森林固碳领域的研究方法及最新进展
2010-06-04殷鸣放殷炜达毕刚蕊张艳会李智伟谭希斌
殷鸣放,杨 琳,殷炜达,毕刚蕊,张艳会,李智伟,谭希斌
(1. 沈阳农业大学 林学院,辽宁 沈阳 110866;2. 东京大学新领域创成研究科自然环境系,柏 277-8563;3. 黑龙江省第二森林调查规划设计院,黑龙江 绥化 152000;4. 辽宁省锦州市林业局,辽宁 锦州 110036)
全球碳循环是碳元素在地球各个碳库之间的不断交换、循环周转过程,它是地球化学循环中极为重要的组成部分。随着全球变化与陆地生态系统关系研究的深入,以及联合国气候变化框架协议的实施,陆地生态系统碳储量和碳平衡已成为国际生态学研究的热点。
陆地生态系统中,森林是碳循环的主体,森林面积占全球陆地面积的 27.6%,森林植被的碳贮量约占全球植被的 77%,森林土壤的碳贮量约占全球土壤碳储量的 39%。森林生态系统碳储量占陆地生态系统碳储量的46.6%左右[1]。赵敏等根据各省、市的针叶林和阔叶林蓄积量资料,估计出中国森林植被碳储量约3 788.1Tg[2]。森林的生态功能包括防风固沙、保持水土、涵养水源、保护生物多样性、提供森林游憩和多种林产品。森林所具有的一般生态功能长期以来已经成为科学家研究的重点,然而森林碳汇功能直到近些年来才被社会和诸多学者重视和研究。森林在陆地生态系统中具有巨大的碳储存能力,增加森林的碳汇量是世界公认的最经济有效的减缓CO2浓度上升的有效方法。森林碳循环是全球变化与陆地生态系统(GCTE)关系研究的重点内容之一。
因此,科学地计量与评估森林生态系统的固碳能力和碳储量,特别是快速准确地计量与评估森林生态系统的碳储量变化速率具有极其重要的科学意义与现实意义。本文试图通过对森林固碳研究领域国内外使用的几种碳汇计量方法及其现状的资料整理,介绍森林固碳研究领域的最新进展,展望未来森林固碳研究的发展方向。
1 森林碳汇
在全球变化与陆地生态系统的研究中,全球碳循环问题即温室气体的“源”与“汇”问题,以主要温室气体——CO2的源汇为主要研究问题。《联合国气候变化框架公约(UNFCCC)》中将碳源定义为向大气中释放碳的过程或活动。动植物的呼吸作用、动植物本身的分解、化石燃料燃烧、大规模森林破坏、土地利用形态的改变,均形成大量的碳排放到大气中。而碳汇一般是指从空气中清除CO2的过程、活动、机制。简单地说,当生态系统固定的碳量大于排放的碳量,该系统称为大气中 CO2的汇,简称碳汇,反之,则为碳源[3]。由于森林具有储存碳的“库”的作用,因此可以形象的将森林看作是储存碳的“碳库”。林业碳汇,是指通过实施造林、再造林和森林管理,吸收大气中的CO2并将其固定在植被或土壤中,从而减少大气中CO2的浓度的过程和活动。
森林在陆地生态系统与大气碳库循环和碳交换过程中具有主导地位,处于良好经营状态下的森林可以极大地提高森林碳库的碳吸收速度和碳吸收能力。合理轮伐以及木材的合理利用可以延伸森林碳汇作用,从而更好地发挥森林的碳汇效果。
森林生物量约占全球陆地植被生物量的 90%[4],它是森林固碳能力的重要标志和评估森林碳收支的重要参数[5]。森林生物量的变化反映了森林的演替、人类活动、自然干扰(如林火、病虫害等)、气候变化和大气污染变化等影响,是度量森林结构和功能变化的重要指标[6]。
我国森林生物量测定最早是由潘维涛、冯宗炜等人[7~8]于20世纪70年代末、80年代初开始进行的,目前相关研究资料较为丰富,但由于各研究者所侧重的领域有所不同,在研究地点、研究方法及森林类型等方面也存在差别,使得碳储量的研究结果有所差异。当时对全国森林植被生物量进行了测定,随后开展了大区域森林生物量的模拟研究,结果表明,对于某一森林类型,森林的生产力和生物量与森林自身的生物学特性(蓄积量、林龄等)有着密切的联系[8]。因此,充分合理利用森林资源清查资料,结合已有的研究结果,可以提出评估不同类型森林碳收支的技术和方法,利用森林资源清查资料、模型和遥感数据也可从不同角度对我国森林生态系统的碳储量和碳密度进行分析[9~11]。
2 森林固碳的研究方法及其现状
2.1 样地清查法
样地清查法是指通过设立典型样地,准确测定森林生态系统中植被、枯落物或土壤等碳库的碳储量,并通过连续观测来获得一定时期内的碳储量变化情况的推算方法[12~13]。它主要可分为3类:平均生物量法[14]、生物量转换因子法[15~16]和生物量转换因子连续函数法。这3种计算方法由同样数学推理方法得来。还有研究者将样地清查法分为生物量法、蓄积量法及生物量清单法。这些方法与之前提到的3种方法虽然存在名称、定义、计算步骤上的差别,但是在原理上基本没有区别,因此本文详细介绍前面提出的3种方法。
2.1.1 平均生物量法 生物量是指一个有机体或群体在一定时间内积累的有机质总量,森林生物量通常以单位面积或单位时间积累的干物质量或能量来表示[17]。平均生物量法是指基于野外实测样地的平均生物量与该类型森林面积来求取森林生物量的方法[18]。该方法的优势主要体现在直观明确,操作简便以及节约成本。获得样地林分平均生物量的方式主要有3种,即皆伐法、相关曲线法和标准木法。
皆伐法是将单位面积上的林木逐个的伐倒后测定其各部分(树干、枝叶、果和根系等)的鲜质量,并换算成干质量,将各部分质量合计,即为单株树木的生物量,将单株树木的生物量累加后除以相应的株数即可得到平均生物量[19]。相关曲线法即在研究区域内选取样木,伐倒后按器官称重,然后根据各器官生物量与某一测树指标间的相关关系进行回归拟合,建立回归曲线方程,以实测的胸径、树高推算林分生物量。目前使用的主要回归关系类型为:

式中:W为各部分的生物量,D为胸径,H为树高,a、b为方程中的待估参数。
标准木法是根据每一块标准地上的标准木推算林分生物量,用标准木各组分(干、枝、叶、根)的生物量乘以该标准地的树木株数。这种方法由于实测资料的取样点较少,导致森林生物量估计偏大[20~21]。而且在应用平均生物量进行野外测定时,人们大都选择生长良好的林分,用平均生物量来推算该森林类型的总生物量,结果偏大是不难想象的[22]。
2.1.2 生物量转换因子法 生物量转换因子法(Biomass Expansion Factor,BEF)又叫做材积源生物量法(Volume derived biomass),是利用林分生物量与木材材积比值的平均值乘以该森林类型的总蓄积量或利用木材密度(一定鲜材积的烘干量)乘以总的蓄积量和总生物量与地上生物量的转换系数得到该类型森林的总生物量的方法[23]。在森林生物量的组成当中,树干(材积)只是森林生物量组成中的一部分,其所占的比率依据树种和立地条件不同会有很大的差异[16]。因此,为了推算某一树种的生物量,就要知道树干占总生物量的比例,即知道根、茎、枝、叶等部分的生物量以计算出树干的比例。由于树干的生物量与其它器官的生物量存在着很强的相关关系,所以用树干材积推算森林总生物量是可行的[24]。
生物量转换因子法的不足主要反映在生物量转换因子,包括木材密度(WD,woodensity)和总生物量与地上生物量的转换系数均取作常数。有学者指出,湿润地区、干旱和半干旱地区没有受干扰的原始森林的木材密度分别为:1.74、1.95和1.57;总生物量与地上生物量的转换系数为1.6[25]。但诸多研究表明,对于某一特定的森林类型而言,生物量转换因子是立木的生物量和蓄积量的集中体现,与林木的年龄、种类组成、其它生物学特性和立地条件等密切相关。海南岛热带山地雨林林分生物量的研究表明,生物量转换因子法估算的生物量较皆伐法高20% ~ 40%,而基于实测资料建立的生物量回归模型的估计值与皆伐法非常相近,相对误差在±10%以内[26]。林分生物量和蓄积量与森林类型、年龄、立地条件和林分密度等诸多因素有关[27]。由此可见,采用常数的生物量转换因子不能十分准确地估算森林生物量。
2.1.3 生物量转换因子连续函数法 研究表明,某森林类型的林分生物量与木材材积比值(BEF)不是不变的,而是随着林龄、立地、个体密度、林分状况等不同而变化的[28]。因此林分材积综合反映了林龄、立地、个体密度和林分状况等因素的变化,可以作为换算因子的函数,以表示 BEF的连续变化。生物量转换因子连续函数法利用幂指函数来表示BEF与林分材积(V)的关系:
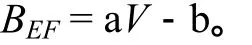
式中,a和b均为大于0的常数。
然而,这种关系难以实现由样地调查到区域推算尺度的转换,即对于由实测资料建立的 BEF值与材积之间的关系推广到处理大尺度的森林资源清查资料时,存在着严重数学推理问题。所以将其更正成利用倒数方程来表示BEF与林分材积之间的关系[10],即:
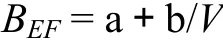
式中,a和b均为常数。也可表示成生物量和蓄积量的简单线性关系[29]:
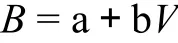
式中,a和b均为常数;B代表生物量,V代表蓄积量。
而如果将估算模型中的系数a当成蓄积量的函数[30],既能克服将生物量与蓄积量之比作为常数的不足,又避免将林分在任一个生长阶段的生物量随蓄积量的变化简单地处理为线性增加的关系。
2.2 微气象学方法
2.2.1 涡度相关法 涡度相关法是一种直接测定植被与大气间 CO2通量的方法,主要是在林冠上方直接测定CO2的涡流传递速率,从而计算出森林生态系统吸收固定CO2量[31]。这是目前测定地气交换最好的方法之一,也是世界上CO2和水热通量测定的标准方法,已被广泛的应用于估算陆地生态系统中物质和能量的交换。该方法可测得生态系统长期或短期的环境变量,使人类可定量理解生态系统中CO2的交换过程,能更深入了解气候变化对生态系统所造成的影响。涡度相关法可以为土壤—植被—大气之间的物质、能量交换模式提供一种直接验证的手段,可通过涡度相关技术所测得的碳通量来推算某一地区的净初级生产力和蒸发量。
涡度相关法所需的关键设备一般是由灵敏度较高的三维超声风速仪、开/闭路式红外CO2/H2O气体分析仪以及温、湿度计等精密仪器组成[32]。涡度相关技术的使用条件是:下垫面平坦均一;大气边界层内湍流间歇期短;研究对象一般在水平均匀的大气边界层内。影响通量观测的因素:①CO2的储存效应。大气比较稳定或湍流作用比较弱时,从土壤和叶片扩散的CO2不能达到仪器测定高度从而造成传感器所测值不包含传感器下方的CO2通量,一般来说,储存项对于低矮植物来说是很小的,但对于较高的森林由于仪器高度以下的气体体积比较大且湍流比较弱,所以储存效应对其测定有较大的影响;②CO2水平平流效应。所测下垫面有一定的粗糙度和地形起伏,或者是所测的源或汇的表面发生变化,这些因素容易引起气流的平流效应;③CO2通量漏流。当大气比较稳定,且地形有一定坡度时容易使空气中的CO2发生漏流。湍流作用比较弱、风速低时,一部分CO2不会通过冠层和大气的交界面。
大气中物质的垂直交换往往是通过空气的涡度状流动来进行的,这种涡度带动空气中不同物质包括CO2向上或者向下通过某一参考面,二者之差就是所研究的生态系统固定或放出的CO2量。其计算公式为:
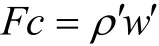
式中,Fc是CO2通量,ρ是CO2的浓度,w是垂直方向上的风速。字母的右上标(小撇)是指各自平均值在垂直方向上的波动即涡度波动,上横线是指一段时间(15 ~ 30 min)的平均值。
Euroflux实验室的科学家应用涡度相关法集中研究了不同纬度欧洲森林的CO2通量的变化[33]。应用涡度相关技术对热带森林、温带森林和北方森林的季节变化模式的研究表明,热带森林全年都表现出净碳汇,而高纬度地区的森林则在生长季节为汇,在冬季则为源[34]。刘允芬等用该方法对千烟洲人工针叶林生态系统的碳通量进行分析,得出该生态系统全年各个月都为碳汇,但碳储量各月之间变化明显[35]。应用此方法对长白山阔叶红松林森林大气界面的CO2湍流交换特征进行了研究,发现森林大气的CO2通量在夜间存在低估的情况[36]。
涡度相关法的特点是直接对森林与大气之间的通量进行计算,能直接对森林生态系统进行CO2通量测定,同时因能够为其他模型的建立和校准提供基础数据而闻名。但是,这一方法需要较为精密的仪器,这些仪器在使用上都有严格的要求。在数据处理上,还需要数学方法和能量守恒闭合等多种方法的校正和数据质量控制,才能得到满意的结果。
2.2.2 驰豫涡旋积累法 驰豫涡旋积累法(Relaxed Eddy Accumulation,REA)起源于涡旋积累法,其基本思想是根据垂直风速的大小和方向采集两组气体样本进行测量。这一技术直到在涡旋积累的思想中引入驰豫的思想,使得不定时采样转换为定时采样,演变成驰豫涡旋积累法。近年来,这一方法已应用到森林CO2通量的计算。这一方法需要用维声速风速仪、红外线CO2分析仪、快速反应螺旋管阀门、数据比较器、数据记录仪、导管系统以及空气泵等。数据比较器用于比较从声速风速仪所得到的即时垂直风速信号与数据记录仪所得到的一定时间的平均值,通过这种比较,数据记录仪就可以估计涡旋是上行还是下行,继而开通或关闭连接2个空气收集袋的阀门。通过数据记录仪的程序化设计,红外线CO2分析仪间隔一定时间(3 min)开启或关闭其通道即可以连续监测2个收集袋内CO2的浓度。林冠层的CO2通量可以通过下式计算:
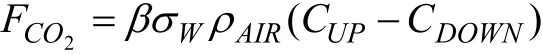
式中,FCO2是CO2通量,是一个半经验常数(0.56);w是垂直风速的标准差;ρAIR是空气的浓度;CUP- CDOWN是0.5 h中2个收集袋的CO2浓度平均值之差。
这一方法在CO2通量测定中有应用,但并不是CO2通量网所使用的标准方法。驰豫涡旋积累法因为其能够收集气体并能够进行长期监测,所以在一些痕量易挥发有机化合物气体的通量测定上更有优势。运用驰豫涡旋积累法对森林CO2通量进行研究在中国还未见报道。
2.2.3 箱式法 箱式法是用来测定土壤和植物群落的微量气体成分排放通量以测定碳储量的方法。其基本思想是:植被的一部分被套装在一个密闭的测定室内,在封闭系统内,CO2浓度随时间的变化就是CO2通量,即:
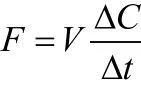
式中,V是测定室的体积;△C/△t是CO2的变化速率。
在一个开放系统时,测定室内有恒定速率(d)的空气流过,通过测定流入与流出测定室的空气中CO2浓度差'''CΔ来计算CO2通量:
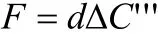
目前,根据以上基本思想已经开发出先进的仪器设备。主要的代表公司是英国ADC公司和美国Licor公司生产的光合测定系统,其中Licor公司生产的Li6400便携式光合测定系统可以测定植物不同器官、土壤的光合或呼吸速率,也可以稍加改进测定树干的呼吸、最大羧化速率、电子传导速率、最大量子利用效率等指标,这些指标对于估计树冠光合作用是必不可少的。然而,ADC系统和Licor系统的一个缺陷就是不能够长期自动进行观测,这导致在估测整个生态系统CO2通量时产生较大的误差。全球CO2通量网的某些站点自行设计了长期观测的设备,主要包括测定室、红外线CO2测定仪及校正设备、数据记录仪等。与涡度相关法相比,箱式法是对森林生态系统CO2通量的间接估计,其优点在于原理简单、仪器价格便宜、操作容易、移动便利、灵敏度高,且能对组成一个生态系统的各个功能团(叶片、根系等)进行定量的测定,这对于阐述微气象学法直接测定的CO2通量出现的现象有帮助,为理解生态系统的功能并进行调控提供了定量数据。国内应用便携式光合测定系统对某个器官进行测定的较多,而长期连续测定很少。应用箱式法,整个森林生态系统的总的通量就是同化器官的CO2同化速率与非同化器官的CO2释放速率之和。
2.3 应用遥感等新技术模型估算法
模型估算法是通过建立准确度相对较高的模型来估算森林生态系统碳储量,因为方法简单、快捷且成本较低,在国际上被广泛的使用到大尺度空间内碳储量的估算上。近年来,各国学者开始逐步在小范围内,针对不同的森林类型和立地条件,创造性的运用各种模型来估算碳蓄积和模拟碳循环过程。
2.3.1 模型模拟法 模型模拟法是通过数学模型估算森林生态系统的生产力和碳储量的方法[37]。模型是研究大尺度森林生态系统碳循环的必要手段,在研究中曾产生过许多模型,其中最著名的是1971年在迈阿密研讨会上提出的Miami模型,以年降水量和年平均气温预测世界不同地带的净第一性生产力(NPP)。此外,影响较大的还有Holdridge生命地带模型、BIOME模型、MAPSS模型、CENTURY,这些模型通常模拟植被的光合、呼吸和土壤分解过程以及不同生态系统层次上碳循环过程之间的关系,通过植被生理生态过程的反映直接模拟气候变化及大气CO2浓度对NPP的影响,预测气候变化对陆地碳循环的影响等。近些年来,利用遥感及相关技术(GIS、GPS等)获得各种植被状态参数,结合地面调查完成植被的空间分类和时间序列分析,可以分析森林生态系统碳的时空分布及动态,并且能够估算大面积森林生态系统的碳蓄积以及土地利用变化对碳蓄积的影响。
模型估计法目前主要分为碳平衡模型、生物生理模型、生物地理模型和生物地球化学模型。碳平衡模型是根据陆地生态系统的分类,模拟各类生态系统的净初级生产力NPP,并从实测数据分析不同生态系统的碳密度,再根据分布面积的估计,然后用简单相乘的方法计算全球陆地的碳平衡。生物生理模型主要根据植物生理特征,如光合作用、CO2施肥效应、植被冠层蒸发、叶片气孔阻力等各种植物生理、植物物理机理,模拟植被生长,分析区域性土地利用的气候影响,探讨植被对气候的反馈等,主要有SIB、BATS和LEAF等几种模型[38]。生物地理模型是描述大尺度上植被或生物群区、生命带、生态系统类型与气候之间关系的模型,可以预测不同环境中各植物类型的优势度。王超等[39]应用BIOME模型模拟了吉林通榆地区NPP通量。这类模型主要描述植被的结构特征,冠层辐射传输、气候土壤和各植物功能类型的生理生态过程,是综合考虑植物生理学特性、优势植物等级、土壤特性和气候的全球植被分类模型。生物地球化学模型是模拟陆地生态系统中碳循环、营养物质循环和水循环等。Tian[40]利用生物地球化学模型研究亚洲季区域碳动态。
总之,模型模拟法适用于估算一个地区在理想条件下的碳储量。这类模型有其独特的优势,它对大气、植被、土壤之间的交换考虑得较全面;其缺陷是没有考虑多个相关因素之间的相互作用和影响,是简单地模拟生态系统中不同平衡状态的碳通量碳储量变化,不能预测植被类型的变化。目前所用的模型多属静态或经验统计模型,其本身包括了许多假设,不确定因素较多且多数仍局限于局部地区甚至个别点,动态或过程模型较少。较为理想的模型是综合考虑多种对象和因素的综合模型。近年来,模型方法也开始由原来的静态统计模型向生态系统机制性模型转变,从单一碳循环模拟转向碳、氮、磷等多种元素循环相耦合的模拟,建立全球碳循环动态模型。碳循环模型与遥感、GIS技术结合,模拟大尺度的碳循环过程是未来模型模拟法的发展趋势。
2.3.2 遥感估算法 遥感估算法主要用于弥补模型模拟的不足,利用遥感手段获得各种植被状态参数,结合地面调查,完成植被的空间分类和时间序列分析,进一步估算土地利用和土地覆被变化对碳储量的影响[41]。它以碳通量与碳循环过程的综合网络观测、生物过程的适应性实验研究以及河流碳输运过程研究为支撑系统的自下而上途径,与以土地利用、土地覆被变化和对地观测数据生态参量反演为基础的自上而下途径,经过相互验证并与尺度转换模型实现有机结合,开展综合观测、调查、比对分析、模拟和评价研究,把握生态系统碳循环的格局与过程规律,辨析自然和人为因素对碳循环过程的影响,以探讨全球气候变化条件下生态系统碳循环过程的演变趋势。近年来,“3S”技术的发展和应用为解决这一问题提供了有效方法,利用遥感手段获得各种植被状态参数,结合地面调查,完成植被的空间分类和时间系列分析,随后可分析森林生态系统碳的时空分布和动态,并估算大面积森林生态系统的碳储量以及土地利用变化对碳储量的影响。
利用遥感估算森林生态碳储量,一般都是通过间接的方式进行,关键是要测量出森林生物量再乘以碳含量转换系数。生物量的遥感估测多利用植被指数(Vegetation Indices)、叶面积指数(LAI)及植被覆盖度等的关系,推断出植被指数与生物量之间的关系进而求生物量。遥感估算森林生物量的方法主要有以下3种:
2.3.2.1 多元回归分析法 一般利用原始波段、植被指数(如归一化差值植被指数、比值植被指数、环境植被指数、叶面积指数等)、地形因子、气象因子等与生物量建立的不同回归模型,得出估测森林植被的回归方程,从而得出生物量。目前,这种方法应用较为普遍。Tomppo等[42]就通过建立多元非线性回归模型,估算出芬兰森林的材积和地上生物量。Ben-challi和Prajapati[43]利用多元回归分析的方法对印度的Haliyal森林的生物量进行了预测。仝慧杰等利用TM影像各波段的灰度值建立回归方程,估算甘肃省小陇山党川林场测量区的生物量[44]。
2.3.2.2 人工神经网络 它是由大量处理单元组成的非线性大规模自适应动力学系统,通过输入林区遥感影像光谱信息及隐层函数转换得出区域森林生物量。Foody等[45]2001年对婆罗洲热带雨林的生物量进行制图分析时,运用神经网络利用TM影像的7个波段进行了回归分析,大大提高了生物量的估算精度。
2.3.2.3 数学建模 它是估算大尺度上森林生态系统生物量的必要手段[46]。利用遥感数据对生物量进行估算不是直接进行的,而是利用遥感数据计算植被指数、叶面积指数、材积等,借助这些因子与生物量的密切关系估算区域生物量。Houghion等[47]结合前人的研究成果,比较了几种不同的数学估算模型,对它们各自的优缺点进行了评估,但始终很难找到一个通用的生物量模型。
近年来,遥感及相关技术和GIS的发展和应用为解决问题提供了有效的方法。利用遥感手段获得各种植被状态参数,结合地面调查,完成植被的空间分类和时间序列分析,随后可分析森林生态系统碳的时间分布及动态,并且能够估算大面积森林生态系统的碳储量以及土地利用变化对碳储量的影响。随着遥感技术的发展,遥感信息在空间分辨率、时间分辨率和光谱分辨率上逐步得到改善,可供选择的遥感信息也越来越多,能满足不同的研究在时间和空间尺度上的要求,因此,遥感技术在森林生态系统及陆地生态系统碳平衡研究中将发挥越来越重要的作用。采用印度遥感卫星数据,以云南省某自然保护区为对象,建立森林蓄积量估测模型,再利用生物量和蓄积量的线性回归方程以及国际上常用的碳转化率,对研究区域阔叶林小班的碳储量进行了估测。
模拟方法采用专门的指示克里格内插技术,并与条件随机模拟相结合,通过确定当前模拟点方差构造中的条件概率分布函数,利用蒙特卡罗法获得当前模拟点位置森林类型的随机函数值[48]。但是,这些研究目前还不能提供基于统计意义和有显著精度的森林碳空间分布[49~51]。运用点同位协同克里格和序列高斯协同模拟方法,研究森林碳(地上部分)储量分布的模拟和尺度转换技术,结合地面调查资料和遥感影像数据探索,获取森林碳分布和进行尺度转换的有效方法,并在森林碳分布的定量评价研究和实践领域中为管理决策者提供一种定量分析方法。
2.4 地面碳同位素方法
该方法假定大气中碳同位素的量是衡定的,植被通过光合作用固定的碳也就含有一定量的碳同位素,由于同位素的衰变,对于土壤或植被中的碳同位素,周转期越长,碳同位素的含量就越低。因此,通过分析植被或土壤中碳同位素的丰度,可以确定植被或土壤中有机碳的年龄,分析有机碳的动态变化。该方法在分析陆地碳平衡时非常有用,是全球变化研究中最为常用的方法之一,特别适用于探讨陆地碳平衡的主导因子及对全球变化的响应等方面[52~57]。
3 小结与讨论
3.1 我国森林固碳研究状况
我国森林生态系统具有复杂的时空结构和内部联系,由于人口的增加、经济发展导致森林资源大幅度减少是森林固碳量减少的主要原因。此后国家大力植树造林,人工林面积的扩大使得森林碳储量又开始回升。研究表明,中国森林对CO2的净吸收(以碳量计算)已经从1990年0.098 Pg/a上升到2000年的0.112Pg/a[58]。王效科等[59~60]以各林龄级森林类型为统计单元,得出中国森林生态系统的植物C储量为3.255 ~ 3.724Pg,且碳密度的分布与人口密度的变化呈负相关,这说明人为干扰极大影响森林植被的固碳作用。
3.2 利用高精度遥感影像数据
遥感技术在森林碳储量的研究中起着重要作用,同时,更高空间分辨率和更多波段的高光谱遥感传感器将使得碳储量的研究更加精确,从源头上解决了数据源对估算精度的影响[61~64]。另外,由于微波遥感具有全天候工作的能力,在碳储量估算中得到了广泛使用。今后,更高波谱和空间分辨率的数据在碳储量估算中的应用将会越来越广泛。
3.3 模型的进一步发展和完善
模型法的发展可有效地解决静态模型估算中不确定性因素对结果的影响,也较好地解决了估算中的尺度转换问题。今后模型法的研究应将更多的方法如神经网络、地统计学、遗传算法等融入森林碳储量估算,建立从几十年到几个世纪或更长时间尺度上的碳循环动态模型,并加强碳循环与 N、P等多种相关元素循环相耦合,这是未来研究的发展方向。
3.4 森林碳储量遥感估算系统的集成
森林生态系统碳循环研究时间持续长,学科涉及面广,时空尺度复杂,数据量庞大,要求有多种新技术和知识作为支撑,强调跨学科的相互交叉、渗透和耦合,并注重人为活动、土地利用和土地覆盖变化对森林生态系统碳循环的影响。今后森林生态系统碳储量的研究将朝着多种方法的综合运用,多学科的介入、联合和渗透,高层次的综合、集成研究方向发展,使得碳储量的估算更加注重多因素的综合影响并能在一定程度上揭示碳储量的动态变化过程及其变化机理[65~71]。高精度遥感影像分析、样地调查和动态模型模拟等方法的综合运用是今后研究森林生态系统碳循环的主要趋势。
综上所述,森林生态系统是一个庞大而复杂的动态变化系统,以上介绍的各种森林固碳方法各有一定的偏重和局限性。在今后的研究中应该注重各种方法的综合运用,取长补短,更要提倡多学科研究人员的参与与联合,使研究向更高、更精确的层次发展。
[1] 李顺龙. 森林碳汇经济问题研究[D]. 哈尔滨:东北林业大学,2005.
[2] 赵敏. 中国主要森林生态系统碳储量和碳收支评估[D]. 北京:中国科学院植物研究所,2004.
[3] FANG J Y,GUO Z D,PIAO S L,et al. Terrestrial vegetation carbon sinks in China, 1981~2000[J]. Sci China D: Earth Sci,2007,50(9):1 341-1 250.
[4] Dixon R K,Solomon A M,Brown S,et al. Carbon pools and flux of global forest ecosystems[J]. Sci,1994,263(5144):185-190.
[5] Brown S L,Schroeder P E. Spatial patterns of aboveground production and mortality of woody biomass for eastern U.S. forests[J]. Ecol App,1999(9):968-980.
[6] Brown S. Management of forests for mitigation of green-house gas emission[M]. Cambridge University Press, Cambridge,1996. 774-797.
[7] 冯宗炜,张家武,邓仕坚. 杉木人工林生物产量的研究[A]. 桃园综合考察报告集[M]. 郑州:河南科学技术出版社,1980. 322-333.
[8] 方精云,刘国华,徐高龄. 中国陆地生态系统碳库[M]. 北京:中国科技出版社,1996. 251-277.
[9] 吕妍,宁虎森,王让会. 人工防护林碳储量估算——以新疆墨玉为例[J]. 环境科学与技术,2010,7(33):28-30.
[10] 张全智,王传宽. 6种温带森林碳密度与碳分配[J]. 中国科学:生命科学,2010,40(7):621-631.
[11] 朱建刚,余新晓,张振明. 森林生态系统碳循环动态仿真系统的设计[J]. 应用生态学报,2009,20(11):2 603-2 609.
[12] 沈文清,马钦彦,刘允芬. 森林生态系统碳收支情况研究进展[J]. 江西农业大学学报,2006,28(2):312-317.
[13] 杨洪晓,吴波,张金屯,等. 森林生态系统的固碳功能和碳储量研究进展[J]. 北京师范大学学报(自然科学版),2005,41(2):172-177.
[14] Woodwell G M,Whittaker R H,Reiners W A,et al. The biota and the world carbon budget[J]. Seience,1978(199):141-146.
[15] Kauppi P E,Mielikainen K,Kuusela K. Biomass and carbon budget of EuroPean forests, 1971 to 1990[J]. Sei,1992,256(5053):70-74.[16] Browns,Lugo A E. Biomass of tropical forest: A new estimate based on forest volumes[J]. Science,1984(223):1 290-1 293.
[17] 冯仲科,罗旭,石丽萍. 森林生物量研究的若干问题及完善途径[J]. 世界林业研究,2005,18(3):25-28.
[18] Whittaker R H,Likens G E. Methods of assessing terrestrial productivity[M]. New york:Springer Verlag,1975. 305-328.
[19] 薛立,杨鹏. 森林生物量研究综述[J]. 福建林学院学报,2004,24(3):283-288.
[20] Birdsey R A,planting a A J,Heath L S. Past and Prospective carbon Storage in United States forests[J]. For Ecol Manag,1993(59):33-40.
[21] Busing R T,Clebseh E C,White P S. Biomass and Production of southern Appalachian cover forests reexamined[J]. Can J For Res,1992(23):760-765.
[22] 方精云. 北半球中高纬度的森林碳库可能远小于目前的估算[J]. 植物生态学报,2004,24(5):635-638.
[23] 赵林,殷鸣放,陈晓非,等. 森林碳汇研究的计量方法及研究现状综述[J]. 西北林学院学报,2008,23(1):59-63.
[24] Leith H,Hittaker R H. Primary Productivity of Biosphere[M]. Berlin: Springer~Verlag,1975.
[25] Brown S,Lugo A E. Aboveground biomass estimates for tropical moist forests of Brazilian Amazon[J]. Interciencia,1992(17):8-18.
[26] 李意德. 海南岛热带山地雨林的森林生物量估测方法的比较分析[J]. 生态学报,1993(13):314-320.
[27] 方精云,刘国华,徐高龄. 我国森林植被的生物量和净生产量[J]. 生态学报,1996(16):497-508.
[28] Brown S,Lugo A E,Wisniewski J. Missing carbon dioxide[J]. Science,1992(11):257.
[29] 樊晓亮,闫平. 森林固碳能力估测方法及其研究进展[J]. 防护林科技,2010(1):61-63.
[30] 王玉辉,周广胜,蒋延龄,等. 基于森林资源清查资料的落叶松林生物量和净生长量估算模式[J]. 植物生态学报,2001,25(4):420-425.
[31] YU Guirui,LI Xuanran,WANG Qiufeng. Carbon Storage and Its Spatial Pattern of Terrestrial Ecosystem in China[J]. J Resour Ecol,2010,1(2):97-109.
[32] 于贵瑞,孙晓敏. 陆地生态系统通量观测的原理与方法[M]. 北京:高等教育教育出版社,2006.
[33] Valentini R,Matteucci G,Dolman A J,et al. Respiration as the main determinant of carbon balance in European forests[J]. Nature,2000(404):861-865.
[34] Malhi Y,Baldoeehi D D,Jarvis P G. The Carbon balance of tropical, Temperature and borcal forests[J]. Plant, Cell Env,1999(22):715-740.
[35] 刘允芬. 中国农业生态系统碳循环研究[J]. 自然资源学报,1995(10):1-8.
[36] 吴家兵,关德新,张弥. 涡动相关法与波文比能量平衡法测算森林蒸散的比较研究——以长白山阔叶红松林为例[J]. 生态学杂志,2005,24(10):1 245-1 249.
[37] 杨洪波,吴波,张金屯,等. 森林生态系统的固碳功能和碳储量研究进展[J]. 北京师范大学学报(自然科学版),2005,41(2):172-177.
[38] Hanqin Tian,Charles A S,Ye Qi. Modeling primary productivity of the terrestrial biosphere in changing environments: Toward a dynamic biosphere mode[J]. Critical Rev Plant Sci,1998,15(5):541-557.
[39] 王超,延晓东,黄耀,等. 应用BIOME-BGC模型研究典型生态系统的潜热通量——半干旱地区吉林通榆的模拟[J]. 气候与环境研究,2006,11(3):404-412.
[40] Tian H Q,Hall C A S,YE Qi. Modeling prinary productivity of the terrestrial biosphere in changing environment: toward a dynamic biosphere model[J]. Critical Rev Plant Sci,1998,17(5):541-557.
[41] 吕景辉,任天忠,闫德仁. 国内森林碳汇研究概述[J]. 内蒙古林业科技,2008,34(2):45-46.
[42] Tomppo E,Nilsson M,Rosengren M,et al. Simultaneous use of Landscap data in estimating large area tree stem volume and aboveground biomass[J]. Remote Sens Environ,2002(82):156-171.
[43] Ben-challi S S,Prajapati R C. Biomass prediction through remote sensing[J]. Indian For,2004(130):599-604.
[44] 仝慧杰,冯仲科,罗旭,等. 森林生物量与遥感信息的相关性[J]. 北京林业大学学报,2007,29(2):156-159.
[45] Foody G M,Cutler M E,Mcmorrow J,et al. Mapping the biomass of Bornean tropical rain forest from remotely sensed data[J]. Glob Ecol Biogeog,2001(10):379-387.
[46] 王秀云,孙玉军. 森林生态系统碳储量估测方法及其研究进展[J]. 世界林业研究,2008,21(5):24-29.
[47] Houghion R A,Lawrence K I,Hackler J L,et al. The spatial distribution of forest biomass in the Brazilian Amazon: A comparison of estimates[J]. Glob Chan Biol,2001(7):731-746.
[48] Feng Y M,Tang S Z,Li Z Y. Simulation of spatial distribution pattern of forest types by using sequential indicator simulation[J]. Acta Ecol Sin,2004,24(5):946-952.
[49] Jiao X M,Xiang W H,Tian D L. Carbon Storage of Forest Vegetation and Its Geographical Distribution in Hunan Province[J]. Journal of Cent South For Univ,2005,25(1):4-8.
[50] Liu P,Ma L Y,Duan J. Study on computerized forest dynamic models[J]. World For Res,2007,20(3):45-50.
[51] XU Xinliang,LI Kerang. Biomass Carbon Sequestration by Planted Forests in China[J]. Chin Geog Sci,2010,20(4):289-297.
[52] Del Galdo I,Six J,Peressotti A,et al. Assessing the impact of land use change on soil C sequestration in agricultural soils by means of organic matter fractionation and stable C isotopes[J]. Glob Chan Biol,2003(9):1 204-1 213.
[53] Hagedorn F,Spinnler D,Bundt M,et al. The input andfate of new C in two forest soils under elevated CO2[J]. Glob Chan Biol,2003(9):862-872.
[54] Matamala R,Gonzalez-Meler MA,Jastrow J D,et al. Impacts of fine root turn over on forest NPP and soil C sequestration potential[J]. Science,2003,302(5649):1 385-1 387.
[55] Reilly C M,Alin S R,Plisnier P D,et al. Climate change decreases aquatic ecosystem productivity of Lake Tanganyika, Africa[J]. Nature,2003(424):766-768.
[56] Grünzweig J,Sparrow S D,Yakir D. Impact of agricultural land use change on carbon storage in boreal Alaska[J]. Glob Chan Biol,2004(10):452-472.
[57] Marie-Anne,Johan S,David H,et al. Decomposition of soil and plant carbon from pasture systems after 9 years of exposure to elevated CO2:impact on C cycling and modeling[J]. Glob Chan Biol, 2004,10(11):1 365-2 486.
[58] Streets D G,Jiang K J,HU XL,et al. Recent reductions in China’s greenhouse gas emissions[J]. Science,2001(294):1 835-1 836.
[59] Wang X K,Feng A W,Ou Yang Z Y. Vegetation carbon storage and density of forest ecosystems in China[J]. Chinese J App Ecol,2001,12(1):13-16.
[60] Wang X K,Feng Z W. The potential to sequester atmospheric carbon through forest ecosystems in China[J]. Chinese J Ecol,2000,19(4):72-74.
[61] 张茂震,王广兴,周国模,等. 基于森林资源清查、卫星影像数据与随机协同模拟尺度转换方法的森林碳制图[J]. 生态学报,2009,29(6):2 919-2 928.
[62] 曹吉鑫,田赟,王小平,等. 森林碳汇的估算方法及其发展趋势[J]. 生态环境学报,2009,18(5):2 001-2 005.
[63] 张骏,袁位高,葛滢. 浙江省生态公益林碳储量和固碳现状及潜力[J]. 生态学报,2010,30(14):3 839-3 848.
[64] 林清山,洪伟. 中国森林碳储量研究综述[J]. 中国农学通报,2009,25(6):220-224.
[65] 王兵,王燕,郭浩,等. 江西大岗山毛竹林碳贮量及其分配特征[J]. 北京林业大学学报,2009,31(6):39-42.
[66] 包艳丽,牛树奎,张国林. 天山云杉林碳储量研究[J]. 干旱区资源与环境,2009,23(9):113-117.
[67] 殷鸣放,赵林,陈晓非. 长白落叶松与日本落叶松的碳储量成熟龄[J]. 应用生态学报,2008,19(12):2 567-2 571.
[68] 耿相国. 平原区农田防护林杨树单株碳含量、碳储量及其分配[D]. 北京:北京林业大学,2010.
[69] 应天玉,李明泽,范文义. 哈尔滨城市森林碳储量的估算[J]. 东北林业大学学报,2009,(9):33-35.
[70] 闫德仁,闫婷. 内蒙古森林碳储量估算及其变化特征[J]. 林业资源管理,2010,3(6):32-33.
[71] 叶金盛,佘光辉. 广东省森林植被碳储量动态研究[J]. 南京林业大学学报(自然科学版),2010,34(4):7-12.
