猪肚菇子实体发育过程中抗氧化酶活性的变化
2010-03-24董洪新程显好蔡德华
董洪新,程显好,朱 林,蔡德华,李 玉*
(1.吉林农业大学菌物研究所,吉林 长春 130118;2.鲁东大学生命科学学院,山东 烟台 264025)
猪肚菇子实体发育过程中抗氧化酶活性的变化
董洪新1,2,程显好2,朱 林2,蔡德华2,李 玉1,*
(1.吉林农业大学菌物研究所,吉林 长春 130118;2.鲁东大学生命科学学院,山东 烟台 264025)
研究猪肚菇子实体不同发育时期膜脂过氧化产物丙二醛(MDA)含量、超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性变化情况。结果表明:随着猪肚菇子实体生长和衰老,子实体中MDA逐渐积累,膜脂过氧化程度加重;SOD和CAT活性先升高后降低,SOD、CAT活性最高值分别出现在杯形期和钉头期,而POD活性则基本稳定。因此,活性氧代谢失调引发的生物膜结构破坏是猪肚菇子实体衰老的重要原因之一。关键词:猪肚菇;子实体;发育;抗氧化酶
自由基学说认为,生物衰老过程是活性氧大量积累与代谢失调的过程[1]。近来研究发现活性氧可能作为第二信使参与生物体内的代谢及生命活动[2]。生物在正常或非正常代谢进程中可通过多种途径产生活性氧自由基,同时它们又存在着清除这些自由基的机制, 包括保护酶系统,如过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)等和非保护酶系统,如VC、类胡萝卜素、VE等。生物种类的不同,表现出活性氧代谢各因素间相互作用的复杂性和多样性。近来关于抗氧化酶活性的相关研究报道多集中在小麦、果蔬、花卉等植物上[3-5],而对衰老过程中食用菌子实体抗氧化酶的响应及调节机理则未见报道。
猪肚菇(Panus giganteus)又名大杯香菇、大革耳等,是国内近年来新开发的一种珍稀食用菌,其口感风味独特,有猪肚般的滑腻,因此而得其商品名[6-7]。猪肚菇子实体的菌肉肥厚,质地细腻,脆嫩可口,汤汁鲜美营养丰富,其生长条件粗犷,适用原料广泛,且在夏季出菇,对于解决食用菌生产淡季和调节市场供应很有意义,具有广阔的开发应用前景。猪肚菇子实体发育和衰老过程中,在形态上有典型的4个时期,可以代表其发育和衰老的不同阶段,是研究子实体发育和衰老的好材料。大型真菌子实体的形成和衰老是十分复杂的生理生化过程,研究子实体形成和衰老机理,对于调节大型真菌子实体发育,具有重要理论和实践意义。本研究以猪肚菇为研究材料,探究子实体形成和衰老过程中其细胞抗氧化酶活性和脂质过氧化情况,为揭示其抗衰老机理提供理论依据,同时对理解子实体生理代谢规律具一定意义。
1 材料与方法
1.1 材料、试剂与仪器
供试菌株猪肚菇 LD-1 鲁东大学食用菌工程技术研究中心。
葡萄糖、KH2PO4等试剂均为国产分析纯;氮蓝四唑(NBT)为进口分装。
752N型紫外-可见分光光度计、冷冻离心机。
1.2 方法
1.2.1 子实体培养
将培养料按照配方(棉籽壳83%,麸皮15%,石灰2%,含水量63%)拌料、装袋,灭菌、接种和培养,菌丝满袋后移入栽培室采用袋内覆土出菇,覆土厚度3~4cm。栽培室温度在23~32℃,空气相对湿度 90%~92%。待子实体形成后,分别采集不同发育阶段的子实体样品,采摘后的子实体放于冰盒中,立即带回实验室,用于生理指标测定。
1.2.2 酶活性及MDA 含量的测定
超氧化物歧化酶(SOD)的活性以(NBT)光化还原法测定,以A560nm下抑制NBT还原的50%所需的酶量为SOD的一个酶活单位(U),用U/g表示;过氧化氢酶(CAT)活性的测定采用紫外吸收法,过氧化物酶(POD)活性的测定采用愈创木酚法,在一级反应范围内,分别以每分钟内A240和A470变化0.01为 CAT和POD的一个酶活单位(U),用U/(mg·min)表示。
MDA含量测定:硫代巴比妥酸(TBA)比色法测定[8]。
1.2.3 数据分析
采用Microsoft Excel 2003和 SPSS13.0统计分析软件进行数据分析及差异显著性检验,所有数值均为3次重复平均值。
2 结果与分析
2.1 猪肚菇子实体不同发育阶段形态的变化
猪肚菇子实体发育的各个阶段形态差别较大,大致划分为如图1所示4个时期。图1a为棒形期,此期为子实体发育的初级阶段,即原基形成期,原基初期白色,球形或似卵形,后长成倒棒形。出土部分见光变灰色,逐步加深至鼠灰色或黑褐色。表面有灰白色鳞片。图1b为钉头期,此期为原基分化,菌盖、菌柄、菌褶等器官形成期,棒形原基顶端细胞迅速增殖长成钉头状,纵切可见菌盖、菌褶和硕长的菌柄。内菌幕未破裂或刚破裂。图1c为杯形期,此期菌盖迅速发育,中部菌肉增厚,内菌幕破裂,露出片状菌褶。盖顶平至下凹,颜色由盖沿开始逐渐变浅。图1d为成熟期,菌盖完全伸展,菌肉变薄,颜色变浅,盖缘平直至波曲,释放出担孢子。

图1 猪肚菇子实体发育的4个时期Fig.1 Four stages of fruit body development for Panus giganteus
2.2 猪肚菇子实体不同发育阶段SOD活性变化
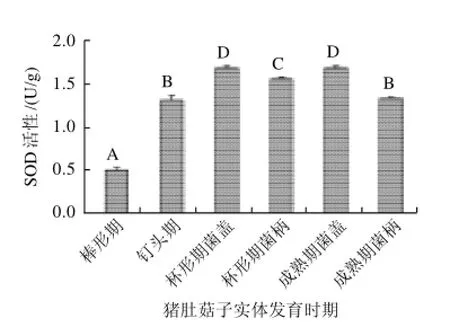
图2 猪肚菇子实体不同发育时期的SOD活性变化Fig.2 Change in SOD activity during fruit body development of Panus giganteus
如图 2所示,猪肚菇子实体发育过程中,各个时期SOD活性达到显著差异(P<0.05),其子实体中SOD活性呈现先升高后降低的趋势,即由棒形期到杯形期发育期间,子实体中SOD逐渐增加,在杯形期 SOD活性达到最大,杯形期菌盖 SOD活性是棒形期的 3.5倍,杯形期菌柄 SOD活性是棒状期的 3.2倍。随后 SOD活性开始降低,成熟期菌柄SOD活性明显低于杯形期。
在杯形期和成熟期,菌盖和菌柄的 SOD含量都达到差异显著(P<0.05),菌盖中 SOD含量显著高于菌柄。菌盖的生理代谢活动比菌柄更活跃,在研究大型真菌对于金属离子的吸收和富集过程中也发现,富集的金属离子在子实层中含量最高,其次是菌盖部分,而菌柄中含量最低[9]。
SOD广泛分布于细胞各组成部分,是防御自由基危害的第一道防线,催化O2-·或 HO·歧化为H2O2,一定程度上控制着活性氧(ROS)的含量,但其活性又受ROS的影响。对环境变化比较敏感。在猪肚菇子实体发育过程中,也表现出不同阶段的明显差异。
2.3 猪肚菇子实体不同发育阶段POD 活性的变化
过氧化物酶(POD)是以过氧化氢为电子受体催化底物氧化的酶。主要存在于细胞的过氧化物酶体中,以铁卟啉为辅基,可催化过氧化氢、氧化酚类和胺类化合物,具有消除过氧化氢和酚类、胺类毒性的双重作用。如图 3 所示,在猪肚菇子实体生长和衰老过程中,与SOD活性的变化趋势不同,POD活性基本保持稳定。
植物衰老过程中,其POD变化趋势随植物种类不同而不同。如荔枝、猕猴桃、胡柚等果实随贮藏时间延长,POD活性增强,认为POD活性升高可以作为果实衰老的指标;而杏果实成熟衰老过程中,POD活性则呈下降趋势[10]。猪肚菇子实体生长和衰老过程中,无论菌柄还是菌盖,其POD活性都基本保持稳定,所以不宜作为子实体衰老的生理指标。POD活性在子实体发育过程中保持基本稳定属于食用菌的普遍现象还是个例现象,需要进一步研究和探讨。
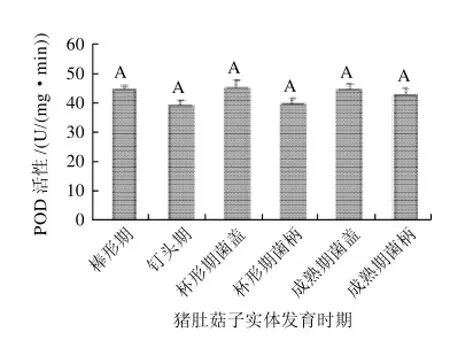
图3 猪肚菇子实体不同发育时期的POD活性变化Fig. 3 Change in POD activity during fruit body development of Panus giganteus
2.4 猪肚菇子实体不同发育阶段CAT 活性的变化
CAT主要存在于过氧化物体,线粒体和乙醛酸循环体中,CAT是细胞内H2O2的重要清除剂,但CAT清除H2O2的效率非常低,CAT活性显著下降会造成H2O2积累。胞内H2O2不能及时清除,可与过量的O2·生成氧化性更强的·OH。
从图 4可以看出,CAT活性从棒形期到钉头期迅速升高,然后逐渐降低。与SOD活性变化相比CAT活性高峰期的出现较早,在钉头期达到最高。在杯形期和成熟期,菌盖和菌柄之间的CAT酶活都达到差异显著(P<0.05),在杯形期的CAT活性菌盖高于菌柄,而在成熟期的CAT活性菌盖低于菌柄。

图4 猪肚菇子实体不同发育时期的CAT活性变化Fig.4 Change in CAT activity during fruit body development of Panus giganteus
2.5 猪肚菇子实体不同发育阶段MDA含量的变化
膜的稳定性是生物衰老过程中的重要生理指标。膜脂过氧化加剧会导致膜脂不饱和度降低,膜流动性下降,对为膜所束缚的细胞酸性磷酸脂酶起明显的增溶作用,膜脂分子构象发生改变,致使细胞膜透性增强[11]。丙二醛(MDA)是细胞膜脂过氧化作用的主要产物之一,MDA在生物体内的积累是活性氧毒害的表现,其含量是判断膜脂过氧化程度的一个重要指标。
由图 5可知,随着猪肚菇子实体的生长和衰老,子实体中MDA含量逐渐增加,到成熟期时,MDA含量明显增加,其含量为棒形期的6.84倍,表明细胞膜膜脂过氧化程度加剧。所以MDA含量明显增加,可以作为猪肚菇子实体衰老的主要指标。
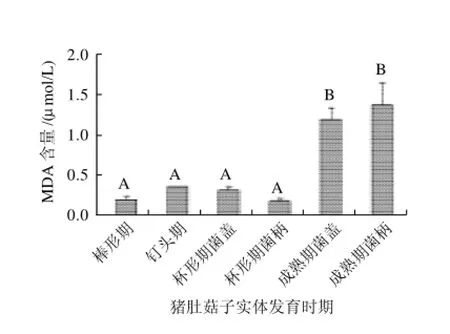
图5 猪肚菇子实体不同发育时期的MDAs含量变化Fig.5 Change in MDA content during fruit body development of Panus giganteus
3 讨 论
在正常情况下植物体内有一套完整的活性氧清除系统(抗氧化酶和抗氧化剂)能将活性氧的产生和清除维持在动态平衡状态。SOD是植物氧代谢过程中的一个极为重要的诱导酶,它能催化超氧阴离子自由基歧化反应成为基态的分子氧和H2O2,并与 POD和 CAT组成活性氧防御酶系统[12-13]。抗氧化酶活性变化与子实体衰老密切相关。在衰老过程中活性氧大量产生,生物体可通过抗氧化保护酶系统中的 SOD、POD、CAT和 APX来调节活性氧产生与清除之间的平衡。
该实验中,随着子实体生长和衰老的进行,子实体中SOD活性开始增加,这可能是生物抗氧化衰老的一种防卫响应,随着活性氧物质的增加,防御系统进一步启动,子实体中CAT活性进一步升高,起到协助清除活性氧物质的作用。但是,随着衰老的不可逆转,整个防御系统遭到破坏,SOD、CAT的活性随之降低,MDA含量迅速升高。但猪肚菇子实体生长和衰老进程中,POD活性则基本保持稳定。因此,活性氧代谢失调引发的生物膜结构破坏是猪肚菇子实体衰老的重要原因之一。
自由基引起的衰老理论在近半个多世纪以来得到了人们的广泛关注,很多研究证实衰老过程中CAT、SOD、POD等抗氧化酶的活性均降低,导致自由基代谢产物增高,致使线粒体结构的损伤,氧化呼吸传递链传递受阻,使自由基的生成量进一步增加,从而加速了衰老进程[14]。Orr等[15]将铜(Cu)锌(Zn)超氧化物歧化酶和过氧化氢酶基因(sod与cat)导入果蝇,使其平均寿命和最高寿限都不同程度得到了延长,与之相关的与蛋白质的氧化损伤现象减轻。从本实验得知,在猪肚菇子实体生长阶段,CAT和SOD活性维持在较高水平,活性氧自由基的水平亦较低;但在衰老阶段,CAT和SOD活性明显下降,致使自由基水平提高,并导致细胞膜损伤加剧。另外,CAT和SOD的活性随猪肚菇子实体的发育阶段的不同呈现明显变化的特点,表明除MDA水平之外,CAT和SOD的活性也可以作为猪肚菇子实体的衰老标记。因此,如果能利用合适的技术手段从生理水平上提高CAT和SOD的活性,将能延缓其衰老过程,延长猪肚菇的货架期,这需要今后进一步深入研究。
[1]关军锋, 束怀瑞. 苹果果实衰老与活性氧代谢的关系[J]. 园艺学报, 1996, 23(4): 326-328.
[2]RON M. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002,7(9) : 405-410.
[3]赵天宏, 孙加伟, 付宇, 等. CO2和O3浓度升高对春小麦活性氧代谢及抗氧化酶活性的影响[J]. 中国农业科学, 2009, 42(1): 64-71.
[4]任艳芳, 何俊瑜, 王思梦, 等. 脐橙花开放和衰老过程中抗氧化酶活性变化[J]. 中国农学通报, 2009, 25(19): 62-64.
[5]房伟民, 郭维明, 陈俊愉. 嫁接提高菊花耐高温与抗氧化能力的研究[J]. 园艺学报, 2009, 36(9): 1327-1332.
[6]邓旺秋, 李泰辉, 陈枝南, 等. 栽培食用菌猪肚菇的学名考证[J]. 食用菌学报, 2006, 13(3): 71-74.
[7]戴玉成, 周丽伟, 杨祝良, 等. 中国食用菌名录[J]. 菌物学报, 2010, 29(2): 1-21.
[8]李玲. 植物生理学模块实验指导[M]. 北京: 科学出版社, 2009: 1-173.
[9]PAVEL K, LUBOMIR S. A review of trace element concentrations in edible mushrooms[J]. Food Chemistry, 2000, 69: 273-281.
[10]吴彩娥, 寇晓红, 王文生. 果实成熟衰老与保护酶系统的关系[J]. 中国果菜, 2002(6): 23-24.
[11]陈拓, 安黎哲, 冯虎元, 等. UV-B辐射对蚕豆膜脂过氧化的影响及其机制[J]. 生态学报, 2001, 21(4): 579-583.
[12]龙云, 邓美玲, 谈锋. 绞股蓝对水分胁迫的适应性研究[J]. 西南师范大学学报: 自然科学版, 1999, 24(1): 81-86.
[13]NEILL S J, DESIKAN R, CLARKE A, et al. Hydrogen peroxide and nitric oxide as signaling molecules in plants[J]. Journal of Experimental Botany, 2002, 53: 1237-1247.
[14]MELOV S, RAVENSCROFT J, MALIK S, et al. Extension of life-span with superoxide dismutase/catalase mimetics[J]. Science, 2000, 289: 1567-1569.
[15]ORR W C, SOHAL R S. Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster[J]. Science, 1994, 263: 1128-1130.
Change of Antioxidant Enzyme Activities in Fruit Body Development Process of Panus giganteus
DONG Hong-xin1,2,CHENG Xian-hao2,ZHU Lin2,CAI De-hua2,LI Yu1,*
(1. Institute of Mycology, Jilin Agricultural University, Changchun 130118, China;2. College of Life Science, Ludong University, Yantai 264025, China)
In this stady, the change in the content of malonaldehyde (MDA) and the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) in fruit body of Panus giganteus during its development process were investigated. Results indicated that MDA content exhibited a gradual accumulation and the membrane lipid peroxidation was increased; meanwhile, the activities of SOD and CAT exhibited an initial increase and a final decrease trend; the highest activity of SOD and CAT was observed in the cup stage and pin-head stage. However, POD activity was stable during the stage of growth and senescence. Therefore, the metabolic disorder induced by the accumulation of active oxygen species was one of important factors for the senescence of Panus giganteus.
Panus giganteus;fruiting body;development;antioxidant enzymes
S646.9
A
1002-6630(2010)24-0469-04
2010-03-24
“十一五”国家科技支撑计划项目(2008BADA1B04);山东省农业良种工程项目(2008LZ-008)
董洪新(1976—),男,讲师,博士研究生,研究方向为食用菌栽培。E-mail:ytdhx@163.com
*通信作者:李玉(1944—),男,教授,博士,研究方向为菌物学。E-mail:yuli966@126.com
