天然人源IgG Fab噬菌体抗体库的构建及多样性分析①
2010-07-30邵红霞付永峰HiroshiTachibana程训佳
邵红霞 章 黎 杨 彬 冯 萌 付永峰 Hiroshi Tachibana 程训佳
(复旦大学上海医学院病原生物学系,上海200032)
健康人外周血B淋巴细胞具有受抗原物质刺 激活化,分化成效应细胞即浆细胞从而分泌相应抗体的潜能。抗体多样性在免疫球蛋白的发生机制上包括了多种淋巴细胞特异的分子机制,特别是不同基因家族的重链V(variable)-D(Diversity)-J(joining)片段的重排与轻链V-J片段的重排及轻重链的不同组合。抗体基因重排过程中随机发生的体细胞突变又进一步增加了抗体的多样性。抗体多样性的分子发生机制为B淋巴细胞产生无限多样性的免疫球蛋白基因产物提供了理论解释。最新数据库资料显示重链可变区基因(VH)可分成7个基因家族,VH1~VH7,其中每个基因家族之间的同源性可达80%以上[1]。一般认为,在健康个体,VH基因的表达常取决于各基因家族的基因片段的数量,其中VH3基因家族包括了21个功能基因片段,因而最常使用(约45.6%),而VH5、VH6及VH7家族仅有1个功能基因,因而表达频率极低[2]。然而近年来,越来越多的研究表明,免疫球蛋白基因的表达不是随机的。无论是感染某种特定病原体的患者还是健康个体,其外周血B淋巴细胞VH基因的表达均可能有偏向选择,表明存在特定的分子机制而非仅VH基因的阳性选择[3]。
此外,轻链基因通过与重链基因组合的多样性共同影响免疫球蛋白的抗原结合部位,轻链基因分κ和λ两种,一般认为在人体内的比例约2∶1,κ轻链的功能基因分属于1~5个家族,以Vκ1基因家族最多见;λ轻链的功能基因包括了11个基因家族,以Vλ3家族最多见。
基因工程抗体库技术是近年来发展起来的体外制备人单克隆抗体的新方法,利用抗体库技术可以筛选到针对任何抗原分子的抗体并可进行人工改造以提高亲和力等各种生物学效应。分析已经建成的抗体库基因特别是VH基因可以在一定程度上反映B淋巴细胞抗体基因的多样性及可能存在的偏向选择,为合理使用抗体库制备相应抗体及进行后续的抗体优化提供借鉴和参考并为外周血淋巴细胞来源的抗体基因的优势选择提供实验室依据。故本课题收集了300名健康志愿者的外周血淋巴细胞,分别获得人免疫球蛋白的重链Fd和轻链基因,构建天然人源IgGFab噬菌体抗体库。
1 材料与方法
1.1 材料
1.1.1 淋巴细胞、载体和菌株 近2个月未患感冒等感染性疾病的健康志愿者抗凝外周血300份(每份2 ml),供血者为中国人,年龄20~40岁之间,以25~30岁年龄段居多,男女比例约1∶1。
噬菌体载体pFabICN(4.4 kb,其中酶切位点NheⅠ至AscⅠ为轻链插入位置,酶切位点SfiⅠ至NotⅠ为重链插入位置)由东海大学实验室提供[4]。测序载体CV-1和CV-2为本实验室自建。电转和普通大肠埃希菌(Escherichia coli)JM109购自大连宝生物工程(TaKaRa)公司。
1.1.2 主要实验材料 葡聚糖-泛影酸钠(瑞典Pharmacia公司);总RNA提纯试剂盒(德国Qiagen公司);Gene Amp RNA PCR Kit,限制性内切酶AscⅠ、NheⅠ、SfiⅠ、NotⅠ,100 bp Ladder DNA,1 kb Ladder DNA,Lambda HindⅢDNAMarker(美国New England公司);TaKaRa Ex Taq DNA聚合酶、限制性内切酶λ Hind Ⅲ、DNA Ligation Kit Ver2.0(TaKaRa公司)等。
1.2 方法
1.2.1 人外周血淋巴细胞分离及总RNA提取 分批采集300份健康志愿者抗凝外周血各2 ml,分别以淋巴细胞分离液分离获得外周血淋巴细胞后混合,以灭菌PBS洗涤后重悬细胞,共获得约8×108个细胞,总RNA提纯试剂盒提取外周血淋巴细胞总RNA。
1.2.2 人免疫球蛋白Fd重链及轻链基因的扩增 以Gene Amp RNA PCRKit进行RT-PCR:先将淋巴细胞总RNA逆转录成cDNA,然后以一组几乎覆盖人免疫球蛋白G(IgG)全部轻链或重链可变区编码序列的特异性引物[5],对上述反应产物行PCR以分别扩增免疫球蛋白κ、λ轻链和γ重链Fd片段。扩增条件为:94℃2分钟,然后94℃0.5分钟,55℃1分钟,72℃1分钟,共30个循环,最终延长为72℃7分钟,其中72℃1分钟步骤每个循环加1秒。PCR产物电泳鉴定后回收纯化。
1.2.3 人源IgG Fab噬菌体抗体库的构建 轻链基因κ、λ分别以限制性内切酶 AscⅠ、NheⅠ酶切、纯化后以κ∶λ轻链基因7∶1混合后与噬菌体载体pFabICN连接,连接产物提纯后溶出于4μl蒸馏水中,分2次电转JM109感受态细胞后将转化菌均匀涂布于含50μl氨苄青霉素的LB平板,收集所有克隆的质粒DNA,即为轻链抗体库。
γ重链Fd片段基因PCR产物以SfiⅠ、NotⅠ酶切,纯化后与轻链库DNA连接,同法电转化获得所有克隆的质粒DNA,即为抗体库DNA,同时计数菌落数目以计算抗体库效价。
1.2.4 人源IgGFab噬菌体抗体库的鉴定及多样性分析 抗体库DNA分三次转化JM109感受态细胞,随机挑选克隆,提取质粒DNA,首先以λHindⅢ酶切初步鉴定,进一步以AscⅠ、NheⅠ或SfiⅠ、NotⅠ双酶切后鉴定轻重链插入片段酶切位点是否正确。对轻重链均插入正确的质粒,以双酶切分别获得轻链、重链基因片段,分别与测序载体CV-2、CV-1连接,送Invitrogen(英潍捷基贸易有限公司)以ABI 3730 DNA测序仪测定核苷酸序列并推算氨基酸序列,利用IgBLAST数据库分析抗体基因的同源家族信息。
2 结果
2.1 人免疫球蛋白Fd重链及轻链基因的扩增 来源于300份健康志愿者的外周血淋巴细胞的总RNA进行RT-PCR,将获得的免疫球蛋白轻链和γ重链Fd片段经2.0%琼脂糖凝胶电泳鉴定,显示重链Fd基因长度约为690 bp,轻链κ.λ基因长度约为660 bp(图1)。
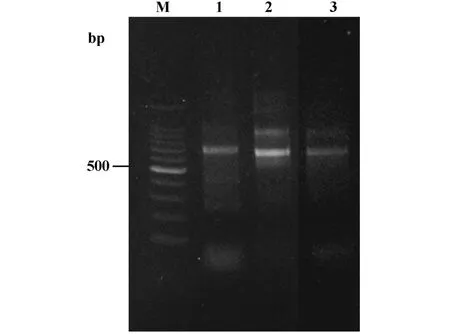
图1 重链和轻链基因PCR扩增产物Fig.1 PCR products of heavy and light chain genes
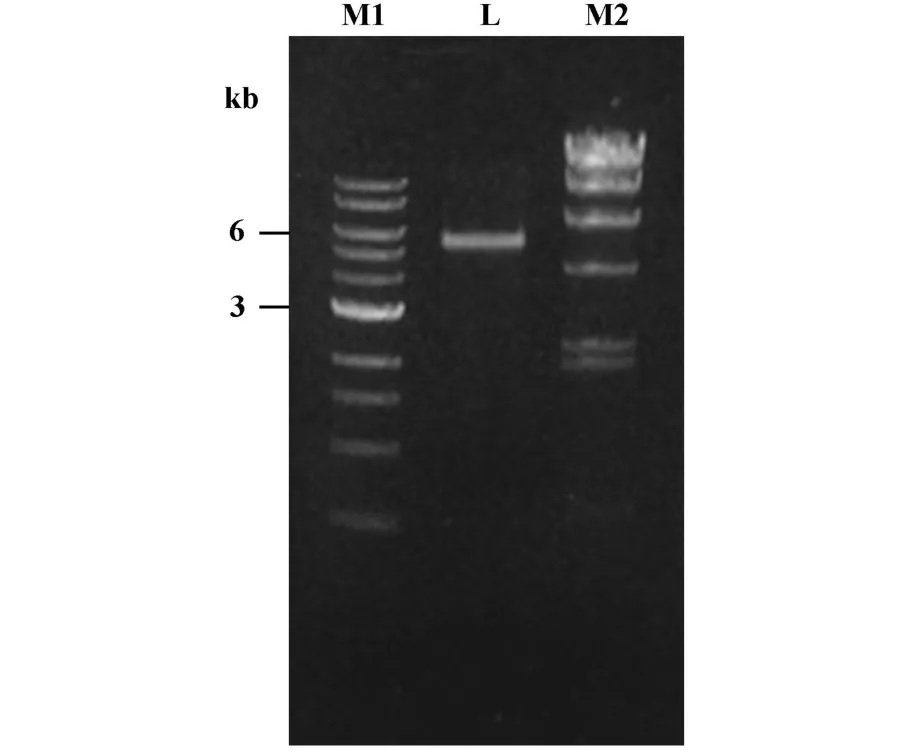
图2 人源IgG Fab噬菌体抗体库Fig.2 Human phage-displayed Fab library
2.2 人源IgGFab噬菌体抗体库的构建 将所有淋巴细胞来源的轻链基因(κ、λ)与噬菌体载体pFabICN连接建成轻链库,进一步将重链Fd基因片段连接于轻链库DNA相应位置中,最终获得非依赖性克隆滴度(即库容量)为4×108的人源IgG Fab噬菌体抗体库。抗体库DNA的0.8%琼脂糖凝胶电泳结果见图2。
2.3 抗体库的鉴定和抗体基因分析 对三次随机挑选克隆获得的轻重链基因插入正确的32个克隆进行轻重链基因测序,以IgBLAST数据库对测序结果进行核苷酸序列分析和氨基酸推算。在总共32条重链核苷酸序列中,其中2条重链基因较正常明显缩短,另1条不属于人免疫球蛋白基因,其余29条重链Fd基因片段彼此完全不同并能正确翻译,占所测序列的90.6%。所有29条重链所对应的轻链基因序列大小正常并能正常翻译。
经IgBLAST比对,29条重链基因的V、D、J片段所属的基因家族分布见图3所示,其中重链可变区V片段中,VH3基因家族有21条,占72%,VH1基因家族3条(10.3%),VH4、VH5基因家族各 2条(各6.9%),VH7基因家族1条(3.4%)。属于VH3基因家族的21条重链基因包括了3-7、3-30(各4条),3-23、3-48、3-66(各 2 条),3-9 、3-11、3-15、3-20、3-21、3-43和3-74(各1条);重链可变区D片段分别属于1~6基因家族,以DH3家族最多(58.6%);重链可变区J片段分别属于1~6家族,以JH4家族最多(31.0%)。29条轻链基因可变区V片段分别属于Vκ1(13 条,44.8%)、Vκ2(4 条,15.4%)、Vκ3(3条,11.5%)、Vκ4(6 条 ,23.1%)基因家族和 Vλ1(3 条,11.5%)基因家族;轻链基因可变区J片段分别属于Jκ1~ 2,Jκ4~ Jκ5和 Jλ3(见图 4)。

图3 天然人源噬菌体抗体库中重链基因家族分布图Fig.3 Distribution of heavy chain genefamilies in a naïve human phage-displayed Fab library
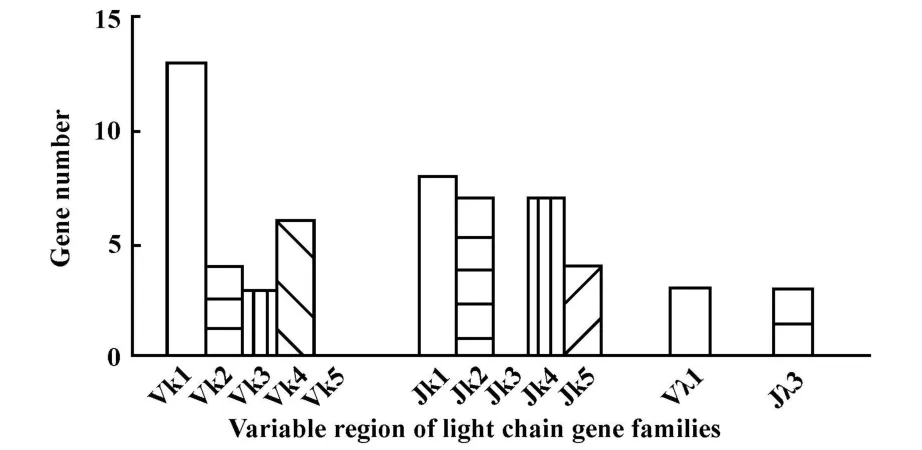
图4 天然人源噬菌体抗体库中轻链基因家族分布图Fig.4 Distribution of light chain gene families in a naïve human phage-displayed Fab library
3 讨论
抗体库的库容量及多样性是衡量抗体库质量的重要技术指标,理论上,抗体库容量越大,多样性越好,筛选到针对任意抗原表位抗体的概率也就越大,亲和力也可能越高。虽然由于体内B淋巴细胞种类和数量有限,同时受克隆清除和克隆无能导致免疫耐受的影响,天然抗体库的容量受限,但天然抗体库是由经体内重排的抗体基因构成,其产生的克隆均为有效克隆,即100%具有活性[6]。因此,对于天然抗体库而言,广泛的基因来源不仅有助于消除不同个体之间的差异更重要的是可以增加基因来源的多样性,同时在建库过程中将轻重链基因分别插入载体的相应位置,有助于增加轻重链基因组合的多样性,对提高抗体库质量意义重大。本研究以来源于300名健康志愿者的外周血淋巴细胞作为抗体库基因的来源,有利于增加抗体库基因来源的多样性,为构建高质量的抗体库奠定了基础。增加转化次数是增大库容的途径之一,Vaughan等[7]通过上百次的电转化获得了1.4×1010的抗体库。然而通过这种方式获得的大库容抗体库又不可避免地影响了抗体库的多样性。本研究在建库过程中努力平衡库容量和库多样性两大技术指标,从控制实验过程中最佳反应条件等细节因素着手,而不刻意以增加转化次数等手段扩库,构建了库容4×108的原始抗体库。建库的抗体基因来源于有活性的B淋巴细胞(相比静息的B淋巴细胞,含有更多的mRNA),又有助于提高抗体库基因产物中有功能抗体的比例[8]。
本研究对人源IgG Fab噬菌体抗体库中随机获得的29条Fd重链基因序列的分析显示,重链VH3基因家族的表达频率最高,VH2和VH6表达频率最低,与Tiller等[9]的研究非常近似,该研究从 3个健康人外周血中获得183个IgG+记忆B细胞的单克隆中提取cDNA进行分析,VH基因家族的分布频率为:VH1(15.8%),VH2(0%),VH3(54.8%),VH4(23.5%),VH5(6%),VH6(0%),VH7(0.7%)。王晋等[10]构建了来源于20位健康人外周血淋巴细胞的2×108库容的噬菌体抗体库并从该抗体库中随机挑选克隆46个进行测序,发现重链VH基因家族的分布趋势为VH1>VH4>VH3>VH6>VH2>VH5,其中VH3仅20%,与本研究结果差异较大。有关健康成人外周血B淋巴细胞VH基因的表达趋势,多个实验室曾做过相关研究,Guigou等[11]采用原位杂交方式进行的研究表明VH基因的随机分布方式与各基因家族中功能基因的数目相关,即VH3>VH4>VH1>VH5>VH6>VH2,但这种结果未被广泛接受,Huang等利用二步PCR法分析B淋巴细胞库发现不同个体存在不同的VH基因家族的偏向性[12],Davidkova等[13]也有类似发现,该研究运用原位杂交的方法分析8个健康成人的外周血淋巴细胞,结果发现各健康个体之间VH基因家族的使用频率存在很大差异,如VH3家族多数集中在40.4%~83.3%之间,平均达57.9%,是所有基因家族中使用频率最高的一族,唯有一人表达仅8.5%,但其表达VH6高达76.6%,而VH6是表达频率最低的一族,VH分布频率总体概括为:VH3>VH5>VH2>VH1>VH4>VH6。值得一提的是,外周血淋巴细胞VH基因的表达受到内在基因和外在环境等多方面因素的影响,不同的体外分析手段、免疫球蛋白基因来源及个体之间均存在差异,导致在现有条件下很难对B淋巴细胞VH基因分布频率进行完全正确和全面的评估。现有多数研究结果表明,健康人免疫球蛋白基因存在VH3基因家族的优势表达,而VH6几乎是各家族中表达最低的一族,而对于VH1、VH2、VH4及VH5的表达情况,各研究结果之间差异较大,以VH4为例,有研究认为仅次于VH3[9],也有数据显示其表达频率更低[13],本研究结果介于二者之间,即VH3>VH1>VH4、VH5>VH7>VH2、VH6。VH4基因家族优势表达于部分自身免疫性疾病患者,健康个体可能存在VH4的阴性选择以避免发生自身免疫性疾病[14],这是否可用来解释健康人存在不同的阴性选择从而导致VH4基因分布频率上的不一致性尚不明确。
本研究获得的重链可变区VH片段基因分布频率的结果与直接来源于外周血B淋巴细胞的研究结果具有较好的符合性,反映出在建库过程中使用的一组几乎覆盖人IgG全部轻链或重联可变区编码序列的特异性引物确实具有较好的覆盖性。此外,由于重链可变区D片段基因和J片段基因(分别属于1~6个基因家族)分布范围较广,有利于V-D-J片段重排产生多样性良好的重链基因。轻链可变区的V片段基因分别属于Vκ1~4和Vλ1基因家族,以Vκ1基因家族最多见;J片段基因分别属于Jκ1~2、Jκ4~5和Jλ3基因家族,有利于轻链基因V-J片段以多种排列组合方式重排。轻重链基因的不同组合进一步增加了抗体的多样性。利用抗体库技术还可以对抗体重轻链可变区特别是6个互补决定区(CDR)进行定向突变,以获得具有更好特异性和更强亲和力的抗体,进一步增加了抗体库的多样性,从而完善和补充现有的抗体库。
1 Pallarès N,Lefebvre S,Contet V et al.The human immunoglobulin heavy variablegenes[J].Exp Clin Immunogenet,1999;16:36-60.
2 Gorny M K,Wang X H,Williams C et al.Preferential use of the VH5-51 gene segment by thehuman immune response tocode for antibodies against the V3 domain of HIV-1[J].Mol Immunol,2009;46:917-926.
3 Brezinschek H P,Brezinschek RI,Lipsky P E.Analysisof the heavy chain repertoire of human peripheral B cells using single-cell polymerase chain reaction[J].JImmunol,1995;155:190-202.
4 Takekoshi M,Maeda F,Tachibana H et al.Humanmonoclonal anti-HCMV neutralizing antibody from phage display libraries[J].J Virol Methods,1998;74:89-98.
5 Tachibana H,Cheng X J,Watanabe K et al.Preparation of recombinant human monoclonal antibody Fab fragments specific for Entamoebahistolytica[J].Clin Diagn Lab Immun,1999;6(3):383-387.
6 Sheet M D,Amersdorfer P,Finnem R et al.Efficient construction of a largenonimmune phage antibody library:the production of high-affinity human single-chain antibodies to protein antigens[J].Proc Natl Acad Sci USA,1998;95:6157-6162.
7 Vaughan TJ,Williams SC,Pritchard K et al.Human antibodies with subnanomolar affinities isolated from a large non-immunized phage display library[J].Nat Biotechnol,1996;14:309-314.
8 Longtenberg T,Schhutte M EM,Ebeling S B et al.Molecular approaches to the study ofhuman B cell and autoantibody repertoiregeneration and selection[J].Immunol Rev,1992;128:23-47.
9 Tiller T,Tsuiji M,Yurasov S et al.Autoreactivity in human IgG+memory B cells[J].Immunity,2007;26:205-213.
10 王 晋,罗文新,李利峰 et al.多样性人源天然噬菌体抗体库的构建及初步应用[J].细胞与分子免疫学杂志,2006;22(6):786-793.
11 Guigou V,Cuisinier A M,Tonnelle C et al.Human immunoglobulin VH and V repertoire revealed by in situ hybridization[J].Mol Immunol,1990;27:935-940.
12 Huang C,Stewart A K,Schwartz RS et al.Immunologlobulin heavy chain geneexpression in peripheral blood B lymphocytes[J].J Clin Invest,1992;89:1331-1343.
13 Davidkova G,Pettersson S,Holmberg D et al.Selective usage of VH genes in adult human B lymphocyte repertoires[J].Scand J Immunol,1997;45:62-73.
14 Pugh-Bernard A E,Sliverman G J,Cappione A J et al.Regulation of inherently autoreactive VH4-34 B cellsin themaintenanceof human B cell tolerance[J].J Clin Invest,2001;108:1061-1070.