贵州四个民族人群线粒体DNA编码区的多态性
2010-02-16李彬彬钟复光易红生王先然李良芳王丽兰吴立甫
李彬彬,钟复光,易红生,王先然,李良芳,王丽兰,吴立甫
(1.广东医学院病理生理学教研室,东莞 523808;2.贵阳医学院医学科学研究所,贵阳 550004;3.贵州省天柱县人民医院,天柱 556700;4.贵州省赫章县人民医院,赫章 553200;5.贵州省务川县人民医院,务川 564300)
人类线粒体NA(mitochondrial DNA,mtDNA)是存在于细胞核外的唯一遗传物质,为闭环双链结构,由16569对碱基组成,包括非编码区和编码区(编码37个基因)。线粒体DNA由于严格的母系遗传、进化速率快、群体内变异大、分子结构简单、拷贝数多等特性,已成为群体遗传学、考古学以及法医学等领域非常有用的遗传标记。
目前对mtDNA多态性的研究主要集中在非编码区中的两个高突变区(hypervariable segmentⅠandⅡ,HVSⅠ、Ⅱ),但由于这两个区域不能提供足够的多态信息量,识别能力有限,因此,对mtDNA编码区进行单核苷酸多态性(single nucleotide polymorphis m,SNP)研究,将为进一步建立mtDNASNP数据库以及线粒体SNP基因座在考古学、进化学和法医学个人识别等应用奠定基础[1,2]。本文采用聚合酶链反应-限制性片段长度多态性(PCRRFLP)技术及DNA测序法对贵州土家族、侗族、仡佬族和彝族共145例样本mtDNA编码区9个多态位点进行了研究,为各少数民族群体遗传学研究提供新的数据。
1 材料和方法
1.1 样本采集及mtDNA提取
随机选取145例健康妇女所生胎儿的胎盘标本,其中土家族38例,侗族37例,仡佬族40例,彝族30例,分别采自贵州省天柱、道真、沿河、赫章等县市的民族聚居地。各样本家庭内部3代均为同一民族,彼此间无直接血缘关系。按王文等[3]的碱变性法从胎盘组织中提取mtDNA,0.8%琼脂糖凝胶电泳检测目的条带。
1.2 mtDNA编码区的PCR扩增
依据文献报道[4,5],选取mtDNA编码区在亚洲人群中多态性较高的8个SNP(A10398G、C10400T、A663G、A4833G、C5178A、G7598A、G12406A、A13263G)基因座及COⅡ/tRNAlys基因间9bp缺失,对相应片段进行PCR扩增。反应参数为94℃预变性5min,94℃变性30s、Tm退火40s、72℃延伸40s,共30个循环,最后72℃延伸5min。扩增产物经2%琼脂糖凝胶电泳和8%聚丙烯酰胺凝胶电泳进行检测。引物序列、退火温度及片段长度见表1。
1.3 扩增产物的RFLP检测
将PCR扩增产物参照表1进行相应的限制性酶切反应。所用限制性内切酶DdeⅠ、HaeⅢ、AluⅠ、HhaⅠ、HpaⅠ均购自北京华美生物公司。酶切反应体系为:PCR产物4μL,限制性内切酶2~4U,10×buffer 1μL,加灭菌水至10μL,37℃温育过夜。酶切产物用3%琼脂糖凝胶电泳进行分离检测。
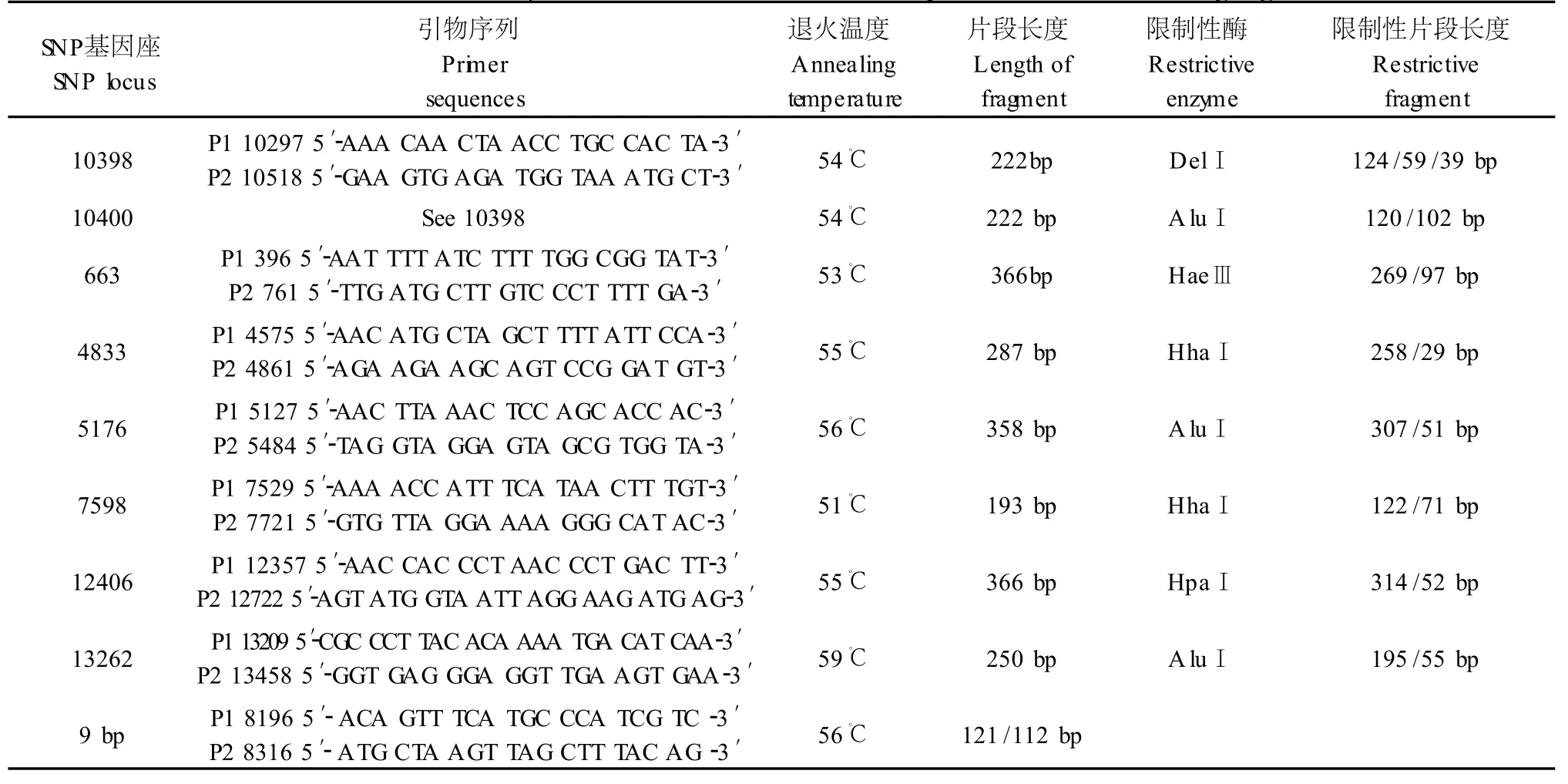
表1 mtDNA编码区的PCR反应参数及RFLP分析Tab.1 The reaction parameters of PCR and RFLP analyses in the mtDNA coding region
1.4 扩增产物的测序
选取部分样本的PCR扩增产物经2%琼脂糖电泳后用小量胶回收试剂盒(大连宝生物工程有限公司)进行回收纯化,采用BigDyeT M Teminator Cycle Sequencing Ready试剂盒(美国PE公司)进行序列测定。测序结果与mtDNA修订剑桥标准序列(revised cambridge reference sequence,rCRS)[6]进行比对。
1.5 数据分析
根据PCR-RFLP及测序结果,计算贵州4个民族群体的mtDNA 9bp缺失频率及8个SNP基因座的等位基因频率。各位点碱基完全相同者视为同一单倍型,统计各单倍型分布频率。
2 结果
2.1 PCR-RFLP及测序结果
对贵州4个民族人群145例样本进行扩增结果表明,mtDNA 9bp缺失频率分别为土家族18.4%(7/38),侗族29.7%(11/37),仡佬族25%(10/40),彝族16.7%(5/30),平均缺失频率为22.8%。其他编码区RFLP分析共发现有187个扩增片段存在特异突变位点,进一步测序分析确定其突变类型(表2)。
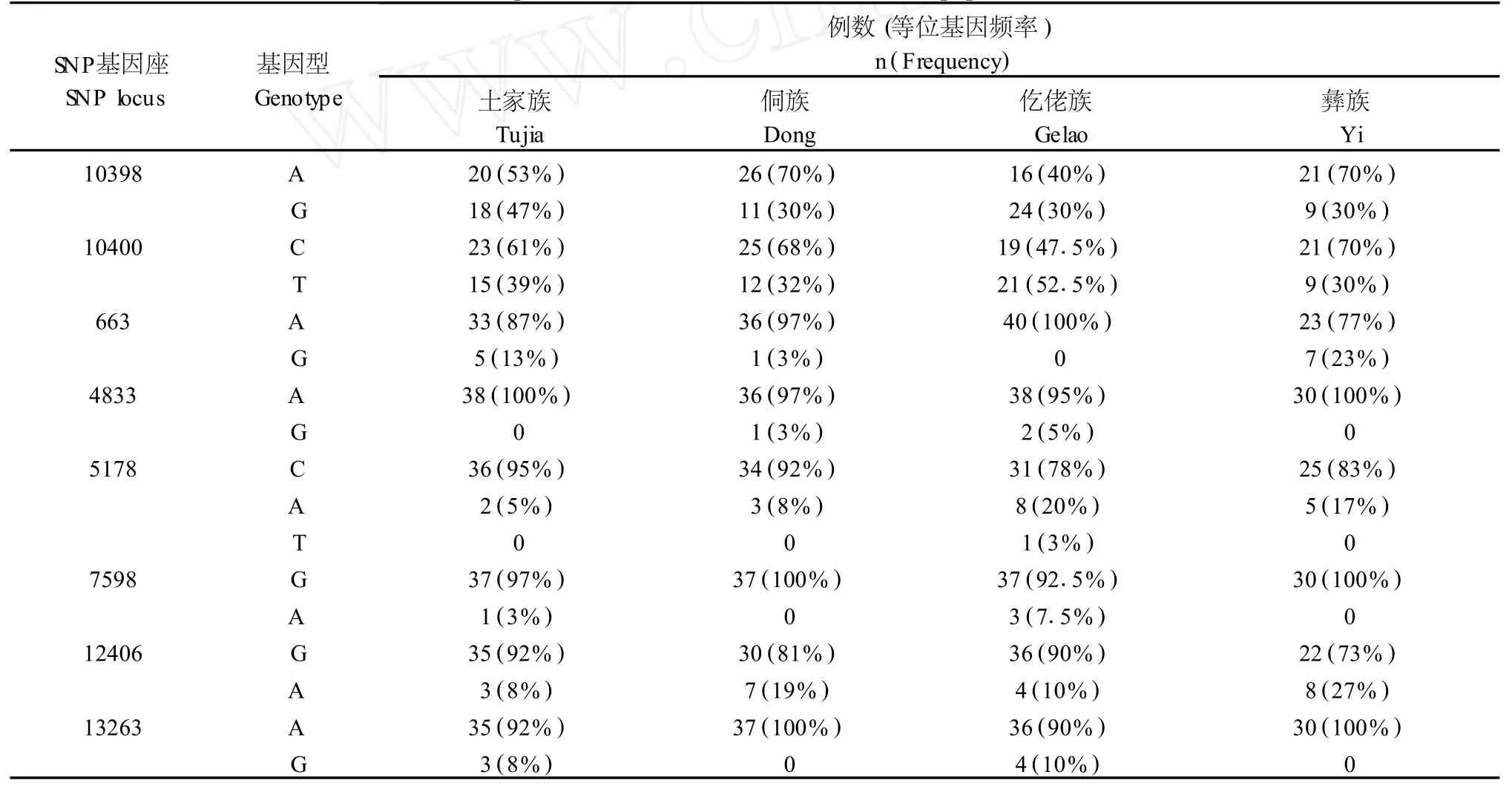
表2 贵州4个群体8个mtDNA SNP基因座的等位基因频率Tab.2 Allele frequencies of 8mtDNA SNP loci in four populations from Guizhou
2.2 mtDNA编码区的多态性分布
贵州4个民族群体8个mtDNA SNP基因座的等位基因频率分布见表2。在145例样本中,共检出14种单倍型,单倍型及其频率分布见表3。

表3 9个mtDNA多态位点在贵州4个民族群体的单倍型分布Tab.3 Haplotype distribution of 9mtDNA polymorphism site in four populations from Guizhou

注:*:“1”表示存在mtDNA 9bp缺失;“2”表示没有9bp缺失。Note:*:“1”denotes the presence of the 9bp deletion,“2”denotes nondeletion
3 讨论
近年来研究发现,仅通过mtDNA高变区对人群进行世系发育分析,得出的结论往往不能完整、真实地反映系统关系,而结合编码区的一些特征位点,则能更加准确、有效地界定群体的mtDNA世系发育关系[4,5,7]。在法医学鉴定中,由于mtDNA为单倍型遗传[8],仅仅依靠mtDNA高变区内的SNP,有时很难将两个无关个体完全区分开,常需要增加编码区的检测位点才能提高mtDNA的个体识别能力[2,9]。同时,编码区信息也将为某些线粒体相关遗传病的研究提供基本数据资料。
有研究表明,人类mtDNA COⅡ/tRNAlys基因间有2个串联重复的9bp(CCCCCTCTA)序列,此重复序列中一个重复单位的缺失,在亚洲人以及祖先是亚洲人的群体中很普遍[10],也是检测人类群体多态性的重要指标。不同人种群、不同地域人群mtDNA 9bp缺失频率不同[11,12]。本次研究的贵州4个民族群体的9bp缺失频率范围为16.7%~29.7%,平均缺失频率为22.2%,与已报道的[12]我国西南地区苗族(32.4%)、傣族(23.5%)和白族(16.7%)等群体较接近,但明显高于北方的蒙古族(4.0%)、维吾尔族(3.3%)和哈萨克族(0.0%)等,这与目前认为的南方民族人群的缺失频率要高于北方民族人群是相一致的[12]。
相对于非编码区的高突变率,mtDNA编码区的序列比较稳定,碱基突变较少。依据文献报道[4,5],我们选取了在亚洲人群中具有较高多态性的8个mtDNA编码区SNP基因座对贵州4个少数民族群体进行了初步研究。在所检测的基因座中,10398 A→G、10400 C→T突变在4个群体中较普遍,是两种古老的突变类型,在已报道[13]的国内各少数民族群体中均有较高的分布频率。此外,土家族人群663G的多态性频率为13%,侗族人群12406A的多态性频率为13%,仡佬族人群5178A的多态性频率为20%,彝族人群663G、12406A的多态性频率为23%、27%,与相应位点其他民族的分布频率存在较大差别。这些结果提示在贵州4个民族群体中可能存在不同的mtDNA编码区突变热点。
本次研究的4个民族均来自贵州边远山区,山岭阻隔,地形相对封闭,各民族之间隔离程度较高,民族特异性保存较好。在145例样本9个多态位点中共检测出14种单倍型,其中H1、H13、H12单倍型占有较高的比例,分别为19.3%、17.2%和15.2%,占总分析样本的51.7%。从单倍型的分布来看,仡佬族的单倍型类型最多,有11种,土家族有10种,侗族有8种,彝族最少,只有6种,不同民族在单倍型分布频率上存在一定差异。
[1]高路,董永利,郝肇菁,等.云南16个少数民族群体的线粒体DNA多态性研究[J].遗传学报,2005,32(2):118-123.
[2]刘雅诚,郝金萍,严江伟,等.线粒体DNA编码区的多态性[J].法医学杂志,2006,22(21):45-47.
[3]王文,施立明.一种改进的动物线粒体DNA提取方法[J].动物学研究,1993,14:197-198.
[4]Kivisild T,Tolk HV,Parik J,et al.The emerging limbs and twigs of the eastAsian mtDNA tree[J].MolBiol Evol,2002,19(10):1737-1751.
[5]Yao YG,Kong QP,Bandelt HJ,et al.Phylogeographic differentiation of mitochondrial DNA in Han Chinese[J].Am J Hum Genet,2002,70(3):635-651.
[6]Andrew RM,Kubacka I,Chinnery PF,et al.Reanalysis and revision of the Cambridge reference sequence forhuman mitochondrialDNA[J].Nat Genet,1999,23:147.
[7]TorroniA,RichardsM,MacaulayV,et al.mtDNA haplogroups and frequencypatterns in Europe[J].Am J Hum Genet,2000,66(3):1173-1177.
[8]Giles RE,Blane H,Cann HM,et al.Maternal inheritance of human mitochondrialDNA[J].Proc NatlAcad SciUSA,1980,77:6715-6719.
[9]顾明波,柳杰,杜庆新,等.中国汉族人群的线粒体DNA控制区多态性研究[J].中国法医学杂志,2001,16(1):6-9.
[10]BarrientonsA,Casademont I,SolansA,et al.The 9 bp deletion in region Ⅴ of mitochondrial DNA:evidence of mutation recurrence[J].Hum Genet,1995,96:225-228.
[11]Yao YG,Kong QP,Wang CY,et al.Different matrilineal contributions to genetic structure of ethnic groups in the silk road region in China[J].MolBiol Evol,2004,21:2265-2280.
[12]姚永刚,袁志刚,周曾娣,等.中国民族人群线粒体DNA 9bp序列缺失的分布[J].自然科学进展,2001,11(4):353-359.
[13]周海燕,倪斌,邹永华,等.应用mAPLP方法分析湖南汉族、苗族和土家族mtDNA编码区多态性[J].2008,30(6):716-722.
