相对密度和季节对菲牛蛭Hirudinaria manillensis繁殖习性的影响*
2010-06-05储霞玲梁万文吕军仪
张 彬,储霞玲,林 强,梁万文,吕军仪
(1. 广西水产研究所,广西 南宁 530021;2. 中山大学生命科学学院,广东 广州 510275;3. 中国科学院南海海洋研究所,广东 广州 510275)
菲牛蛭(HirudinariamanillensisLesson, 1842)是国际上一种重要的医用水蛭,具有抗凝、抗血栓、纤溶、降血脂和抗炎功能,主治女子月闭、跌打损伤、产后血晕及与血栓形成有关的心脑血管疾病[1]。
近年来,由于环境污染和生态平衡的破坏以及人类的酷捕滥捞,药用蛭类资源锐减,已日趋枯竭[2-4]。正因为如此,北美和欧洲的重要医用水蛭——欧洲医蛭已被国际上一些权威机构所重视[4-5]。为了满足医学临床、传统中药和其他科学研究的需要,水蛭的人工养殖也作为一个新兴的产业受到重视,中国、俄罗斯、英国、法国和土耳其等国家已经有相当规模的水蛭养殖业,并取得了较好的经济和社会效益[6]。自然界中,随着种群相对密度的增加,对资源、空间的竞争加剧,引起动物摄食、生长、能量代谢、行为、生理以及免疫功能的一系列变化,最终可能使动物的生存能力降低[7]。因此,在人工养殖环境中,相对密度是动物种群生态学研究的一个重要内容。另外,水蛭的繁殖习性具有明显的种属特异性,如生活在温带气候的日本医蛭必须通过1-3月份的越冬期才能繁殖,且时间集中在4-7月份[1];而亚热带蛭种海南山蛭的繁殖时间为3-11月份[8-9],周期明显比日本医蛭延长;菲牛蛭是广泛分布于东南亚的吸血类水蛭,生活的地理环境与海南山蛭相似[10-11],具有一年多次繁殖的可能性,并且不同季节下的繁殖效率也可能存在差异。
目前国内外对菲牛蛭研究的文献报道主要集中在分类、生态调查和生化成分以及临床应用方面,而有关人工养殖下的繁殖生物学尚未见报道。因此,我们在人工养殖条件下,研究春夏和秋冬不同季节中,相对密度对菲牛蛭繁殖的影响,以探讨在华南地区一年多季繁殖的可能性及差异性,以期为其大规模人工繁殖提供数据参考。
1 材料与方法
1.1 实验材料
实验用健康菲牛蛭亲本(F2)由广州市安圣药源生物开发有限公司提供,原种为华南热带山区野生种。
1.2 养殖管理
亲蛭暂养于育苗池(2.0 m × 1.0 m × 0.8 m, 长×宽×高),水体深度为0.4 m,池底层平铺一层光滑的河卵石,养殖用水为充分曝气的去氯自来水,暂养20 d后入繁殖箱。暂养时间为2006年7月23日至8月12日(秋冬季节繁殖用)和2007年3月1日至3月21日(春夏季节繁殖用)。采用充氧泵及时补充水体溶氧量,每隔3天换水一次,每次换水量为50%。水体温度由电热棒加热调节。水温、溶解氧(DO)、pH和光照分别是(28±0.5) ℃,(7.6±0.2)mg·L-1,7.5±0.2和1 000~1 500 lx;整个亲蛭暂养过程中,氨氮含量控制在0.08 mg·L-1以下。进行繁殖实验之前,亲蛭投喂2次,每次间隔时间为10 d。
1.3 实验设计
设置4个相对密度梯度[5, 20, 30, 50尾/箱(0.05 m3土层)],每个处理组设5个重复箱。从暂养池中精心挑选个体大、体格健壮、无伤病以及生殖环带明显的用于试验。亲蛭平均体质量为(10.51±1.68) g,各实验组间的亲蛭体质量差异不显著(ANOVA,P>0.05)。繁殖实验于三面透光的玻璃瓦大棚内进行,自然变温。繁殖基质为偏酸性壤土,取自广州市郊有菲牛蛭生活的稻田附近,经检测无农药和化肥残留。繁殖箱体规格相同(0.5 m× 0.5 m× 0.5 m, 长×宽×高)。秋冬季节繁殖实验:期间平均气温为(23.03±6.60) ℃;土壤pH、相对湿度和光照分别是6.9±0.2,(80±5.0)% 和1 000~1 500 lx;繁殖时间为2006年8月12日至2006年12月12日。春夏季节繁殖实验:期间平均气温为(26.95±6.56) ℃;土壤pH、相对湿度和光照分别是6.9±0.5,(78±5.0)%和2 000~3 000 lx;繁殖时间为2007年3月21日至2007年6月2日。
1.4 数据统计与分析
实验参量为亲蛭的产茧率和平均产茧数,卵茧孵化率和孵化数、亲蛭死亡率以及卵茧大小和湿质量;对不同季节下卵茧孵化时间和孵化规律进行观察记录。孵化时间为从产茧时间到孵化结束时间。孵化期间,每天收集并计数孵出幼蛭的尾数,据此得出孵化规律。随机收集20枚未孵化的新产卵茧,测量并记录各个卵茧的长度、直径和湿质量。




数据由平均数±标准差(mean±S.D.)表示,采用单因子方差分析(One-Way ANOVA)。如果差异显著(P<0.05),再进一步以Duncan法进行多重比较。所有数据均采用SPSS 11.5(Statistical Program for Social Sciences.11.5)统计软件进行统计分析。
2 结 果
2.1 秋冬季节,相对密度对菲牛蛭繁殖的影响
秋冬季节,亲蛭产茧率和产茧数在各相对密度组间差异不显著 (P>0.05);而卵茧孵化率和孵化数在各相对密度组间存在显著差异 (P<0.05),高相对密度组(50尾·箱-1)所产卵的孵化率和孵化数最低,显著低于其他相对密度组(P<0.05);相对密度对亲蛭死亡率无显著影响 (P=0.247),各个相对密度组中的亲蛭死亡率均低于10%;卵茧长和直径在各个相对密度组间均存在显著差异 (P<0.01),最大茧长出现在相对密度组(5尾·箱-1),最大茧直径出现在相对密度组(30尾·箱-1),而高相对密度组(50尾·箱-1)卵茧个体最小,均显著低于其他相对密度组(P<0.01);此外,卵茧湿质量在各相对密度组间也存在显著差异 (P<0.01),其中最大茧质量出现在相对密度组(30尾·箱-1),达(0.98±0.21) g,而高相对密度组(50尾·箱-1)湿质量最小,仅为(0.57±0.18) g,显著低于其他相对密度组(P<0.01)(表1)。秋冬季节各相对密度组卵茧孵化趋势无明显规律性。各相对密度组的卵茧孵化时间均为100 d,长达3个多月,孵化过程断断续续,周期长,无同步性(图1)。
2.2 春夏季节,相对密度对菲牛蛭繁殖的影响
春夏季节,产茧数在各相对密度组间存在显著差异 (P<0.01),其中相对密度组(5尾·箱-1)最高,达3.84±0.26枚,而产茧率在各相对密度组间差异不显著(P=0.187);孵化率在各相对密度组间存在显著差异 (P<0.01),其中相对密度组(5尾·箱-1)最高,达(96.82±2.92) %,而孵化数在各相对密度组间差异不显著 (P>0.05);相对密度对亲蛭的死亡率具有显著影响 (P<0.01),随着相对密度升高,亲蛭死亡率显著上升,相对密度组(50尾·箱-1)最高,达(29.60±5.55)%,而其他相对密度组的亲蛭死亡率均低于10%;相对密度显著影响亲蛭产卵茧长和直径,均随着相对密度的升高而逐渐降低;此外,相对密度对茧湿质量也存在显著影响 (P<0.05),随着相对密度的升高而逐渐降低(表2)。
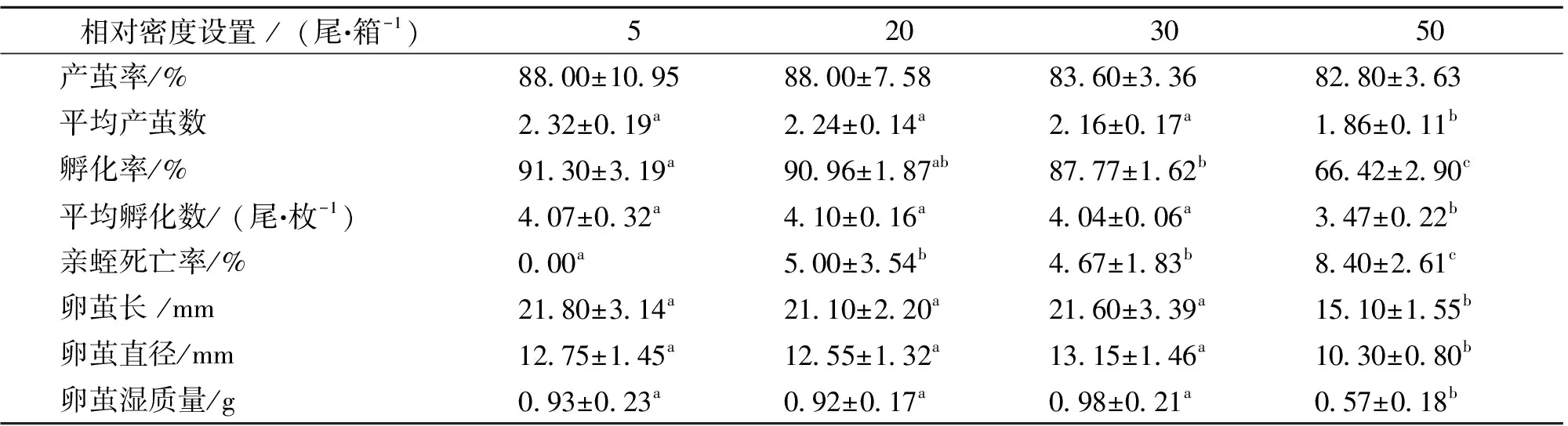
表1 秋冬季节,不同相对密度下菲牛蛭繁殖指标的比较1)
1)表中数据由平均值±标准差表示;每行数据右上方的字母不同表示有显著性差异(P<0.05)
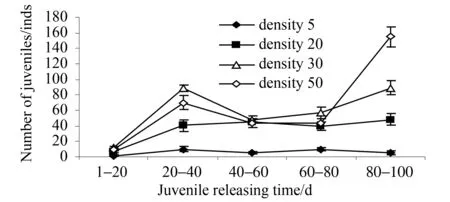
图1 不同相对密度下幼蛭孵化时间和孵化趋势(秋冬季节)
春夏季节各个相对密度组实验中,幼蛭的孵化趋势一致,孵化时间均为44 d左右。同时观察到在幼蛭整个孵化周期,存在5个时期,分别为Ⅰ(0407-0413),Ⅱ(0413-0419),Ⅲ(0419-0427),Ⅳ(0427-0504)和Ⅴ(0504-0521)。其中,Ⅱ(0413-0419)和Ⅳ(0427-0504)是孵化的两个高峰期,在这两个时期孵化出的幼蛭数占总量的85%以上(图2)。

图2 不同相对密度下幼蛭孵化时间和孵化趋势(春夏季节)
表2 春夏季节,不同相对密度下菲牛蛭繁殖指标的比较1)
Table 2 Comparisons of condition indices ofH.manillensisLesson at different broodstock densities in spring and summer seasons
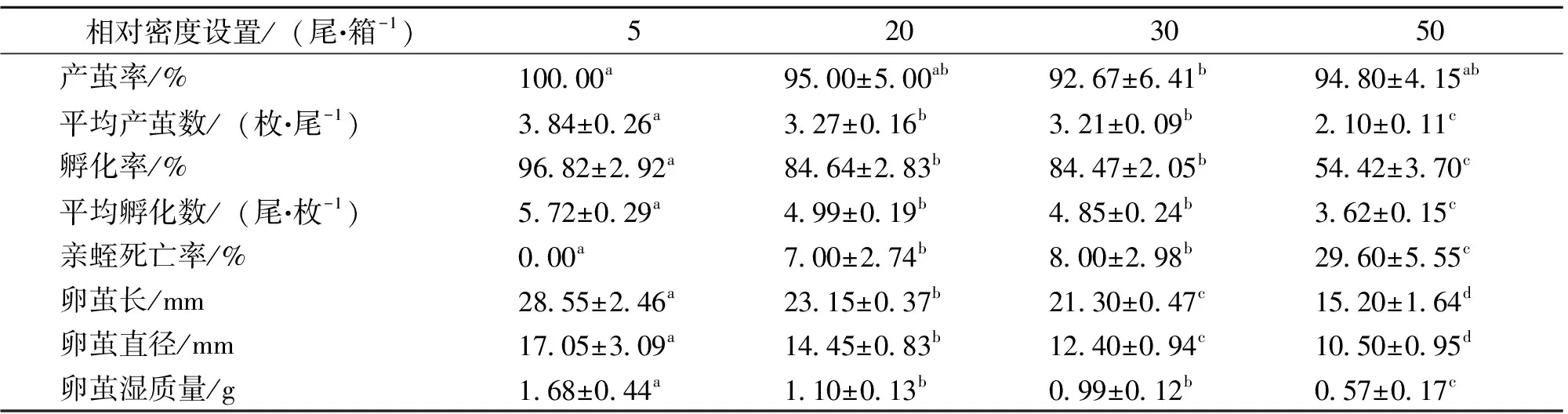
相对密度设置/(尾·箱-1)5203050产茧率/%100.00a95.00±5.00ab92.67±6.41b94.80±4.15ab平均产茧数/(枚·尾-1)3.84±0.26a3.27±0.16b3.21±0.09b2.10±0.11c孵化率/%96.82±2.92a84.64±2.83b84.47±2.05b54.42±3.70c平均孵化数/(尾·枚-1)5.72±0.29a4.99±0.19b4.85±0.24b3.62±0.15c亲蛭死亡率/%0.00a7.00±2.74b8.00±2.98b29.60±5.55c卵茧长/mm 28.55±2.46a23.15±0.37b21.30±0.47c15.20±1.64d卵茧直径/mm17.05±3.09a14.45±0.83b12.40±0.94c10.50±0.95d卵茧湿质量/g 1.68±0.44a1.10±0.13b0.99±0.12b0.57±0.17c
1)表中数据由平均值 ± 标准差表示;每行数据右上方的字母不同表示有显著性差异(P<0.05)
2.3 不同季节对菲牛蛭繁殖的影响
季节对菲牛蛭的繁殖具有显著影响(表1, 2)。以中间相对密度组(20尾·箱-1)为例,比较不同季节的繁殖指标,除产茧率和亲蛭死亡率差异不显著外,其它指标在春夏季节均显著高于秋冬季节(P<0.01)(表3)。同时卵茧的孵化时间和孵化趋势在春夏季节要明显优于秋冬季节,首先孵化时间在春夏季节为44 d,远远短于秋冬季节的100 d;其次是孵化趋势,春夏季节的孵化曲线一致,时间集中,有两个明显的孵化高峰,在这两个高峰期孵化出的幼蛭数占总量的85%以上,而秋冬季节孵化曲线凌乱、分散,无规律性且时间明显延长(图1, 2)。
3 讨 论
相对密度是水蛭繁养殖过程中的重要环境因子,能显著影响水蛭的生长[1, 12]。本研究结果表明相对密度能显著影响菲牛蛭的繁殖效率,相对密度升高对亲蛭的产茧数产生明显负面影响。秋冬和春夏季节中,相对密度组(5尾·箱-1)均为最佳的产茧相对密度。高相对密度下亲蛭产茧力低与其生存环境有关,单位养殖空间里,相对密度的升高,使得对食物和空间的竞争加剧,产生生存压力,从而干扰了其正常的繁殖行为。由于菲牛蛭营群居生活,高相对密度繁殖时,30~40 d,亲蛭排泄物将占体质量的1/3,使土壤基质迅速处于富营养化状态,给亲蛭的生存造成危害。因此,在繁殖过程中,要定期清理箱底部被排泄物污染的土层,并加入经消毒处理的新土层,给菲牛蛭繁殖提供适宜的环境条件。相对密度对菲牛蛭的交配率无显著影响,这个结果与我们挑选的菲牛蛭亲蛭均已性成熟有关。性成熟个体由于生殖腺发育成熟而变大,使生殖环带区变大,外表可见环带区灰色的生殖腺组织[13],当菲牛蛭头部稍后出现棕红色或淡红色戒指状生殖带腺体,说明该个体已经交配。对菲牛蛭而言,平均寿命为5~6 a,而2龄是其生命中的繁殖高峰期[6, 10]。本文实验材料采用2龄F2代亲本,性成熟个体挑选依据上述特征判断,所选亲本个体大且均匀,发育状态一致。正是由于相似的生理指数,才使得各个相对密度间产茧率差异不显著,这与Holmstrand and Collins[14]关于Nephelopsisobscura的研究一致。

表3 不同季节下,菲牛蛭繁殖性能的比较(以相对密度组(20)的繁殖指标为参照)1)
1)表中数据由平均值±标准差表示;每行数据右上方的字母不同表示有显著性差异(P<0.05)
水蛭的生长、发育和繁殖时间受温度的影响显著[8-9]。日本医蛭在平均气温13.1~15.4 ℃时开始交配,产茧的平均气温为19.2~21.3 ℃,孵化的平均气温为21.3~32.1 ℃[1]。在本研究中,秋冬季节的繁殖周期长达122 d,期间的室内气温为(23.03 ± 6.60) ℃;而春夏季节仅需73 d,期间的室内气温为(26.95 ± 6.56) ℃。这两个季节下的平均气温均在已报道的菲牛蛭正常生长、发育和繁殖温度 (19~32 ℃) 范围之内[10]。本实验中,相对密度显著影响卵茧的孵化率和孵化数,不同季节中,最高的孵化率和孵化数均出现在低相对密度组5。谭恩光等[8]报道海南山蛭的孵化率和孵化数分别为77.4%和6~17尾,其中孵化率明显低于本实验结果(平均超过80%),而孵化数略高于本实验结果(平均低于4.5尾),这种差异可能是由于物种本身以及繁养殖环境不同而造成的。
相对密度能显著影响亲蛭产茧的大小和湿质量,低相对密度组的卵茧大小和湿质量要明显大于高相对密度组,这个结果在不同季节是一致的。所不同的是,春夏季节的卵茧大小和湿质量相比之下要大于秋冬季节,这是季节对菲牛蛭繁殖能力影响的一种表现。谭恩光等[10]报道了菲牛蛭产卵茧的平均长和直径分别为22 mm和13 mm,这与本研究中的数据相吻合。研究表明,水蛭卵茧的湿质量在不同蛭种间差异显著,如宽体金线蛭的卵茧湿质量为1.6~2.0 g[14-15],而海南山蛭的卵茧湿质量为0.15~0.18 g[8]。本实验研究中,菲牛蛭的卵茧湿质量为0.57~1.68 g,个体差异悬殊,说明蛭体大小和繁殖环境对菲牛蛭的产茧性能有显著影响。
水质、温度和疾病均显著影响蛭类的存活[11, 16-17],同时水蛭的年龄也是其存活力的一个重要因子[12]。本研究结果表明,繁殖相对密度也能显著影响水蛭亲本的存活率,随着相对密度的升高,亲蛭死亡率逐渐上升。通过大量的实验观察,可以推断产生这种结果的原因主要是由于高相对密度加剧了菲牛蛭生存空间的竞争,伴随着环境因子的变化,产生大量有毒污染物,从而导致亲蛭的高死亡率。春夏季节的亲蛭死亡率要高于秋冬季节,这说明温度对其存活率影响是显著的。
本研究中,菲牛蛭在春夏季节的孵化时间是44 d,而在秋冬季节长达100 d,不同季节中的温度,尤其昼夜温差是产生这个结果的原因。在长期低温以及温差大的环境下,卵茧内的胚胎发育受阻,发育时间延长,有些卵茧中的幼蛭虽已发育完全,但迟迟不破茧而出,导致孵化时间明显延长。谭恩光等[10]报道了菲牛蛭H.manillensis卵茧的孵化时间为26 d,这与本研究结果相差很大,我们分析产生这种差异的原因是由不同的孵化条件造成的。另外,谭恩光等的实验对象是实验室内同时产下的几枚茧,而本实验研究对象是一批菲牛蛭亲本在一个繁殖季节中产下的所有卵茧,这种实验设计上的不同,推测也是产生孵化时间大相径庭的原因。
海南山蛭是我国海南省特有蛭种,野外生活在热带原始森林里,室内饲养的海南山蛭产茧季节为4-11月份,这与室内维持较高的温度和湿度有关,在野外条件下,繁殖季节为6-11月份,4月份海南省处于高温干旱季节,山蛭一般不活动或少活动,5月份进入雨季,山蛭大量活动,6月份开始产卵繁殖,12月份温度太低,且雨量少,不能繁殖,8-10月份是海南山蛭的繁殖高峰期[8]。可见海南山蛭具有较长的繁殖季节,菲牛蛭和海南山蛭同属热带吸血蛭类,在摄食和繁殖习性上十分相似,本研究中菲牛蛭的一年多季繁殖成功很大程度上归功于其相似的生活环境和繁殖特性。而另一种吸血蛭类日本医蛭Hirudonipponia生活在长江中下游,其对冷比对热更能适应,通常在水温15~25 ℃最理想,只有处在恒定的温度下交配与产卵的周期才能较长,日本医蛭只有在低温下(4~12 ℃)蛰伏越冬1~3个月的成熟个体才能交配和产茧,高温的夏天和低温的冬天是不能繁殖后代的[1]。然而,本研究结果证实,菲牛蛭在华南地区进行反季节繁殖是成功的,即1年可多次繁殖。菲牛蛭是热带亚热带种,其对冷比对热更敏感,华南地区相对北方地区温差波动较小,也给菲牛蛭的多季繁殖创造了有利温度条件,因此物种本身的生理差异和地理因素是产生上述研究结果分歧的原因。
致谢:本试验得到了广州市安圣药源生物开发有限公司吴川和龚元的大力帮助,在此一并表示感谢。
参考文献:
[1] 刘岱岳, 余传隆, 刘鹊华. 生物毒素开发与利用[M]. 北京:化学工业出版社, 2007: 337-340.
[2] ELLIOTT J M, TULLET P A. The status of the medicinal leech.Hirudomedicinalisin Europe and especially in the British isles[J]. Biological Conservation, 1984, 29(1): 15-26.
[3] SAWYER R T. Why we need to save the medicinal leech?[J]. Oryx, 1981, 16: 165-168.
[4] WELLS S, COOMBES W. The status of and trade in the medicinal leech[J]. Traffic Bull, 1987, 8: 64-69.
[5] WELLS S M, PYLE R M, COLLINS N M. The IUCN Invertebrate Red Data Book[M]. Cambrige, UK: IUCN. The World Conservation Union, 1983.
[6] 杨潼. 中国动物志环节动物门蛭纲[M]. 北京: 科学出版社, 1996: 117-126.
[7] SCHRECK C B, SANCHEZ W C, FITZPATRICK M S. Effects of stress on fish reproduction, gamete quality, and progeny[J]. Aquaculture, 2001, 197, 3-24.
[8] 谭恩光, 陈晶, 钱月桃. 海南山蛭HaemadipsahainanaSong. et al的生长摂食和生殖[J]. 海南大学学报: 自然科学版, 1992, 10(1): 16-23.
[9] 谭恩光. 海南山蛭对土壤湿度和气温的适应[J]. 生态学报, 2001, 21(3): 458-461.
[10] 谭恩光, 黄立英, 关莹, 等. 广东菲牛蛭生长和生殖的研究[J]. 中草药, 2002, 33(9): 837-840.
[11] 谭恩光. 广东菲牛蛭生活水体的化学环境[J]. 中草药, 2005, 36(10): 1561-1563.
[12] MANN K H. The breeding, growth and age structure of a population of the leechHelobdellastagnalis(L.) [J]. The Journal of Animal Ecology, 1957, 26(1), 171-177.
[13] COLLINS H L, HOLMSTRAND L L. Indicators of sexual maturity in the leech,Nephelopsisobscura(Annelida: Hirudinea)[J]. American Midland Naturalist, 1984, 112(1): 91-94.
[14] HOLMSTRAND L L, COLLINS H L. Reproductive biology of the Erpobdellid leechNephelopsisobscuraVerrill, 1872 in culture[J]. Freshwater Invertebrate Biology, 1985, 4(1): 30-34.
[15] 史红专, 刘飞, 郭巧生. 温度对蚂蟥生长及摂食规律影响的初步研究[J]. 中国中药杂志, 2006, 31(23): 1944-1946.
[16] 史红专,刘飞,郭巧生. 温度和体质量对蚂蟥人工繁殖影响的研究[J]. 中国中药杂志, 2006, 31(24): 2030-2032.
[17] SAWYER R T. Observations on the natural history and behavior ofErpobdellapunctata(Leidy) (Annelida: Hirudinea)[J]. American Midland Naturalist, 1970, 83(1): 65-83.
