低温胁迫对冬小麦恢复生长后植株细胞膜透性和丙二醛含量的影响
2010-07-09陈禹兴付连双王晓楠李卓夫
陈禹兴,付连双,王晓楠,李卓夫
(东北农业大学农学院,哈尔滨 150030)
全球气候的变暖为高纬度地区冬小麦的种植提供了条件,上个世纪90年代国家开展冬麦北移项目以来,东北农业大学开展冬麦的育种工作,东农024品系2007年经黑龙江品种审定委员会审定命名为东农冬麦1号,该品种能在黑龙江省冬季积雪较多的北部和东部种植,在冬季积雪较少的西部种植风险很大,因此要求育种上必须培育出抗寒性更强,适应范围更广的冬小麦新品种[1-3]。目前,冬小麦安全越冬受两个因素限制,一是越冬期的低温伤害;二是返青期倒春寒的伤害。
从返青开始到起身之前,历时约1个月,属苗期阶段的最后一个时期,称返青期,倒春寒是指3月下旬至4月气温持续回升,突遭寒流袭击,气温急剧下降到0℃以下的气候现象。在黑龙江这种高寒地区,倒春寒更是频繁出现。此时黑龙江省的冬小麦正处于返青起身的关键时期,抗寒性相对较弱,比较容易受冻。寒流过后气温急剧上升,冬麦幼苗在很短时间内经历了气温“回升-急剧下降-急剧升高”的过程,原本受冻的细胞由于温度回升过快其受损程度进一步加重[3-4]。本试验拟通过自然低温驯化后的不同负积温冷冻处理,以及在冷冻处理恢复生长后给予不同的低温胁迫,对不同抗寒性的冬麦品种进行细胞膜透性和膜脂过氧化终产物丙二醛(MDA)含量比较研究,探讨恢复生长后冬麦植株体内细胞膜透性和丙二醛含量的变化规律与抗寒性之间的关系,为寒地冬小麦返青期抗寒性的生理基础研究奠定基础。
1 材料与方法
1.1 材料
以抗寒性存在显著差异的东农冬麦1号(返青率为78%,DN1)、024-4(返青率为79%)、03-528(返青率为 54%)及济麦 22(返青率为 0%,JM22)四个品种为材料。供试材料于2009年9月10日播种。
1.2 方法
1.2.1 试验处理
采用塑料方盒种植冬小麦品种,方盒规格为550 mm×400 mm×200 mm。在露天自然条件下进行低温驯化。盆中土壤为细筛过的黑壤土,土层深度15 cm,土壤水分为最大持水量的70%。每盆土壤施入氮、磷、钾纯量分别为3、3.6和1.5 g。采用的肥料为尿素、二铵和硫酸钾。每盆种植150株,每品种每处理种植3盆。
根据不同冬麦区冬季负积温适应性,共设置120、240和480℃3个冷冻负积温处理。当负积温累积到这3个温度点时,分别取回4份供试材料,并置于当日相同温度下平衡72 h。随后由各平衡温度开始进行升温处理。升温处理分为两个阶段,自平衡温度以2℃·h-1,升温至-7℃后,再以1.5℃·h-1升温至5℃。5℃平衡48 h后,置于12~14℃室温下使其恢复生长10 d后取样。
恢复生长后的低温程序:模拟返青期倒春寒的温度变化对恢复生长10 d后的供试材料进行低温胁迫处理。降温以2℃·h-1的速度进行,详细温度处理如下:

1.2.2 膜透性的测定
采用电导法测定[5]。取分蘖节0.2 g,切薄片,置于20 mL烧杯中。加入10 mL去离子水后,用塑料膜封住瓶口,用针头在塑料膜上均匀刺20个小孔,将这些小烧杯放入真空器中抽真空15 min。抽真空后将烧杯在室温下放入振荡培养箱中振荡1 h,测定其电导率(S1),然后将烧杯放入沸水浴中15 min,杀死分蘖节组织,取出冷却至室温,并在室温下在振荡培养箱中振荡10 min,测定其电导率(S2)。相对电导率的计算公式:S1/S2。
1.2.3 丙二醛(MDA)含量的测定
参照赵黎芳等方法[6]。取1.5 mL粗提液A,加入2.5 mL 0.5%硫代巴比妥酸(用5%三氯乙酸溶解定容)的溶液。混合物沸水浴30 min,冷却后再离心一次分别测定上清液在450、532和600 nm处的吸光度值,并按公式算出MDA浓度,再算出单位鲜重组织中的MDA含量(μmol·g-1)。
计 算 公 式 :C(μmol·L-1)=6.45(A532-A600)-0.56A450
式中,A532、A600、A450分别代表450、532和600 nm波长下的吸光度值。
2 结果与分析
2.1 相对电导率的变化
逆境会导致细胞损伤,膜透性增大,因此可以测定相对电导率来反映逆境对植物组织伤害的程度[7]。越冬期间东农冬麦1号、03-528品系和024-4品系3个品种(系)的相对电导率随着温度的降低而逐渐升高。济麦22相对电导率随着温度的下降表现为先下降后大幅升高的趋势。东农冬麦1号、03-528和024-4品系3个品种(系)在负积温120℃和240℃下却没有明显差异,负积温240℃到480℃处理下,济麦22上升幅度显著高于其他3个品种(系),这说明与其他3个品种(系)相比,济麦22的相对电导率在低温胁迫时受温度影响较大。在480℃负积极温处理中济麦22的相对电导率达到0.6,与其他品种相比差异极显著,受低温伤害严重。在负积温为480℃时,济麦的相对电导率高于0.5,表明此时济麦22已经死亡[7]。结果见图1)。比较各品种在恢复生长后经受低温胁迫时,不同品种的相对电导率变化趋势,结果见图2。


在不同冷冻负积温处理中,相对电导率均随着处理温度的降低而升高。120℃负积温处理中,其中济麦22在-6℃和-9℃低温胁迫时相对电导率显著上升,达到0.69和0.71,较-3℃胁迫下相对电导率增幅为70%;东农冬麦1号、03-528和024-4品系3个品种(系)虽有上升但是上升幅度较缓。240℃的负积温处理中,东农冬麦1号、03-528和024-4品系恢复生长后,在-3℃和-6℃两个处理温度下品种(系)相对电导率差异不显著,而经受-9℃低温胁迫时显著上升,其中以024-4上升幅度最大。480℃负积温处理中,恢复生长后植株相对电导率随着低温胁迫的加剧而上升,03-528品系显著高于东农冬麦1号和024-4品系。这说明随着冷冻负积温的积累,各品种恢复生长后对低温胁迫的敏感度增加。03-528品系对低温的敏感度比东农冬麦1号和024-4品系两个品种(系)高。
比较在不同负积温冷冻处理后恢复生长的植株在经受相同低温胁迫处理时相对电导率的变化,结果见图3~6。未经受低温胁迫时,不同负积温冷冻处理后恢复生长的植株相对电导率差异不明显;-3℃低温处理下,负积温240℃冷冻处理恢复生长后,各品种的相对电导率略低于另两个处理。在-6℃低温胁迫下,负积温120℃相对电导率高于另两个处理,处理内,济麦22相对电导率显著高于其他品种(系)。在-9℃低温胁迫下,负积温120℃冷冻处理恢复生长后,济麦22高于其他品种,负积温240℃时,冬麦1号、024-4、03-528的相对电导率差异不显著,负积温480℃时,只有03-528的电导率有所升高,冬麦1号和024-4升高不显著。由此可以看出,适当的负积温积累可以在一定范围内提高细胞膜对渗透胁迫的抗性。
2.2 丙二醛(MDA)含量的变化
MDA是细胞膜脂过氧化反应的重要产物,通常以MDA含量作为膜脂过氧化反应的主要指标,MDA含量的多少体现了细胞膜被破坏的程度[8]。由图7可以看出,负积温为120℃时济麦22品种的MDA含量显著高于东农冬麦1号、03-528品系和024-4品系3个品种(系)。

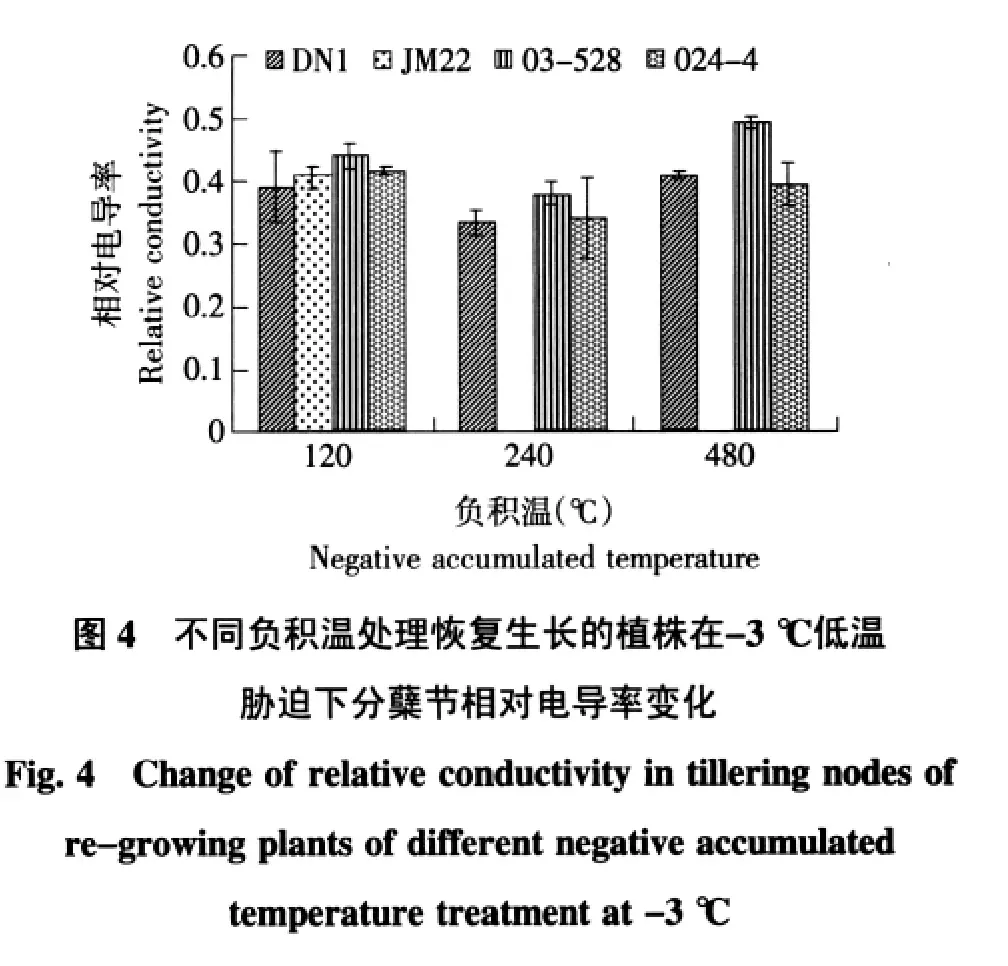
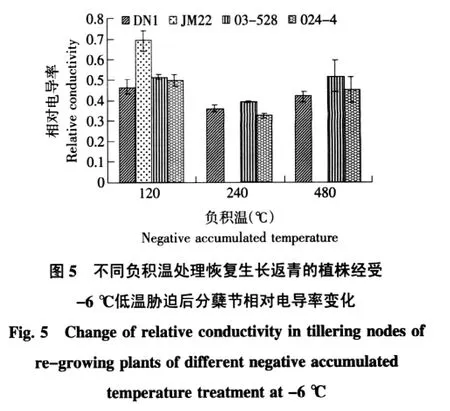


图7 负积温处理下不同抗寒性冬小麦分蘖节中丙二醛含量变化Fig.7 Change of MDA content in tillering nodes of different cold resistance winter wheat in negative accumulated temperature treatment
随着负积温的增加,东农冬麦1号、03-528品系的MDA含量升高,而济麦22、024-4品系的含量却下降。负积温240℃时,济麦22的MDA含量仍然高于其他品种(系),024-4品系的MDA含量显著低于其他品种。负积温480℃下,东农冬麦1号、济麦22、03-528品系的MDA含量较负积温240℃时略有下降,024-4品系的MDA含量上升,4个品种(系)之间差异不显著。
比较分析各品种在冷冻处理恢复生长后经受低温胁迫时,不同品种的MDA含量变化趋势,结果见图8。负积温120℃冷冻处理的植株恢复生长,济麦22丙二醛含量高于东农冬麦1号、03-528品系和024-4品系,在未经受低温胁迫和经受-9℃胁迫时这种差异最显著。各品种未经受低温胁迫时MDA含量最低,-3℃和-6℃胁迫下的各品种MDA含量差异不显著,济麦22和03-528品系在-6℃胁迫下的MDA含量相比-3℃胁迫分别下降 0.4 和 0.56 μmol·g-1。-9 ℃胁迫下各品种MDA含量显著上升,其中济麦22上升到7.12 μmol·g-1增幅最大,为73.34%。负积温240℃冷冻处理的植株恢复生长后,03-528品系的MDA含量在不同温度低温胁迫中均高于东农冬麦1号、024-4品系。-6℃胁迫处理下各品种的MDA含量较-3℃和-9℃胁迫处理的含量有所降低。负积温480℃冷冻处理的植株恢复生长后,东农冬麦1号、03-528和024-4品系3个品种(系)之间的MDA含量差异比前两个处理品种间差异小,且在-6℃低温胁迫下各品种的MDA含量最低。通过以上分析可以得出,抗寒性弱的品种在低温下MDA含量较高,而不同返青处理的植株-6℃低温胁迫下的MDA含量较其他低温处理时低。

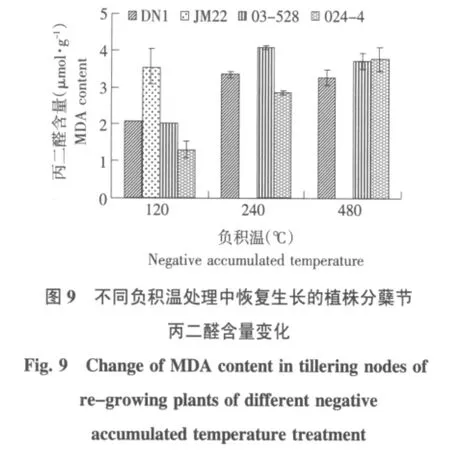
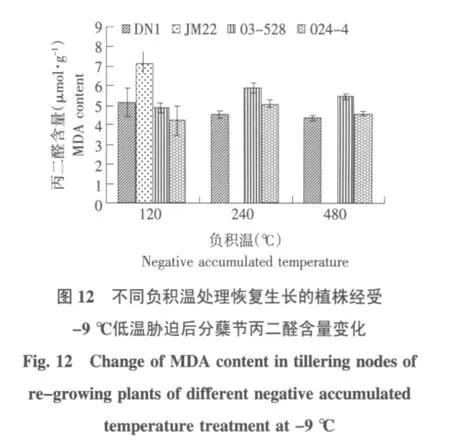
进一步分析不同冷冻处理恢复生长的冬麦植株在经受相同低温胁迫处理时MDA含量的变化,结果见图 9~12。
由图9可知,在未接受低温处理时,负积温120℃冷冻处理的植株恢复生长后,济麦22的MDA含量最高,随着冷冻处理温度的降低,东农冬麦1号、03-528和024-4品系的MDA含量普遍增加,东农冬麦1号和03-528在冷冻负积温480比240℃处理低。由图10可知,-3℃低温胁迫时,负积温240℃冷冻处理恢复生长的东农冬麦1号、03-528和024-4品系3个品种(系)的MDA含量高于另外两个返青温度处理,其中03-528品系差异最显著;负积温480℃冷冻处理恢复生长后,各品种(系)MDA含量为冷冻处理中最低。由图11可知,在-6℃低温胁迫下,随着返青起始温度的下降各品种(系)MDA含量呈现下降趋势;各起始温度处理内部MDA含量差异较小。由图12可知,在-9℃低温胁迫下,负积温120℃冷冻处理恢复生长后,济麦22品种MDA含量最高。不同的低温胁迫下,济麦22和03-528品系的MDA含量普遍高于东农冬麦1号和024-4品系。


3 讨 论
3.1 相对电导率的的变化
质膜是细胞与环境之间的界面,各种逆境对细胞的影响首先作用于质膜,而逆境胁迫对质膜结构和功能的影响通常表现为选择性的丧失,电解质和某些小分子有机物质大量渗漏,从而引起相对电导率的上升[9]。
本试验中,抗寒性较高的东农冬麦1号、03-528和024-4品系在越冬期间相对电导率随着负积温的积累有所上升,但上升幅度显著低于抗寒性弱的济麦22。当负积温累积到480℃时,济麦22相对电导率大幅度上升,达到0.6,说明此时济麦22植株受冻害严重,甚至于死亡。不同负积温处理的植株返青后,在低温胁迫下随着胁迫温度的降低,相对电导率均有不同幅度升高,抗寒性弱的品种升高幅度较大。在-9℃低温胁迫时东农冬麦1号、03-528和024-4品系的相对电导率均接近0.5,这说明在-6℃至-9℃低温胁迫之间可能具有植株耐受低温的临界值。值得注意的是,-3℃和-6℃低温胁迫下,240℃负积温返青处理中各品种的电导率值比其他两个负积温处理的电导率值低,这说明一定程度上的负积温的积累可以提高植株的抗寒性。但随着负积温的逐步累积,低温对植株造成伤害有所加深。
3.2 MDA含量的变化
MDA作为膜脂过氧化终产物,对植物细胞膜系统结构和功能都具有较显著影响。MDA不仅能与蛋白结合从而引起生物膜中结构蛋白和酶的聚合并使它们的结构功能和催化功能发生改变,还可以降低膜中不饱和脂肪酸的含量,从而降低膜流动性[10]。很多研究都证实,当MDA含量积累到一定程度时,相对电导率会显著上升[11-12]。Simon指出膜脂过氧化引起膜透性增加的直接原因可能是磷脂性质的改变,间接原因可能是膜蛋白在过氧化过程中受到伤害。
本试验中,当负积温积累到240℃时,东农冬麦1号和03-528品系的MDA含量升高,而济麦22和024-4品系的含量却下降。此时,济麦22的MDA含量仍然高于其他品种,而024-4品系的MDA含量显著低于其他品种。济麦22品种的MDA含量的变化可能是由于11月初气温突然下降,从而导致越冬初期抗寒性较弱的济麦22膜系统受损并随着温度的逐渐稳定而有所恢复。说明济麦22在遭遇突然降温的情况下,膜脂过氧化反应比其他品种剧烈。024-4品系的MDA含量变化的原因不明,还有待进一步研究。在不同负积温返青处理之间,480℃负积温处理中各品种(系)在不同低温胁迫下均表现出较低的丙二醛含量水平。可以推测,越冬期负积温的累积对膜脂抗氧化具有积极意义。还有一点值得关注的是,无论120℃、240℃,还是480℃负积温的返青处理,给予-3℃、-6℃和-9℃三个低温处理后,丙二醛含量均表现为-6℃处理下最低,显著低于-3℃和-9℃低温处理下的丙二醛含量,这点与不同负积温起始低温处理下电导率的变化结果一致,由此可以得出,冬小麦分蘖节对返青期低温也有一个适应过程,经过-3℃低温胁迫下,抗寒性有所增强,在-6℃低温胁迫时,细胞膜也有所恢复,但是进一步的低温,又会导致膜脂过氧化的加剧。
4 结 论
伴随着植物恢复生长后低温胁迫的加剧,MDA含量呈上升趋势,细胞膜透性增大,相对电导率相应增加。因此,丙二醛和相对电导率可在一定程度上反映冬小麦品种抗寒性的强弱。不同抗寒性冬麦品种(系)植株恢复生长后,在低温胁迫下仍表现出显著差异,抗寒性强的品种(品系)质膜的稳定性大于抗寒性弱的品种(系)。恢复生长的冬小麦对低温也有适应的过程,-3℃低温胁迫下,抗寒性加强,-6℃细胞膜有所恢复,进一步的低温,则膜脂过氧化加剧。
[1]于晶,周子珊,牟永潮,等.低温下不同抗寒性冬小麦叶片组织结构比较[J].东北农业大学学报,2010,41(4):7-11.
[2]于晶,孟健男,曾俨,等.温度及土壤含水量对冬小麦生长习性及返青率的影响[J].东北农业大学学报,2010,41(5):9-13.
[3]陈儒钢,巩振辉,逯明辉.植物抗寒基因工程研究进展[J].西北植物学报,2008,28(6):1274-1280.
[4]万富世.冬小麦品种越冬性改良理论与实践[M].北京:中国农业科学技术出版社,2006:43.
[5]高俊凤.植物生理学实验技术[M].西安:世界图书出版社,2000:35.
[6]赵黎芳,张金政,张启翔.盐和水分预处理对扶藤幼苗抗寒性的影响[J].植物研究,2004,24(3):313-316.
[7]巩振辉.作物育种学[M].北京:农业出版社,2007:45.
[8]刘艳阳,李俊周,陈磊,等.群低温胁迫对小麦叶片细胞膜脂质过氧化产物及相关酶活性的影响[J].麦类作物学报,2006,26(4):70-73.
[9]陈少裕.膜脂过氧化对植物细胞的伤害[J].植物生理学通讯,1991,27(2):84-90.
[10]王连敏,王立志,张国民,等.苗期低温对玉米体内脯氨酸电导率及光合作用的影响[J].中国农业气象,1999,20(2):28-30.
[11]Dobretsov G E,Borschevskaya T A,Petrov V A,et al.The increase of phospholipid bilayer rigidity after lipid peroxidation[J].FEBS Letters,1977,84:195.
[12]周龙,廖康,王磊,等.低温胁迫对新疆野生樱桃李电解质渗出率和丙二醛含量的影响[J].新疆农业大学学报,2006,29(1):47-50.
[13]Simon E W.Phospholipids and plant membrane permeability[J].New Phytol,1974,73:377.
