东太平洋表层海水中添加氮、磷的现场培养实验
2010-09-11唐松
唐 松
(中国科学院广州地球化学研究所,广东 广州 510640)
东太平洋表层海水中添加氮、磷的现场培养实验
唐 松
(中国科学院广州地球化学研究所,广东 广州 510640)
借助“中国首次环球科学考察”航次,在东太平洋表层海水进行了添加氮、磷的现场培养实验。现场记录了实验水体温度的变化,用分光光度法对水体硝酸氮和活性磷酸盐浓度进行了检测,并用荧光法分析了水体叶绿素a浓度。结果表明,氮的添加会引起水体中叶绿素a浓度短期内快速增大,同时伴随硝酸盐浓度的显著降低,而单独添加P对水体中叶绿素a浓度影响并不显著;水体中N/P比值与叶绿素a浓度、N/P比值与浮游植物生长速度、温度与叶绿素a浓度以及温度与浮游植物生长速度之间均缺乏相关性。因此认为,在东太平洋实验海区表层海水中添加氮会引起浮游植物快速爆发,而磷的添加并不能刺激浮游植物快速生长,水体 N/P比值和水体温度都不能单独控制浮游植物群落的生长。
东太平洋;培养实验;营养盐;浮游植物
Abstract:Nutrient enrichment experiment was carried out in the East Pacific Ocean during the Chinese First Around-world Researching Cruise,with the addition of nitrogen or phosphorus to surface seawater.The variation of water temperature was recorded.The nutrient concentration was analyzed with spectrophotometry,and chlorophyll a concentration with fluorescence analysis.The addition of nitrogen resulted in the rapid increase of chlorophyll a concentration with the depletion of nitrogen in the experimental water,but no apparent variation occurred in chlorophyll a concentration when phosphate was added.The correlation was absent between the ratio of nitrogen to phosphorus and chlorophyll a concentration,the ratio of nitrogen to phosphorus and phytoplankton growing rate,water temperature and chlorophyll a concentration,and water temperature and phytoplankton growing rate.So it was considered that addition of nitrogen could stimulate phytoplankton bloom in the surface seawater in the East Pacific Ocean,while addition of phosphate could not.In addition,the ratio of nitrogen to phosphorus and the water temperature cannot control the growth of phytoplankton community.
Keywords:East Pacific Ocean; enrichment experiment; nutrient; phytoplankton
长期以来,营养盐限制作用作为影响浮游植物生长最为重要的因素之一,一直是海洋生态学研究的重要领域。目前已经认识到,在多数开阔大洋,浮游植物的生长主要受表层海水中可利用的氮和磷浓度限制[1]。但浮游植物生长到底是氮限制还是磷限制,目前还没有形成统一观点。在东热带太平洋海区的贫营养区域[2-5],贫营养的南太平洋亚热带海区[6,7],以及南海[8]的培养实验结果都表明,氮是这些海区最重要的限制营养盐。此外,美国北卡罗来纳州河口[9]、夏威夷Kaneohe海湾[10]、菲律宾西北部海域[11]等也都为氮限制。而在地中海[12]、长江口[13,14]、东海[15],以及莱州湾附近海域[16]等,则表现出磷限制。我国大亚湾海域[17,18]、台湾海域上升流区[19]以及东、黄海典型海区[20]的添加营养盐培养实验表明,氮和磷对这些海区浮游植物生长都有限制作用。
可以看出,在不同海域氮和磷对浮游植物生长的限制作用也不同。而且,目前国内关于开阔大洋氮和磷对浮游植物生长的限制作用报道较少。借助2005年“中国首次环球科学考察”航次,笔者利用东太平洋表层海水进行了添加氮、磷的现场培养实验,这为研究该海区的营养盐限制因子提供了宝贵的机会。本文通过对实验过程中水体温度、营养盐浓度、叶绿素a浓度等参数进行分析,探讨温度、营养盐氮和磷的添加,以及N/P比值等对东太平洋实验海区浮游植物生长的影响。
1 材料与方法
2005年9月,借助“中国首次环球科学考察航次”,利用东太平洋表层海水(102°37.94’W,3°15.10’S)进行了现场添加N、P的现场培养实验。实验在3个500 L的丙烯酸培养桶中进行,并配备一套冷却装置可控制水体温度,有关实验装置及其操作的详细介绍见参考文献[21]。1号桶为采样点的表层海水,2号和3号培养桶在采取的表层海水中分别添加了 0.01mol的 N(KNO3)和 0.01mol的 P(KH2PO4),实验过程持续了13天。实验海区表层海水营养盐背景值见表1。
实验过程中每间隔12 h或24 h采样一次,记录水体温度。利用7230 G分光光度计现场分析水体中的硝酸氮、活性磷酸盐浓度,同时用荧光法分析水体叶绿素a浓度。具体实验流程及营养盐和叶绿素a浓度分析方法参见参考文献[21]。

表1 东太平洋实验海水背景值Tab.1 Background value of the experimental water in the East Pacific Ocean
2 结果与讨论
2.1 营养盐的添加对浮游植物生长的影响
叶绿素a浓度是反应浮游植物生长最直接、准确的参数,同时,其变化速度反应了浮游植物的生长速度。
实验过程中水体叶绿素a浓度变化见图1。整个实验过程中1、2、3号培养桶中叶绿素a浓度平均值分别为0.68 mg/m3,1.97 mg/m3和0.83 mg/m3。由图1可以看出,各培养桶水体中叶绿素a浓度的相对快速增大发生在实验的第2-5天。其中,2号加N桶中叶绿素a浓度增大最为显著,在实验第5天达到最高值6.69 mg/m3,高出初始值(0.16 mg/m3)40多倍。1号空白桶和3号加P培养桶水体中叶绿素a浓度的最高值分别出现在实验第5天和第4天,值分别为1.51 mg/m3和1.92 mg/m3,比水体初始值高出8倍和9倍。培养实验第5天后,三个培养桶中的叶绿素a浓度均下降,至实验结束时,分别降至0.65 mg/m3,0.65 mg/m3和0.45 mg/m3。不难看出,在东太平洋表层海水中添加N对浮游植物生长有明显促进作用。虽然添加P的3号桶中叶绿素a浓度也发生较明显增大,但与1号空白桶相比,无论其变化趋势,还是平均值或者最高值,都非常接近。因此,笔者认为在该海区单独添加P对刺激浮游植物爆发并没有明显作用。

图1 实验水体中叶绿素a浓度变化Fig.1 Variation of Chl-a concentration in the experimental water
除了浮游植物生长量,其生长速率的变化也是判断浮游植物生长状态的重要参数。本文根据下面的公式计算浮游植物生长 速率(R):
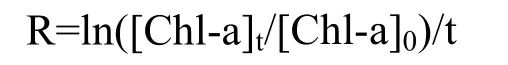
式中:t代表实验时间,[Chl-a]t和[Chl-a]0分别代表了在实验时间t时和初始时的叶绿素a浓度。
1-3号培养桶水体中平均R值分别为0.44/d、0.58/d和0.46/d。图2显示了东太平洋海水培养实验中浮游植物生长速度的变化。可以看出,三个培养桶中R值变化趋势基本一致,在实验过程前期的增大后,都表现出降低的趋势。1号和3号桶中浮游植物生长速度在实验第3天分别增至最大值0.90/d和0.87 /d。2号桶中R值的最大值出现在实验的第4天,为1.03 /d。各培养桶中R值在出现最高值后,均保持下降趋势。实验结束时,1-3号培养桶中的R值分别为0.12 /d 、0.11 /d 和0.07 /d。可见,浮游植物生长速度的变化也表明,2号桶中N的添加对实验水体中浮游植物生长影响最明显。3号桶中R值的变化趋势、平均值和绝对值均接近于1号桶,因此认为,P的添加对浮游植物的生长并没有显著影响。

图2 实验水体中浮游植物生长速度变化Fig.2 Variation of phytoplankton growth rate in the experimental water
2.2 温度对浮游植物生长的影响
东太平洋海水培养实验过程中温度变化见图3。整个实验过程平均温度为22.8 ℃,变化范围为21~26 ℃。在实验的前10天,温度曲线表现出锯齿状的变化特征,此后温度一直升高,最高值出现在实验结束时,达到26 ℃。虽然已有研究表明,水体温度对硅藻的生长有显著影响[22-24],但由于不同浮游植物种类适宜的生存温度有所差异,水体温度对整个浮游植物群落的影响要远远小于对具体种的影响[25]。在东太平洋培养实验过程中,各培养桶浮游植物相对快速生长时,水体温度并没有表现出明显的规律性变化与之对应,而且水体温度与叶绿素a浓度以及R值之间都缺乏明显相关性(表2)。因此认为,至少在该实验过程中,水体温度并不能单独主控整个浮游植物群落的生长量,关于其对具体浮游植物种类的影响还有待深入研究。

图3 实验水体中温度的变化Fig.3 Variation of temperature in experimental water

表2 温度与Chl-a浓度以及R值的相关性分析结果Tab.2 Result of correlation analysis between temperature,Chl-a and the value of R
2.3 实验水体中的营养盐浓度的变化
图4显示了实验过程中水体硝酸盐和磷酸盐浓度的变化。与各培养桶水体中叶绿素a浓度在实验第2-5天的明显增长相对应,1号和3号培养桶中水体硝酸盐浓度分别由第 2天的 6.27 μM 和3.82 μM 降低至第 4天的 0.16 μM 和 0.13 μM,2号加 N桶中的硝酸盐浓度则在实验第 2-6天快速下降,由32.65 μM降低至3.95 μM。很明显,浮游植物的生长与水体中硝酸盐的消耗存在密切联系,而且,N的添加可促进浮游植物快速爆发。同时,伴随着水体中硝酸盐浓度的降低,1号和2号培养桶中磷酸盐浓度分别由初始的1.00 μM和0.93 μM降低至实验第4天的0.07 μM和0.26 μM,3号培养桶中磷酸盐浓度初始值为18.61 μM,至实验第5天降为17.82 μM。这说明在浮游植物生长消耗水体硝酸盐的同时,也利用了其中的磷酸盐。此外,虽然 3号培养桶中的平均磷酸盐浓度高出1号和2号培养桶50多倍,但该桶中浮游植物的生长量仅接近于1号空白桶(见图),而且其中磷酸盐浓度的消耗也与其它两个培养桶中相近。这说明,磷酸盐并不是实验海区表层海水中浮游植物生长的主要限制营养盐。
此次培养实验中,三个培养桶中浮游植物生长速度变化趋势基本一致。在实验第3天前,1号和3号培养桶中R值持续增大,而2号桶中浮游植物生长速度的升高发生在实验前4天。与R值的增大对应,各培养桶水体中的硝酸盐浓度都发生降低,尤其2号加N桶中表现最明显,硝酸盐浓度在实验第5天比初始值降低了18.22 μM(见图4)。这说明N的消耗对于浮游植物生长有明显的促进作用。在R值增大期,虽然各培养桶水体中磷酸盐浓度在硝酸盐浓度降低的同时也有所减小,但在3号加P培养桶中,R值快速增大至峰值时,水体磷酸盐浓度仅减小了0.79 μM,与2号桶和3号桶中磷酸盐浓度的降幅接近。因此认为,实验海区海水中浮游植物不能单独利用P促进自身快速生长。

图4 实验水体中营养盐浓度变化Fig.4 Variation of nutrient concentration in experimental water
2.4 N/P比值与浮游植物生长的关系
图5显示了此次培养实验过程中水体N/P值的变化。考虑到1号培养桶中水体营养盐浓度偏低,实验误差对N/P比值的影响较大,在此不做讨论。2号、3号培养桶中水体平均N/P值分别为94.98和0.14。在实验第 6天之前,2号培养桶水体以高氮为特征(见图4),平均硝酸盐和磷酸盐浓度分别为21.61 μM和0.34 μM。在浮游植物爆发时,N/P比值由初始值 32.29突增至实验第 4天的 385.60。3号培养桶水体始终以高磷为特征(见图4),硝酸盐和磷酸盐的平均浓度分别为0.69 μM和17.42 μM。在浮游植物相对快速生长期内,水体N/P比值由实验第2天的0.44下降至第4天的0.07。可见,在2号和3号培养桶水体中浮游植物相对快速生长时,N/P比值的变化规律并不统一。已有研究表明,水体中N/P比值与浮游植物生长特性有很大关系[26],而且不同种类浮游植物生长的最佳营养盐条件对N/P比值的要求也不同[27-29]。由于添加营养盐不同,2号和3号培养桶中浮游植物组成肯定不同。正是由于满足这些浮游植物有利生长的N/P比值要求不同,造成在2号和3号培养桶中叶绿素a浓度相对快速增大时,水体N/P比值变化表现出不同特征。另外,对实验过程中2号和3号培养桶水体N/P比值和Chl-a浓度以及R值进行相关性分析的结果(表3)表明,水体中的 N/P比值同浮游植物生长量和生长速度之间无明显相关性。这说明东太平洋实验海区水体中的N/P比值不能单独控制浮游植物群落的整体生长特性。

图5 实验水体中N/P值变化Fig.5 Variation of the ratio of N to P in experimental water

表3 N/P比值与Chl-a浓度以及R值的相关性分析结果Table.3 Result of correlation analysis between the ratio of N to P ,Chl-a and the value of R
此外,在高氮的2号培养桶中,无论是整个实验过程的平均N/P值,还是浮游植物快速生长时的N/P平均值都远大于16。而在高磷的3号桶中,水体N/P比值则一直远小于16。根据Redfield定律,2号培养桶中浮游植物生长应受P限制,3号桶中则为N限制。而前面的分析已表明,在2号桶中,浮游植物生长与水体硝酸盐浓度的降低有明显的对应关系;而在3号桶中,水体中硝酸盐浓度在实验第3天就降低至接近表层海水的水平,浮游植物生长也没有表现出受N限制的影响。可见,Redfield定律有其一定的适用范围,水体N/P比值并不能单独决定浮游植物群落生长的营养盐限制因子。
3 结 论
利用东太平洋表层海水进行了添加氮、磷的现场培养实验,通过对水体温度、营养盐浓度和叶绿素a浓度等参数分析认为:
a)至少在该实验过程中,水体温度不能单独控制浮游植物群落生长;
b)实验海区水体中浮游植物生长表现出明显的N限制作用,单独添加P并不能刺激浮游植物短期内快速爆发;
c)水体中的N/P比值既不能控制游植物群落的整体生长特性,也不能决定其限制因子。
致谢:感谢彭晓彤老师、林荣澄老师、蒋磊和吴自军博士,以及“大洋一号”船员在现场实验过程中给予的大力支持和帮助!
[1]Gruber N.The dynamics of the marine nitrogen cycle and its influence on atmospheric CO2 variations [A].The ocean carbon and climate [C].Dordrecht: Kluwer Academic,2005: 97.
[2]Thomas W H.Surface nitrogenous and phytoplankton in the northeastern tropical Pacific Ocean [J].Limnol Oceanogr,1966,11:393-400.
[3]Thomas W H.The nitrogen nutrition of phytoplankton in the northeastern tropical Pacific Ocean [J].Stud Trop Oceanogr Miami,1967,5: 280-289.
[4]Thomas W H.Phytoplankton nutrient enrichment esperiments off Baja California and in the eastern equatorial Pacific Ocean [J].J Fish Res Bd Can,1969,26: 1133-1145.
[5]Thomas E.Effect of ammonium and nitrate concentration on chlorophyll increases in natural tropical pacific phytoplankton populations [J].Limnol Oceanogr,1970,15: 386-394.
[6]Dufour P,Charpy L,Bonnet S,et al.Phytoplankton nutrient control in the oligotrophic South Pacific subtropical gyre [J].Mar Ecol Prog Ser,1999,179: 285-290.
[7]Dufour P,Berland B.Nutrient control of phytoplankton biomass in atollagoons and Pacific Ocean waters: Studies with factorial enrichment bioassays [J].J Exp Mar Bio Ecol,1999,234: 147-166.
[8]Chen Y L,Chen H C,Karl D M,et al.Nitrogen modulates phytoplankton growth in spring in the South China Sea [J].Continental Shelf Research,2004,24: 527-541.
[9]Mallin M.Phytoplankton ecology of North Carolina estuaries [J].Estuaries,1994,17: 561-574.
[10]Larned S T.Nitrogen-versus phosphorus-limited growth and sources of nutrients for coral reef macroalgae [J].Marine Biology,1998,132 (3): 409-421.
[11]Terrados J.nutrient limitation of the tropical sea grass Enhalus acoroides (L.)Royle in Cape Bolinao,NW Philippines [J].Aquatic Botany,1999,65 (14): 123-139.
[12]Thingstad T,Frede U L Z,Fereidoun R P.1imitation of heterotrophic bacteria and phytoplankton in the northwest Mediterranean [J].Limnology and Oceanogrophy,1998,43 (1):88-94.
[13]Ning X,Vaulot D,Liu Z,et a1.Standing stock and production of phytoplankton in the estuary of the Changjiang (Yangtze River)and the adjacent East China Sea [J].Marine Ecology Progress Series,1988,49 (10): l4l-150.
[14]胡明辉,徐春林.长江口浮游植物生长的磷酸盐限制 [J].海洋学报,1989,11 (4): 439-443.
[15]Wong G T F,Gong G C,Liu K K,et a1.Excess Nitrate’in the East China Sea [J].Estuarine,Coastal and Shelf Science,1998,46:411-4l8.
[16]邹立,张经.渤海春季营养盐限制的现场实验 [J].海洋与湖沼,2001,32(6): 672-678.
[17]王晓伟,李纯厚,李占东,等.大亚湾海域春季浮游植物生长的限制性营养元素 [J].中国水产科学,2007,14(5): 836-842.
[18]朱艾嘉,黄良民,许战洲.氮、磷对大亚湾大鹏澳海区浮游植物群落的影响 [J].热带海洋学报,2008,27(1): 38-45.
[19]王玉珏,洪华生,王大志,等.台湾海峡上升流区浮游植物对营养盐添加的响应 [J].生态学报,2008,28(3): 1 321-1 327.
[20]刘媛,曹振锐,黄邦钦,等.东、黄海典型海区浮游植物对营养盐添加的响应 [J].厦门大学学报(自然科学版),2004,43(增刊):147-152.
[21]唐松.中大西洋营养盐富集实验中浮游植物生长的营养盐限制作用 [J].矿物岩石地球化学通报.2008,27(2): 112-118.
[22]Eppley R W.Temperature and phytoplankton growth in the sea [J].Fishery Bulletin,1972,70: 1 063-1 085.
[23]Goldman J C,Carpenter E J.A kinetic approach to the effect of temperature on algal growth [J].Limnology and Oceanography,1974,19: 756-766.
[24]Yoder J A.Effect of temperature on light-limited growth and chemical composition of Skeletonema costatum (Bacillariophyceae)[J].Journal of Phycology,1979,15: 362-370.
[25]Sarmiento j L,Gruber N.Ocean biogeochemical dynamics [M].Princeton University Press,2006.
[26]John E H,Flynn K J.Growth dynamics and toxicity of Alexandrium fundyense (Dinophyceae): The effect of changing N:P supply ratos on internal toxin and nutrient levels [J].Eru J Phycol,2000,35:11-23.
[27]候继灵,张传松,石晓勇,等.磷酸盐对两种东海典型赤潮藻影响的围隔实验 [J].中国海洋大学学报,2006,36: 163-169.
[28]康燕玉,梁君容,高亚辉,等.氮、磷比对两种赤潮藻生长特性的影响及藻间竞争作用 [J].海洋学报,2003,28(5): 117-122.
[29]刘东艳,孙军,陈宗涛,等.不同氮磷比对中肋骨条藻生长特性的影响 [J].海洋湖沼通报,2002,2: 39-44.
In situ nutrient enrichment experiment with addition of nitrogen or phosphorus to surface seawater in the East Pacific Ocean
Tang Song
(Guangzhou Institute of Geochemistry,Chinese Academy of Sciences,Guangzhou 510640,China)
Q178.1+12; Q179.1
A
1001-6932(2010)04-0402-05
2009-11-09;
2009-11-25
国家自然科学基金资助项目(40903033,40403004)
唐松(1979-),男,陕西渭南人,博士,副研究员,主要从事海洋地球化学研究。电子邮箱:tangsong@gig.ac.cn
