低温弱光对苗期紫花苜蓿根颈生理特性的影响
2010-07-25张彩峡呼天明
张彩峡,呼天明
(1.西北农林科技大学动物科技学院,陕西杨凌 712100;2.西藏职业技术学院,西藏拉萨 850030)
①根颈是指根部上端膨大处,是联系地上部和根系的关键部位,根颈上能够形成大量的芽,是产生分枝的重要部位[1],同时也是吸收、运输、储存养分和水分的重要器官,直接影响植物的再生性、耐寒性、抗旱性和抗病虫害性等生产性能和植物的可持续利用[2-3]。苜蓿Medicago sativa根颈为冷冻害的最敏感部位[4],对苜蓿越冬和春季返青时的萌芽发枝至关重要。目前,国内外有关苜蓿根系的研究主要集中于根系形态发育、根蘖特性与环境的关系等方面[5],有关苜蓿根颈的研究较少[6],只有少量研究涉及根颈病害及防治[5]。试验采用人工控制温度及光照条件的方法,分析了低温弱光胁迫对不同秋眠级紫花苜蓿根颈的影响,以期为紫花苜蓿的品种选育及栽培提供支持,为苜蓿种质资源的鉴定、利用提供理论参考。
1 试验材料与方法
1.1 试验材料试验选用的2个紫花苜蓿品种(WL414和巨人201)的种子由杨凌金道种子公司提供,秋眠级分别为6和2,发芽率90%以上。
1.2 处理方法将供试紫花苜蓿种子先用98%浓硫酸处理 1 h,再用5%次氯酸钠溶液消毒5 min,最后用去离子水清洗5遍。将100粒种子置于铺2层滤纸的培养皿(直径10 cm)中,于培养箱(ZPQ350型)内20℃恒温连续暗培养3 d。试验期间根据种子吸胀情况适当补加蒸馏水。
试验用苗于2009年6月1日-11月10日在西北农林科技大学全自动PVC温室(光照培养16 h,暗培养8 h,恒温25℃)营养盆培养。营养盆为圆锥形,内径10 cm,高 25 cm,盆底有小孔,营养盆内装细沙(0.8 kg/盆),浇透水后将发芽的幼苗从培养皿移栽于营养盆中(1株/盆),每隔1 d浇1次自来水,每周浇2次Hoagland营养液以促进其生长。植株到分枝期(形成大量分枝)时,将完整植株移至人工气候培养箱(ZPQ350型)内,进行不同低温弱光处理,处理方式见表1。
处理72 h后,采取根颈样品,测定各生理指标。每处理中各参试材料分别为5株,每项测定设3次重复。
1.3 测定项目与方法参照高俊凤主编的《植物生理学实验指导》的方法[7]测定超氧化物歧化酶(SOD)、过氧化物酶(POD)活性和丙二醛(MDA)含量。

表1 处理方式
1.4 统计分析运用SPSS13.0数据进行统计分析,Microsoft Office Excel 2003进行数据运算及图表处理。
2 结果与分析
2.1 低温弱光对SOD活性的影响SOD作为植物体内保护酶之一,能清除过多积累的超氧阴离子自由基,避免或减轻逆境伤害。低温弱光处理72 h后,WL414和巨人201根颈中的SOD活性显著降低(图1)。72 h后,WL414根颈的SOD活性各处理分别比 CK下降 57.24%、41.06%、54.12%和 38.63%,巨人201根颈中的SOD活性各处理分别比 CK下降 29.65%、15.01%、27.47%和13.52%。相同弱光条件下,低温0~5℃处理SOD活性下降幅度显著大于5~10℃处理(P<0.05)。相同低温条件下,弱光50 μ mol/(m2·s)处理SOD活性下降幅度显著大于 100 μ mol/(m2·s)处理(P<0.05),即2个品种的T1和T3处理的SOD活性下降幅度均大于T2和T4,T1的下降幅度最大,T4的下降幅度最小,且 WL414的SOD活性下降程度大于巨人201。在相关研究中也得到一致结论,秋眠级高的紫花苜蓿在低温胁迫下SOD活性下降程度大于秋眠级低的[8]。
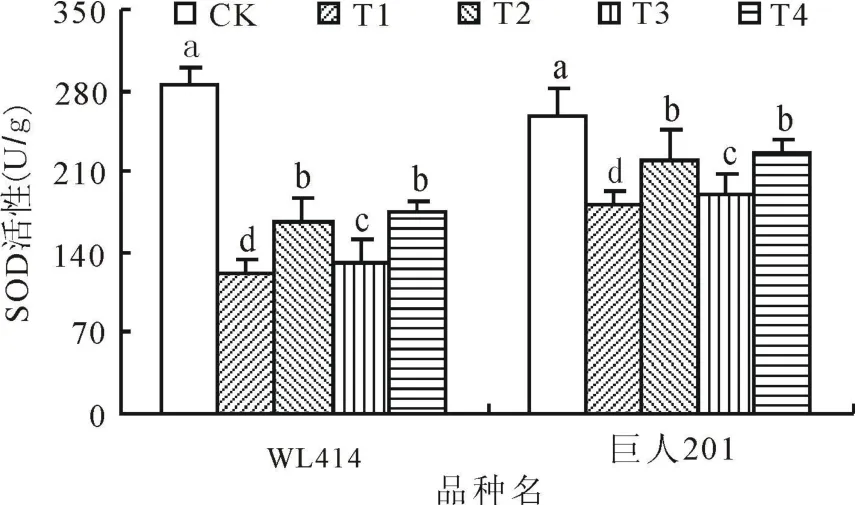
图1 低温弱光胁迫对紫花苜蓿根颈SOD活性的影响
2.2 低温弱光对POD活性的影响POD是植物体内普遍存在的、活性较高的一种保护酶,是分解H2O2的主要保护酶,其活性的高低及增强幅度的大小直接关系到植物细胞的损伤程度。低温弱光处理72 h后,WL414和巨人201根颈中POD活性呈显著上升趋势(图2)。相同弱光条件下,低温5~10℃处理POD活性增加幅度显著大于0~5℃处理(P<0.05)。相同低温条件下,弱光50和100 μ mol/(m2·s)处理组间 WL414差异不显著,而巨人201的POD活性增加幅度弱光50 μ mol/(m2·s)处理显著大 于 100 μ mol/(m2·s)处理(P<0.05)。WL414根颈的POD活性增加幅度小于巨人201,WL414根颈的POD活性各处理分别比CK增加13.30%、20.20%、10.20%和 18.70%,而巨人201根颈中POD活性各处理则分别比CK增加41.97%、50.43%、35.83%和44.27%,T2和 T4处理的POD活性增加幅度均大于T1和T3,其中T2的增加幅度最大,T3的增加幅度最小。从整个变化过程来看,巨人201保持着较高的POD活性,具有较高的H2O2清除能力,抵御低温弱光伤害的能力较强。

图2 低温弱光胁迫对紫花苜蓿根颈POD活性的影响
2.3低温弱光对MDA含量的影响MDA是膜脂过氧化产物,是反映细胞膜伤害程度的重要指标之一。低温弱光处理72 h后,WL414和巨人201根颈中MDA含量显著上升(图3)。相同弱光条件下,低温0~5℃处理MDA含量增加幅度显著大于5~10℃处理(P<0.05)。相同低温条件下,弱光 50 μ mol/(m2·s)处理MDA 含量增加幅度显著大于 100 μ mol/(m2·s)处理(P<0.05),WL414的MDA含量增加幅度大于巨人201。WL414根颈的MDA含量各处理分别比 CK 增加 79.53%、33.57%、62.90%和29.97%,而巨人201根颈中的MDA含量各处理则分别比CK增加21.44%、5.57%、14.99%和1.92%。T1和T3的MDA含量增加幅度均大于T2和T4,其中 T1的增加幅度最大,T4的增加幅度最小。与CK相比,低温弱光胁迫均导致紫花苜蓿根颈中MDA含量的上升,这说明经低温弱光胁迫,紫花苜蓿根颈细胞膜遭到不同程度的破坏[9],MDA大量积累。
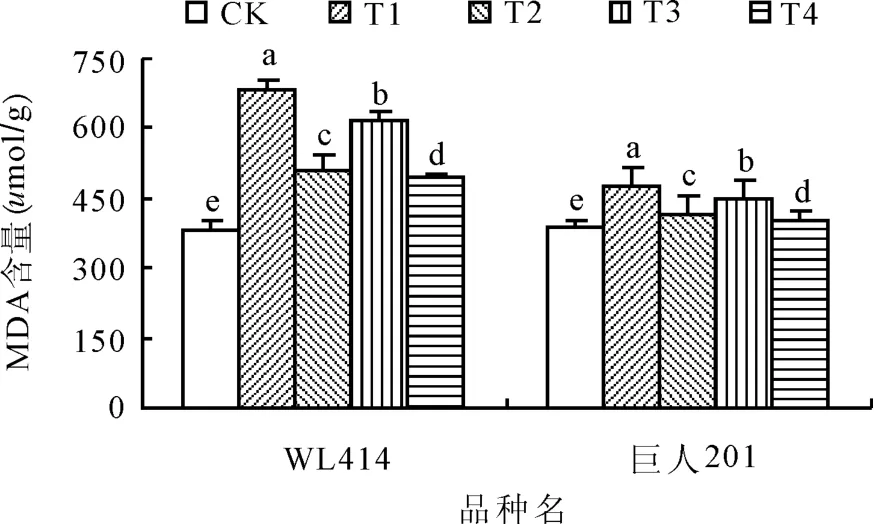
图3 低温弱光胁迫对紫花苜蓿根颈MDA含量的影响
3 讨论与结论
植物体内清除活性氧的酶类主要包括SOD、过氧化氢酶(CAT)和POD等,这些酶能减轻或消除逆境对植物造成的伤害,清除自由基并防止自由基毒害,它们活性的变化可作为植物的耐寒指标[10]。当植物遭遇逆境胁迫时,往往保护酶活性越高其忍耐逆境的能力越强。近年来的研究表明,逆境下植物体内活性氧超过正常水平时,加剧了膜脂过氧化而导致膜系统受损,最终使组织结构遭到破坏。SOD的主要功能是催化超氧物阴离子自由基(O2-)发生歧化反应,生成 O2和H2O2。H2O2可以产生更多的羟基自由基,影响植物生长与光合生理特性,而POD可以清除体内的H2O2,维持体内的活性氧代谢平衡,保护膜结构,从而使植物能在一定程度上忍耐、减缓或抵抗逆境的胁迫。MDA作为膜系统磷脂不饱和脂肪酸的过氧化产物对细胞膜起伤害作用[10]。
本试验中,低温弱光处理72 h后,WL414和巨人201根颈中的POD活性上升,但SOD活性下降,MDA含量增大,表明为避免遭受低温弱光逆境的伤害根颈作出了适应性反应,而根颈活性氧积累过多,超出了SOD和POD等膜保护酶的清除能力,部分未能清除的活性氧引起膜脂过氧化(MDA含量的增加显著),导致膜的完整性被破坏。就相同弱光条件而言,低温0~5℃对植株的影响比5~10℃更为显著,POD活性上升幅度较小,而SOD活性的下降、MDA含量的增加幅度较大,这与蔡仕珍等[11]在花叶细辛Asarum gephilum的MDA变化研究中的结果一致,随着湿度的降低MDA含量升高,使膜系统伤害更为严重。相同低温条件下,弱光 50 μ mol/(m2·s)处理使POD活性上升、SOD活性的下降、MDA含量的增加幅度比 100 μ mol/(m2·s)更大。从品种间比较可知,巨人201对低温弱光胁迫的适应能力强,能维持较高水平的SOD和POD活性[12-13],避免或减轻膜脂过氧化作用,细胞组织受的伤害较小[14]。结果表明,SOD和 POD活性、MDA含量可较好地反映植物抗逆能力[15]。
植物耐低温弱光的机制相当复杂,仅从某一侧面或层次去研究植物的耐低温弱光能力是远远不够的。植物耐低温弱光的表现是多方面的,紫花苜蓿根颈的耐低温弱光性与成株阶段及开花结实阶段的耐低温弱光性是否一致,仍有待于进一步研究。
[1]耿华珠,吴永敷,曹致中,等.中国苜蓿[M].北京:中国农业出版社,1995.
[2]Marquez-Ortiz J J,Johnson L D,Basigalup D H,et al.Crown morphology relationships among alfalfa plant introduction and cultivars[J].Crop Science,1996,36:766-770.
[3]王月胜,于林清,张利军.播种当年苜蓿根系研究初报[J].草地学报,2008,16(3):313-315.
[4]张宝田,穆春生,李志坚,等.紫花苜蓿受冻害后促进根颈枝条再生方法的研究[J].中国草地,2003,25(5):48-52.
[5]孟嫣,李敏权.苜蓿根和根颈腐烂病病原及防治研究进展[J].草业科学,2005,22(5):89-92.
[6]刘玉华,贾志宽.苜蓿秋眠性的研究进展[J].陕西农业科学,2002(7):20-22.
[7]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[8]史莹华,王成章,张伟毅,等.苜蓿秋眠型与其超氧化物歧化酶和过氧化物酶关系的研究[J].草业科学,2009,26(8):128-131.
[9]施征.三个枣品种抗寒生理特性的研究[D].河北:河北农业大学,2003.
[10]崔国文.紫花苜蓿田间越冬期抗寒生理研究[J].草地学报,2009(2):145-150.
[11]蔡仕珍,潘远智,陈其兵,等.低温胁迫对花叶细辛生理生化及生长的影响[J].草业学报,2010,19(1):95-102.
[12]寇建村,杨文权,贾志宽,等.不同苜蓿品种抗寒性研究[J].中国草地学报,2008(4):11-15.
[13]魏臻武,王德贤,贺连昌.超氧化物歧化酶在苜蓿抗寒锻炼过程中的作用[J].草业科学,2006,23(7):15-18.
[14]卢欣石,申玉龙.苜蓿秋眠性的研究与应用[J].国外畜牧业——草原与牧草,1992(1):1-4.
[15]何云.苜蓿秋眠性研究进展[J].草业科学,2005,22(11):25-29.
