在0.15以上,特别是湛江群体和大连群体形态距离达到0.18,趋异程度最大。
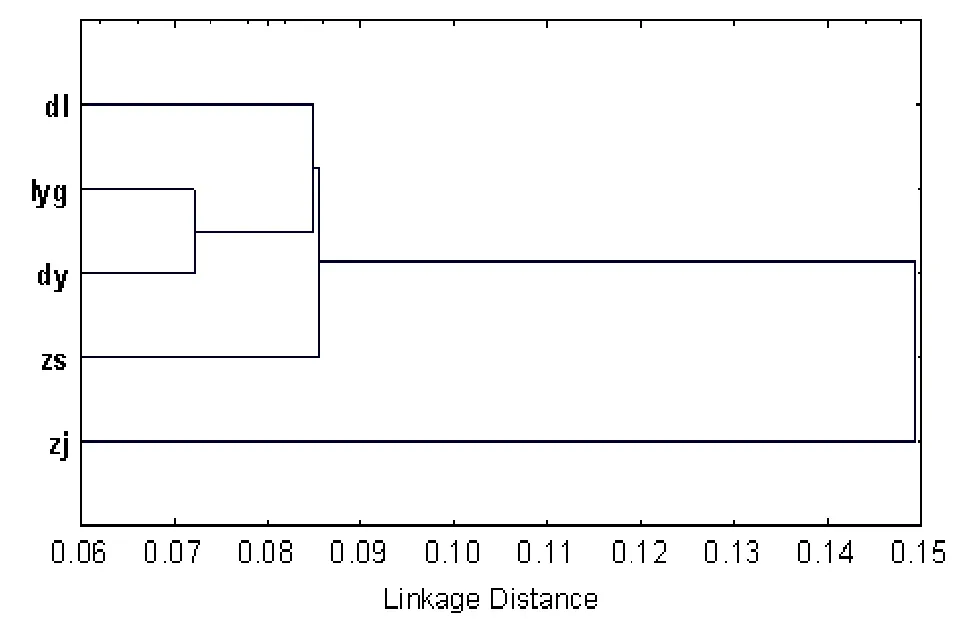
图1 5个地理种群三疣梭子蟹聚类分析图Fig. 1 Diagram of cluster analysis of five populations of P. trituberculatus
2.2 主成分分析
主成分分析结果见表2。总方差的贡献率分别为,主成分1为27.39%,主成分2为15.23%,主成分3为10.27%,累积贡献率为52.89%,三个主成分的累积贡献率较低,说明用几个相互独立的因子难以概括不同群体之间的形态差异。在第1主成分中,比例性状ICW/CL、8CW/CL、PWC/CL和SW/CL的影响比较大。图2为5个地理种群三疣梭子蟹的第1、2主成分的散布图。由图2可以看出,除了湛江种群之外的其他四个种群有较多的重叠区,说明这四个种群在形态上很相近,而湛江种群相对于其他种群在形态上差异较大,与聚类分析的结果相一致。

表2 5个地理种群三疣梭子蟹3个主成分的贡献率及负荷值Tab. 2 Loadings of three principal components of five populations of P.trituberculatus

图2 5个地理种群三疣梭子蟹第1、2主成分散布图Fig. 2 Scatter diagram for the first and the second principal components of the five populations of P. trituberculatus
2.3 判别分析
对5个地理种群22个比例性状的判别分析的F检验结果表明差异极显著(P<0.000 1),说明不同种群及群体间形态差异显著。为更细致的判别哪几个比例性状在三疣梭子蟹判别分析上贡献的大小,对以上实验数据进行逐步判别分析,最终的判别结果是在进行完16步逐步判别分析之后得出的,检验出CW/CL、ICW/CL、AW/CL、ML/CL、5PW/CL和OCS/CL这6个比例形状对判别分析的贡献较大。对这6个比例形状建立了5个地理种群的判别方程,公式中X1~X6分别代表CW/CL、ICW/CL、AW/CL…OCS/CL各函数的自变量和常数。
判别方程如下:

由这6个比例性状对三疣梭子蟹进行判别,对测量数据按上述公式进行预测分类,结果见表3,判别准确率P1为63.64%~90.91%,P2为67.74%~96.97%,综合判别率为82.24%。因此,上述判别方程是可靠的,可以作为判别这5个种群的参考依据。

表3 5个地理种群三疣梭子蟹逐步判别分析测试结果Tab. 3 Testing discriminant results of the five populations of P. trituberculatus
3 讨 论
3.1 三疣梭子蟹不同种群的生长与生理差异
在中国沿海野生三疣梭子蟹群体中,不同三疣梭子蟹地理种群间在生长和存活率方面存在不同程度的差异。高保全等[19]在对来自舟山种群、莱州湾种群和海州湾种群自繁一代的120日龄三疣梭子蟹生长发育和存活率研究结果见表4,相同条件下海州湾连云港群体在全甲长和体重等6个生长相关性状方面均极显著低于莱州湾野生种群和舟山野生种群(P<0.01)。

表4 三群体三疣梭子蟹的生长表型Tab. 4 Growth performance of P. trituberculatus in three populations
陈萍等[20]对海州湾、舟山沿海、辽东湾、莱州湾4 个地理群体的野生三疣梭子蟹于同一条件下孵化和饲养,各群体随机取健康蟹20 只用于试验来研究三疣梭子蟹的免疫生理差异。结果表明上述4群体中海州湾群体的超氧化物歧化酶(SOD)显著高于莱州湾群体,而对谷胱苷肽过氧化物酶(GSH-px)研究也具有同样的规律(见表5)。
鉴于中国不同海区三疣梭子蟹具有其独特的种群特性,从形态学上鉴别三疣梭子蟹不同群体对于种质鉴定、引种及资源保护均有重要价值。

表5 四种群三疣梭子蟹的免疫酶活差异Tab. 5 Activity of immune enzymes P. trituberculatus in four populations
3.2 3种多元分析在三疣梭子蟹形态判别上的应用价值
目前,等位酶、基于基因组(AFLP,RAPD,ISSR,SSR)、线粒体基因(D-Loop,16s rRNA)或特定基因(MHC)作为分子标记技术在群体遗传学、物种系统发生、种质鉴定中开展了大量的卓有成效的工作,分子生物技术已显示出了它的巨大作用。但这些研究方法还要和表型鉴定结合起来,特别是形态学性状相结合。形态学研究方法是最直接的研究手段,具有直观、方便、经济等诸多优点,仍是上述研究领域的重要研究手段。由于多元分析方法在形态研究中具有很好的应用价值,目前在鱼类[18,21]、蟹类[22-24]和贝类[25,26]研究中已取得良好的研究效果。特别是在贝类和蟹类的研究中,由于壳较硬,产生的测量误差较小,这对于形态研究是非常有利的。本实验研究使用了三种多元分析,通过测量三疣梭子蟹背甲、前额裂片、腹甲、步足和螯肢等部位的23个形态学参数,22个比例参数,可以较好地在形态上区分我国近海这5个海区的三疣梭子蟹。这三种方法分析结果相似,但各自从不同角度反映了不同种群间的形态学差异,因此都互相不可替代。聚类分析将不同种群进行初步归类,量化种群间的差异程度,分析种群间的相似程度。主成分分析将多个形态比例性状综合成少数几个因子,从而得出不同种群之间的差异大小,并可根据不同种群的主成分值找出各种群在各主成分值上差异较大的参数。判别分析是种群鉴定的常用方法,通过建立判别函数,对种或种群进行判别[27]。本研究中采用逐步判别方法对5个地理种群三疣梭子蟹的22个比例性状建立判别函数,其判别效果比较理想。逐步判别方法,可对多个因子进行合理选择,从大量因子中挑选出若干必要的最佳组合因子建立判别函数,本研究经16步判别分析后选出CW/CL、ICW/CL、AW/CL、ML/CL、5PW/CL和OCS/CL这6个比例构建的方程综合判别率仍达82.24%,这意味着这6个性状可以对5个地理种群的三疣梭子蟹种质作出初步的鉴定,具有应用价值。
3.3 5个地理种群三疣梭子蟹的形态差异
5个地理种群三疣梭子蟹的聚类分析和主成分散布图的结果一致,五个群体可以分为两支,一支由大连、东营、连云港和舟山4群体组成,而另一支则为湛江群体。聚类分析的结果中,连云港种群、东营种群、大连种群三者形态接近,形态距离最小,为0.08~0.09;舟山种群与这三个群体的形态差异略大些为0.09~0.11,而湛江种群与其他种群的趋异程度最大,均超过了0.15。这显示了湛江群体明显与其余4个群体在形态上产生了明显的差异。这种差异可能是由于地理差异和栖息环境条件不同造成的,因为大连海区,东营海区和连云港海区处于黄渤海区。大连种群来自于渤海,该海是一个深入大陆的浅海,面积较小,为北温带生物区系。与渤海相连的是黄海,为一个半封闭的海,生物区系处在北温带海的边缘,黄海环流基本上由中国沿岸地区的黄海暖流和沿岸流所组成,处于北温带,来自寒带、亚寒带、热带和亚热带的生物种群与本地土生种汇在一起构成独特的生物区系。舟山群体来自东海,大部分陆架区平均水深370 m,最深达2 719 m。东海沿岸流和台湾暖流是东海浅水区域的两支主要海流。上述4个群体在栖息地地理和气候环境均差异不是太大,均位于北回归线以北具有四季更替温差较大的特点。且黄渤海和东海相连,造成了大连、东营、连云港和舟山4群体的基因流,差异趋小。而湛江海区位于我国南海,在北回归线南边,处热带、亚热带气候,海水表层水温高 (25℃~28℃),年温差小(3℃~4℃),南海北部有沿岸流和南海暖流两大流系。终年高温高湿,长夏无冬,形成了特有的生境,因此使湛江三疣梭子蟹群体在形态上与其余4个群体有较大差异。
从生物地理学来看,本研究与冯建彬等(2005)对我国四大海区文蛤形态聚类一致,但本研究中三疣梭子蟹的形态欧氏距离(0.08~0.18)远高于对文蛤的地理种群差异(0.01~0.06)[28];从形态学来讲,这也说明三疣梭子蟹的遗传多样性要高于文蛤。从5个地理种群的主成分散布图还可以看出,湛江种群与其他4个种群的重叠区域较小,而其他4个种群之间的重叠区域较大,表明湛江种群与其他4个种群在形态上有较大的差异,而其他4个种群之间的重叠区域表示它们之间的相似程度,非重叠区域则表示它们之间的差异程度。形态上的差异是有遗传因子和环境因子共同作用的结果,这5个地理种群的差异属于种内变异,由于生活的环境不近相同,造成形态上的一定差异,同时,除了湛江种群之外的其他4个种群之间的地理距离较近,在外界因子的影响下,如水团运动、海流等,4个种群之间存在基因交流,因此,大连种群、东营种群、连云港种群以及舟山种群之间的形态差异较小。
显然,三疣梭子蟹的外部形态在中国海四大海区内既有一定的相似性,又存在较大的变异。如大连、连云港、东营和舟山相互间的相对变异较小,而与湛江与其余群体的变异则较大。这提示了中国海三疣梭子蟹存在着较大的遗传多样性,但也要看到地处黄海和渤海的大连、连云港、东营和地处东海的舟山四群体的形态相似性,这种结果究竟是海区相近造成三疣梭子蟹的自然交流还是由于人类活动造成的结果,而且这种贡献大小还不得而知。因此,应从多方面研究中国海三疣梭子蟹的种群结构和遗传多样性状况,进而进行合理的保护和有续的利用,这对于渔业管理和水产养殖都具有十分重大的意义。
[1] Aileen, T S-H, B H Y Zulfigar, Y Fujii, et al. JSPS/UCC Report:Culture of Japanese blue crab (Portunus trituberculatus) [R]. The Center for International Cooperation, The Ocean Research Institute,Univ. Tokyo, 2000, 29.
[2] Hamasaki K. Theory and practice of seed production of swimming crab [J]. Tokyo, JASFA Series, 2000(3):181.
[3] 孙玉忠, 王雪梅, 宋全山, 等. 三疣梭子蟹全人工工厂化育苗技术 [J]. 海洋湖沼通报,2002(3): 76-80.
[4] Fan Yang, Hua-Teng Xu, Zhong-Min Dai, et al. Molecularcharacterization and expression analysis of vitellogenin in the marine crab Portunus trituberculatus [J]. Comparative Biochemistry and Physiology (Part B), 2005, 142: 456-464.
[5] 薛俊增, 吴惠仙. 三疣梭子蟹卵附着机制及相关形态学特征 [J].动物学报, 2004, 50(5): 873-879.
[6] Katsuyuki HAMASAKI, Hideyuki IMAI, Nobuhiko AKIYAMA, et al. Ovarian development and induced oviposition of the overwintering swimming crab Portunus trituberculatus (Brachyura:Portunidae) reared in the laboratory [J].FISHERIES SCIENCE,2004, (70): 988–995.
[7] Ariyama, H. Studies on Ecology and Stock Enhancement of Swimming Crab Portunus trituberculatus in Osaka Bay [J]. Bulletin of the Osaka Prefectural Fisheries Experimental Station, 2000, 12:1-90.
[8] Ariyama H, T Katayama, Y Matsuda, et al. Marking and tagging methods of swimming crab Portunus (Portunus) trituberculatus. II.Method for small-sized crabs [J]. Bulletin of the Osaka Prefectural Fisheries Experimental Station, 2001, 13: 29-43.
[9] Ariyama H. How to enhance the stock of swimming crab Portunus trituberculatus by releasing juveniles [R]. Report of Osaka Prefectural Fisheries Experimental Station, 2001, 15.
[10] Kazutoshi OKAMOTO. Malformed regeneration of partly cut swimming leg as a marker for swimming crab Portunus trituberculatus [J]. FISHERIES SCIENCE, 2006, 72: 1 121–1 123.
[11] 郭天慧, 孔晓瑜., 陈四清, 等. 三疣梭子蟹线粒体DNA 16S rRNA和COI基因片段序列的比较研究[J]. 中国海洋大学学报,2004, 34(1): 22-28.
[12] Mitsugu M. Yamauchi, Masaki U. Miya, Mutsumi Nishida.Complete mitochondrial DNA sequence of the swimming crab,Portunus trituberculatus (Crustacea: Decapoda: Brachyura) [J].Gene, 2003, 311: 129–135.
[13] 高保全, 刘萍, 李健, 等. 三疣梭子蟹4个野生群体形态差异分析 [J]. 中国水产科学, 2007, 14(2): 223-228.
[14] 黎中宝, 李少菁, 王桂忠. 中国东南沿海锯缘青蟹群体的形态判别分析 [J]. 厦门大学学报, 2004, 1(43): 102-106.
[15] Clive P, Keenan, Peter J F, et al. A revision of the genus Scylla de haan, 1833 (Crustacea:Decapoda: Brachyura:Portunidae) [J]. The Raffles Bulletin of Zoology, 1998, 46(1): 217-245.
[16] Brzeski V J, Doyle R W. A morphometric criterion for sex discrimination in tilapia [C]. The Second International Symposium on tilapia in Aquaculture: ICLARM Conference Proceeding Department of Fisheries, Bangkok, Thailand, and International Center of Living Aquatic Resources Management, Manila,Philippines, 1988: 439-444.
[17] 张尧庭, 方开秦. 多元统计分析引论 [M]. 北京: 科学出版社,1982: 393-401.
[18] 李思发, 李晨虹, 李家乐. 尼罗罗非鱼品系间形态差异分析 [J].动物学报, 1998, 44(4): 450-457.
[19] 高保全, 刘萍, 李健. 三疣梭子蟹3个地理种群杂交子一代生长和存活率的比较 [J]. 大连水产学院学报, 2008, 23(5): 325-329.
[20] 陈萍, 李健, 李吉涛, 等. 不同地理群体三疣梭子蟹非特异性免疫功能的比较 [J]. 中国农学通报, 2008, 24 (11): 496-499.
[21] Ana Pinheiro, Celia M. Teixeira, Ana Luisa Rego, et al. Genetic and morphological variation of Solea lascaris (Risso, 1810) along the Portuguese coast [J]. Fisheries Research, 2005, 73: 67–78.
[22] J L. Overton D J. Macintosh R S. Thorpe. Multivariate analysis of the mud crab Scylla serrate (Brachyura:Portunidae) from four locations in southeast Asia [J]. Marine Biology, 1997, 128: 55-62.
[23] Michael S.Rosenberg. Fiddler crab claw shape variation: a geometric morphometric analysis across the genus Uca (Crustacea:Brachyura: Ocypodidae) [J]. Biological Journal of the Linnean Society, 2002, 75: 147-162.
[24] Teresa Fernandes, Jayne V Brian , Richard J Ladle, et al. Patterns of morphological and genetic variability in UK populations of the shore crab, Carcinus maenas Linnaeus, 1758 (Crustacea: Decapoda:Brachyura) [J]. Journal of Experimental Marine Biology and Ecology, 2006, 329: 47–54.
[25] J K Plejdrup, V Simonsen, C Pertoldi, et al. Genetic and morphological diversity in populations of Nucella lapillus (L;neogastropoda) in response to tributyltin contamination [J].Ecotoxicology and Environmental Safety, 2006, 64(2): 146-54.
[26] Krause M K, Arnold W S, Ambrose Jr, W. G. Morphological and genetic variation among three populations of Calico scallops,Argopecten gibbus [J]. J. Shellfish Res, 1994, 13(2): 529–537.
[27] W J Krzanowski, Francis Henry, Charles Marriott. Multivariate Analysis (part II) [M]. published Oxford University Press, 1995.
[28] 冯建彬, 李家乐, 王美珍, 等. 我国四海区不同群体文蛤形态差异与判别分析 [J]. 浙江海洋学院学报(自然科学版), 2005, 4:318-323.
Morphological variation analysis among five populations of the swimming crab Portunus trituberculatus from China Sea areas
DONG Zhi-guo1,2,3, LI Xiao-ying1, YAN Bin-lun1, Gao Huan1, WU Xu-gan2, CHENG Yong-xu2, SUN Xiao-wen3
(1. Key Laboratory of Marine Biotechnology of Jiangsu Province, Huaihai Institute of Technology, Jiangsu, Lianyungang 222005, China;2. Aquatic and Life School, Shanghai Ocean University, Shanghai 201306, China;3. Heilongjiang River Fishery Research Institute of Chinese Academy of Fishery Sciences, Haerbin 150070, China)
Based on 23 morphological characters of Portunus trituberculatus, three multivariation analysis methods(cluster analysis, principal component analysis and discriminant analysis) were used to comparably reveal the morphological variation among the five geographical populations from China Sea areas (Dalian, Dongying, Lian yungang, Zhoushan and Zhanjiang). The results of cluster analysis and principal component analysis indicated that the morphological characters were similar between Dongying and Lianyungang, but quite different from that of Zhanjiang. In the principal component analysis, three principal components were constructed, and the contributory ratio of the first principal component was 27.39%, the second was15.23%, and the third was 10.27%.The cumulative contributory ratio was 52.89%.The results of discriminant analysis indicated that there were significant differences among the five populations (P<0.01). The identification accuracy was 63.64%~94.44%(P1) and 67.74%~92.86%(P2), and the total discriminant accuracy was 82.24%.
China Sea; Portunus trituberculatus; morphological variation; multi-variation analysis
Q959.223.63
A
1001-6932(2010)04-0421-06
2009-08-11;
2009-12-25
国家科技支撑重大项目(2006BAD09A01)、江苏省海洋生物技术重点实验室开放基金项目(HS2005014)和(HS2006002)
董志国 (1977-),男,博士,讲师,主要从事水产生物种质资源与遗传育种研究。电子邮箱:dzg7712@yahoo.com.cn
孙效文 (1955-),男,研究员,主要从事鱼类育种研究。电子邮箱:sunxw2002@163.com
