RNA干扰PAR基因表达对PC3细胞生长的影响
2010-08-14徐晓峰周秀敏张征宇葛京平周文泉程文位志峰侯健全高建平
徐晓峰 周秀敏 张征宇 葛京平 周文泉程文 位志峰 侯健全 高建平
1. 南京军区南京总医院泌尿外科,江苏 南京 210002;2. 苏州大学附属第一医院泌尿外科,△肿瘤科,江苏 苏州 215006
当前在美国前列腺癌高居男性肿瘤发病的首位[1],目前我国前列腺癌亦呈不断上升的趋势。内分泌治疗作为晚期前列腺癌主要的治疗方法,但最终会因为前列腺癌转变为雄激素非依赖性导致治疗失败[2-3]。前列腺癌由雄激素依赖性转变为雄激素非依赖性的机制是目前前列腺癌研究的热点。PAR(prostate androgen regulated)是新近发现的一种雄激素非依赖性前列腺癌特异性高表达的癌基因。已有研究提示PAR基因可能是与细胞的恶性转化具有相关性[4],但对于PAR基因高表达是否和雄激素非依赖性前列腺癌细胞的恶性表型相关,目前尚未见报道。本研究中采用RNA干扰技术抑制PAR基因在雄激素非依赖性前列腺癌细胞系PC3中的表达,观察PC3细胞恶性表型的变化并初步研究其机制,从而更为深入地了解PAR基因的功能。
1 材料和方法
1.1 细胞和试剂 人前列腺癌细胞株PC3购自中国典型培养物保藏中心。质粒psiRNA-hH1-neo购自InvivoGen公司。限制性内切酶BbSⅠ购自Fermentas公司。总RNA提取试剂TRIzol和脂质体LipofectamineTM2000购自Invitrogen公司。M-MLV反转录酶购自Promega公司。高纯度质粒提取试剂盒购自上海华舜生物工程有限公司。
1.2 方法
1.2.1 ShRNA表达质粒的设计和构建 从GenBank经比对获得人PAR基因(序列号为AF115850)序列,根据shRNA的设计原则,从起始密码子下游100 nt后选取3个符合设计特征的靶序列,分别命名为PAR-a、PAR-b和PAR-c(表1)。用BLAST软件进行分析,未发现所选取的核苷酸序列与人体其他任何mRNA有同源性。
依据以上所选的序列设计3对53nt的寡核苷酸,相应分别命名为PAR1、PAR2和PAR3。每条寡核苷酸两端为带有BbSⅠ酶切位点的序列,两个20 nt反向互补排列的PAR特异性序列,中间由一个5 nt的间区隔开,转录后可形成茎环结构(表2)。寡核苷酸由上海博亚生物技术有限公司合成。

表1 ShRNA靶向的PAR基因特异性位点Tab.1 ShRNA targeting regions of PAR gene
将合成的单链DNA经退火处理获得双链结构,插入已经用BbSⅠ酶切处理过的线性质粒psiRNA-hH1-neo中,构建获得重组体psiRNAPAR1、psiRNA-PAR2和psiRNA-PAR3,转化感受态细菌DH5α,挑取生长良好的克隆送上海博亚生物技术有限公司测序鉴定。
1.2.2 细胞培养和质粒转染 人前列腺癌细胞株PC3在含10%小牛血清的高糖型DMEM培养液中,置于37 ℃,CO2体积分数为5%的条件下培养传代。将PC3细胞以3×105个/孔的细胞密度接种于6孔板中。当细胞生长密度达到50%~70%时,用脂质体法转染,具体操作按试剂说明书步骤进行。实验组中每孔加入500 μL含有6 μL LipofectamineTM2000和2 μg质粒的OPTI-MEM无血清培养液,同时设立对照质粒组。
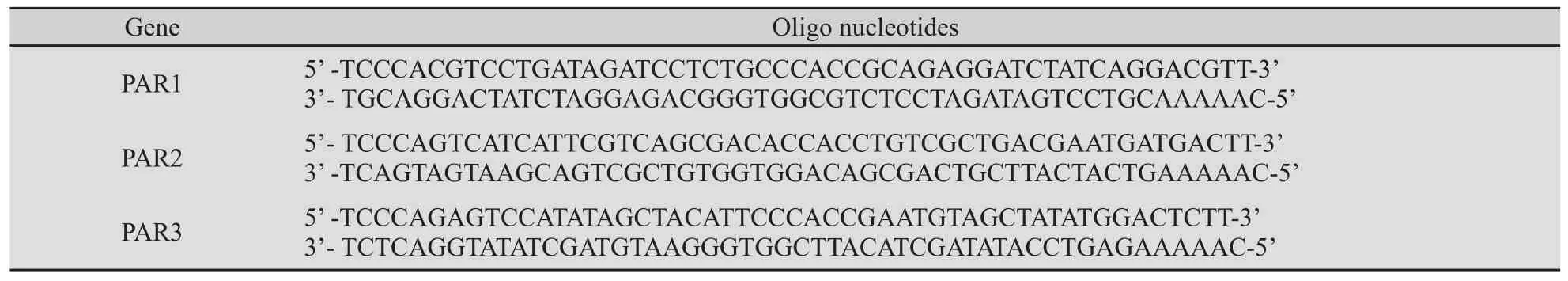
表2 合成的53nt寡核苷酸Tab.2 Oligo nucleotides
1.2.3 RT-PCR法检测PAR基因的表达采用 RT-PCR 筛选有效抑制PAR基因表达的shRNA表达质粒和转染时间:转染48 h后,收集各组细胞,提取总RNA,以RNA反转录后合成的第一条cDNA链为模板,进行PCR扩增。PAR基因的引物序列,上游引物5’-GTCAGCAAGCACCTCAAAT-3’,下游引物5’-GAAGAAGATGGGGAAAAGG-3’;内参照β-actin基因的引物,上游引物5’-GTGCCACCAGACAGCACTGTGTTG-3’;下游引物5’-TGGAGAAGAGCTATGAGCTGCCTG-3’。 PAR基因和β-actin的PCR扩增产物片段的长度分别为451 bp和202 bp。反应条件:94 ℃ 5 min;94 ℃30 s, 55 ℃ 30 s,72 ℃ 1 min(共30个循环);最后72 ℃ 延伸10 min。扩增后的产物行1%琼脂糖凝胶电泳鉴定,溴化乙锭染色,再经凝胶成像系统成像,用Gelworks 1D Advanced v4.01软件进行条带灰度分析。
1.2.4 测定细胞生长曲线 取对数生长期细胞,0.25%的胰酶消化后,转染前24 h以5×104个/mL的细胞密度接种于24孔板中。转染后,每天取3个孔的细胞计数,取其平均值,共7 d,绘制成生长曲线。
1.2.5 软琼脂克隆形成实验 将约50 ℃、含0.6%琼脂、10%小牛血清的DMEM培养液3 mL灌入直径为60 mm的培养皿中,置于室温待其凝固。将2 mL的50 ℃、含200个细胞、0.3%琼脂和10%小牛血清的DMEM培养液再均匀灌入入已铺有底层培养基的60 mm培养皿中,置于37 ℃,CO2体积分数为5%条件下培养15 d,然后在光学显微镜下计数含50个以上细胞的克隆,计算克隆形成率。每实验组设3个平行样本,实验重复2次,计算平均值。
1.2.6 流式细胞仪分析细胞调亡和细胞周期 用筛选获得的抑制效应最明显的shRNA表达质粒psiRNA-PAR1转染PC3细胞,转染后24和48 h后收集细胞,然后用PBS洗涤细胞2次,再以500 μL PBS重悬细胞,加入-20 ℃预冷的85%乙醇溶液,4 ℃固定过夜。再用PBS洗涤细胞2次,加入100 μL PC 缓冲液(phosphate-citric acid buffer)(0.2 mol/L Na2HPO4和0.1 mol/L citric acid按192∶8的体积比混匀,pH值为7.8)室温条件下避光处理20 min。再以PBS洗涤1次,加入碘化丙啶(propidium iodide,PI)染液500 μL(含10 μg/mL的PI和100 μg/mL的RNase A),避光,室温条件下处理30 min后上机,用流式细胞仪进行细胞调亡和细胞周期的检测。
1.3 统计处理 所有数据采用统计学软件SPSS 11.0行单因素方差分析。以P<0.05为差异有统计学意义。
2 结 果
2.1 shRNA表达质粒的构建 成功构建获得shRNA表达质粒,经测序证实,针对PAR基因的3个特异性靶序列的寡核苷酸均成功插入psiRNA-hH1-neo质粒,和设计序列完全一致。
2.2 转染shRNA表达质粒对PC3细胞PAR mRNA表达的影响 RT-PCR检测结果显示, PC3细胞中psiRNA-PAR1、psiRNAPAR2和psiRNA-PAR3对PAR mRNA的表达均有一定的抑制作用,其中以psiRNA-PAR1抑制效果最好,这3种表达质粒和对照质粒组相比平均抑制率分别为(81.18±1.68)%、(47.65±1.97)%和(5.25±0.59)%,差异有显著的统计学意义(P<0.01,图1)。
在转染48 h后,psiRNA-PAR1对PAR mRNA表达的抑制率达最高峰,为(81.18±1.68)%;而24和72 h时的抑制率分别为(28.96±3.95)%和(68.67±1.00)%,经比对,差异有显著的统计学意义(P<0.01,图2)。
2.3 转染shRNA表达质粒对PC3细胞生长速率的影响 根据细胞计数结果绘制细胞生长曲线,由生长曲线结果提示:转染psiRNA-PAR1的细胞生长速率减慢最明显,相同时间内细胞增长数最少;转染psiRNA-PAR2的细胞次之;转染psiRNA-PAR3的细胞生长减慢变化最小,这与RT-PCR检测结果一致(图3)。
2.4 转染shRNA表达质粒对PC3细胞克隆形成能力的影响 软琼脂克隆检测结果显示:转染了质粒psiRNA-PAR1、psiRNA-PAR2和psiRNA-PAR3的PC3细胞的克隆形成率分别为0、2%和48%,与转染空载体组的62%和空白对照组(未转染质粒)的68%相比较,明显降低(P<0.05),尤其是psiRNA-PAR1组的细胞克隆形成能力几乎丧失。
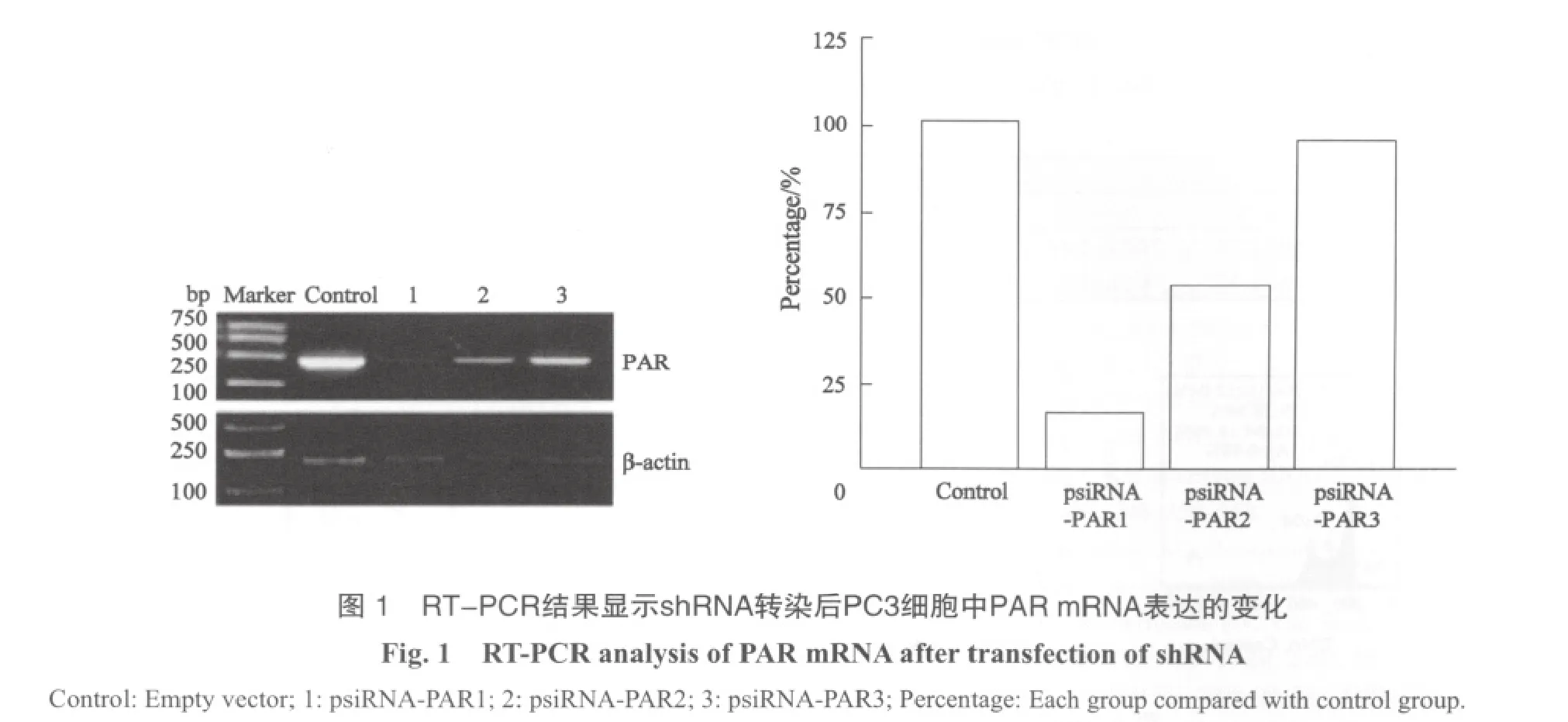
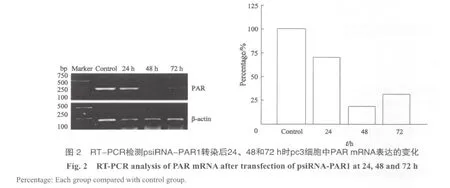
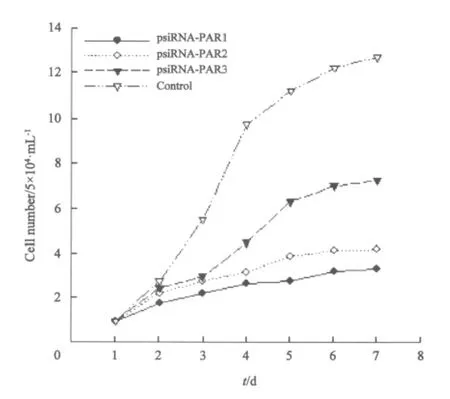
图3 转染shRNA后PC3细胞的生长情况Fig.3 Growth of PC3 cells after transfected with shRNA
2.5 细胞调亡和细胞周期变化 FCM法检测结果提示,shRNA 转染PC3细胞24和48h时,对照组处于G2/M期的细胞分别为(21.39±1.39)%和(23.79±3.16)%,凋亡率分别为(0.30±0.04)%和(1.49±0.13)%;而 psiRNA-PAR1组处于G2/M期的细胞分别为(39.32±1.56)%和(29.95±3.25)%,凋亡率分别为(12.16±1.18)%和(20.61±2.73)%,与对照组相比,差异有显著的统计学意义(P<0.01,表3,图4),说明转染psiRNAPAR1对PC3细胞有明显的细胞周期阻滞作用,使细胞阻滞在G2/M期,并诱导PC3细胞发生凋亡。
表3 psiRNA-PAR1转染对PC3细胞周期和凋亡的影响Tab.3 The effects of siRNA on cell cycle and apoptosis of PC3 cells( )

表3 psiRNA-PAR1转染对PC3细胞周期和凋亡的影响Tab.3 The effects of siRNA on cell cycle and apoptosis of PC3 cells( )
?

图4 psiRNA-PAR1转染对PC3细胞周期和细胞凋亡的影响Fig.4 The effects of siRNA on cell cycle and apoptosis of PC3 cells
3 讨 论
临床研究结果显示,在晚期前列腺癌内分泌治疗过程中癌细胞出现雄激素非依赖性是治疗失败的主要原因[8]。近年来,人们在分子水平对雄激素非依赖性前列腺癌的发病机制和治疗进行了大量的研究,不断寻找新的致癌基因并作为其潜在的分子靶点[9-10],而其中具有前列腺癌组织特异性的致癌基因更有研究价值[4-5]。
新近发现的在前列腺癌中表达异常的PAR基因位于人1号染色体上,片段长度为1 038 bp,编码146个氨基酸,其在正常的前列腺组织和癌组织中均有表达,但大约67%标本中,癌组织中有更高的表达;PAR在LNCaP、DU145、PC3和LNCaP-OM等前列腺癌细胞系中的表达也比正常的前列腺上皮细胞要高;而在雄激素非依赖性细胞系DU145、PC3和LNCaPOM中的表达明显高于雄激素依赖性细胞系LNCaP。雄激素可以使雄激素依赖性细胞系的PAR表达下调,而对雄激素非依赖性细胞系的PAR表达则无抑制作用[6]。Platica等[7]将PAR cDNA重组质粒转染成纤维细胞NIH3T3,使其获得恶性表型,但抑制PAR基因在雄激素非依赖性前列腺癌细胞系中的表达是否会使其恶性表型逆转,目前尚无相关研究报道。
RNA干扰(RNA interference, RNAi)技术的作用机制是:以同源互补序列的mRNA为靶目标,通过活化的21~23 nt的小片段干扰RNA(siRNA),降解特定的mRNA,使目的基因表达下调。siRNA作用的高效性、特异性和 稳定性均优于反义核酸和核酶等反义技术[11]。本实验采用了psiRNA-hH1-neo质粒系统,成功构建了针对PAR基因3个不同位点的siRNA表达质粒并转染PC3细胞,通过抑制PAR基因的表达研究其功能。转染shRNA表达质粒的PC3细胞系与对照组相比,PAR基因的mRNA表达明显下调,提示RNAi技术可以有效抑制PAR的表达,有望作为前列腺癌基因治疗的有效手段。本研究利用细胞计数、软琼脂克隆形成实验表明PAR表达下调可明显抑制PC3细胞恶性增殖。为探讨细胞生长抑制的机制,进一步利用流式细胞术检测,发现PAR基因的mRNA表达下调能够明显引起PC3细胞阻滞于G2/M期,凋亡增加。G2/M期检查点是细胞存活和死亡的重要决定点,细胞经过此点即进入分裂期[12-13]。下调PAR基因的表达可使肿瘤细胞生长变慢,其主要机制可能是诱导肿瘤细胞G2/M期阻滞和凋亡。这一结果提示PAR基因可能参与前列腺癌的恶性转化,PAR基因可能是一个新的与雄激素非依赖性前列腺癌细胞恶性增殖密切相关的癌基因,有望作为今后前列腺癌基因治疗的有效靶点之一。
[1] Greenlee RT, Hill-Harmon MB, Murray T, et al. Cancer statistics, 2001[J]. CA Cancer J Clin, 2001, 51(1): 15-36.
[2] Hegeman RB, Liu G, Wilding G, et al. Newer therapies in advanced prostate cancer[J]. Clin Prostate Cancer, 2004,3(3): 150-156.
[3] Miyamoto H, Messing EM, Chang C. Androgen deprivation therapy for prostate cancer: current status and future prospects[J]. Prostate, 2004, 61(4): 332-353.
[4] Shi XB, Gumerlock PH, de Vere White RW. Molecular biology of prostate cancer[J]. World J Urol, 1996, 14(5): 318-328.
[5] Huncharek M, Muscat J. Genetic characteristics of prostate cancer[J]. Cancer Epidemiol Biomarkers Prev, 1995, 4(6):581-687.
[6] Platica O, Chen S, Ivan E, et al. PAR, a novel androgen regulated gene, ubiquitously expressed in normal and malignant cells[J]. Int J Oncol, 2000, 16(5): 1055-1061.
[7] Platica M, Ivan E, Ionescu A, et al. Transformation of NIH3T3 cells by enhanced PAR expression[J]. Biochem Biophys Res Commun, 2004, 314(3): 891-896.
[8] Bonaccorsi L, Muratori M, Carloni V, et al. Androgen receptor and prostate cancer invasion[J]. Int J Androl, 2003; 26(1):21-25.
[9] Horvath LG, Henshall SM, Kench JG, et al. Loss of BMP2,Smad8 and Smad4 expression in prostate cancer progression[J]. Prostate, 2004, 59(3): 234-242.
[10] 李燕,周志毅,石群立. 前列腺癌的分子病理学研究[J]. 医学研究生学报, 2007, 20(10): 1088-1093.
[11] Brummelkamp TR, Bernards R, Agami R. A system for stable expression of short interfering RNAs in mammalian cells[J].Science, 2002, 296: 550-553.
[12] Labazi M, Phillips AC. Oncogenes as regulators of apoptosis[J]. Essays Biochem, 2003, 39: 89-104.
[13] Vermeulen K, Berneman ZN, Van Bockstaele DR. Cell cycle and apoptosis[J]. Cell Prolif, 2003, 36(3): 165-175.
