不同诱变处理对苦荬菜M1代农艺性状和品质影响的初报
2010-01-02唐凤兰刘丽陈积山张月学韩微波刘杰淋刘风岐
唐凤兰,刘丽,陈积山,张月学,韩微波,刘杰淋,刘风岐
(1.黑龙江省农业科学院草业研究所,黑龙江哈尔滨 150086;2.哈尔滨师范大学,黑龙江 哈尔滨150086)
研究表明,空间飞行(AF)和60Co-γ射线辐照植物干种子均会发生明显的变异,如生长势变化、同工酶谱带增减、染色体数目改变、叶绿体损伤[1-3]等。当前辐射诱变已成为世界上普遍应用的育种方法之一,获得了许多宝贵的突变材料和优良品系[4,5]。目前对突变后代的研究多集中在水稻(Oryza sativa)、蔬菜等作物上[6-8],在牧草方面的研究相对较少。
苦荬菜(Lactuca indica)是菊科一年或越年生草本植物。它的高产、优质、适应性强、适口性好,适宜北方地区种植,是极具推广价值的优良牧草[9]。因此,在经过多年的驯化和选育苦荬菜的基础上,旨在为南方和华北、东北地区大面积种植提供各种畜禽的优良多汁饲料[10],本研究利用空间飞行和60Co-γ射线诱变育种技术处理苦荬菜风干种子,通过对当代突变体的主要农艺性状、品质构成因子的分析,进一步探讨辐照突变群体的遗传变异规律,为辐射诱变育种进一步应用于生产提供科学依据。
1 材料与方法
1.1 供试材料
选用黑龙江省农业科学院草业研究所提供的龙饲0374苦荬菜干种子。
1.2 试验方法
2003年11月3日通过第18颗返回式卫星搭载苦荬菜干种子在酒泉发射基地升空,在空间运行18 d后返地,用AF(空间飞行)表示该处理的所有种子及材料。同时在黑龙江省农业科学院玉米研究所,将相同等量的苦荬菜干种子利用60Co-γ钴源进行辐照,分别为10,20,30 Gy的3个剂量水平。将未做任何处理的相同等量的苦荬菜干种子作为对照(CK)。

粗蛋白(CP):采用凯氏定氮法(Kjeldahl method,GB/T6432-94)测定。粗脂肪(CF):采用索氏(Soxhlet)脂肪法测定。粗纤维(CDF):采用浓H2SO4消煮法(GB/T6434-94),根据Van Soest和 Roberston方法测定[12]。株高:每个小区随机取10株单株测定其生长高度,取平均值。
1.3 统计方法
本试验数据应用Excel(Office 2000)和SPSS(版本11.5)软件进行统计分析,5%水平下Student-Newman-Keuls(S-N-K)多重比较检验均值之间差异显著性。
2 结果与分析
2.1 发芽率与发芽势的变化
发芽率是指一定数量的干种子在适宜条件下,在规定天数内,发芽的种子数占供试种子数的百分率。发芽势表示种子发芽的快慢和整齐度,它们是测定种子萌发活力的重要指标[12]。空间飞行和60Co-γ射线3种剂量(10,20,30 Gy)处理下的发芽势和发芽率差异显著(P<0.05),空间飞行和60Co-γ射线处理均能提高苦荬菜M1代的发芽势和发芽率,其中AF和20 Gy提高的幅度较对照最大(表1)。
2.2 株高的变化
空间飞行和60Co-γ射线3种剂量(10,20,30 Gy)处理的株高差异显著(P<0.05),尽管处理的总体呈负增加,但AF和20 Gy剂量均较对照在株高方面提高7%,且达到差异极显著(P<0.01)。这表明空间飞行和60Co-γ部分射线处理对苦荬菜的株高有较为显著的影响(表2)。
2.3 产量的变化
空间飞行和60Co-γ射线3种剂量(10,20,30 Gy)处理的产量差异显著(P<0.05),其中空间飞行处理的小区产量最高为15.91 kg,比对照(10.78 kg)高5.13 kg,达到极显著差异。60Co-γ射线(10,20,30 Gy)处理中,产量分别为10.57,11.78,9.04 kg,其中20 Gy处理产量高于对照外,其他2个处理均低于对照,且差异极显著(P<0.01),其中20 Gy剂量和AF均较对照在产量方面分别提高9%和48%(表3)。

表1 不同处理水平对M1代发芽势和发芽率的影响Table 1 Effects of germination potential and germination potential subjected to different treatments with different levels
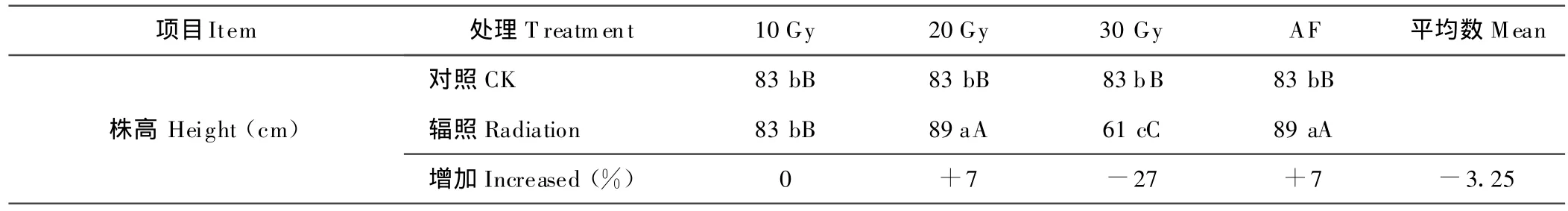
表2 不同处理水平对M1代株高的影响Table 2 Effects of height subjected to different treatments with different levels

表3 不同处理水平对M1代产量的影响Table 3 Effects of yield subjected to different treatments with different levels
2.4 品质构成因子的变化
空间飞行和60Co-γ射线辐射影响了苦荬菜的品质构成因子,表现在粗纤维含量、粗蛋白含量上存在显著差异(P<0.05)。试验表明,60Co-γ射线20 Gy剂量处理和空间飞行处理均能提高苦荬菜粗蛋白含量(表4)。

表4 不同处理水平对M1代品质因子的影响Table 4 Effects of different treatments with different levels on quality factors
2.5 变异系数分析
诱变M1代群体中各农艺性状和品质构成因子的遗传变异系数从大到小的顺序是:产量(21.5%)>粗纤维(20.0%)>株高(12.5%)>粗蛋白(11.9%)>发芽势(7.2%)>发芽率(5.4%)>粗脂肪(0.45%)。在这些指标中产量的变异系数最大(21.5%),说明这些特性选择的潜力大,通过选择容易达到预期目标。而其他特性的变异系数较低,被选择的几率有限,需扩大选择群体来提高选择效果[11]。
2.6 诱变处理对M1代苦荬菜农艺性状与品质构成因子的相关性影响
由于基因连锁和一因多效等使之不同性状间发生相关,空间飞行和60Co-γ射线处理苦荬菜突变株各个农艺性状和品质构成因子间的相关系数(表5)表明,各个农艺性状和品质构成因子的相关性显著(P<0.05)。试验表明,空间处理和60Co-γ射线辐射苦荬菜突变株农艺性状与品质构成因子的相关性更趋复杂、主要性状特征更趋加强。

表5 苦荬菜农艺性状、品质性状间的简单相关系数Table 5 Correlation coefficient of agronomic and quality characteristics of different treatment
2.7 诱变处理对M1代苦荬菜农艺性状与品质构成因子的主成分分析
主成分分析是通过对一组变量的几个线性组合来解释这组变量的方差和协方差结构,以达到数据的压缩和数据的解释的目的。从因子提取后的特征值可看出,提取前2个主成分可以解释86.446 1%的贡献。如果主成分分析中所提取主成分的特征值能达到85%以上的贡献率,就可以用这几个主成分对事物的属性进行概括性分析,基本可以得出影响事物性质的主要因素[13]。本试验的前3个成分的累积贡献率已达到86.446%,表明包含的信息已基本反应了各处理对苦荬菜诱变效应的影响。根据主要农艺性状的主成分矩阵显示,空间飞行和处理对M1代苦荬菜的影响主要在产量和品质2个方面。因此,以此为育种方向选育优质饲草品种具有可行性。
3 讨论
1)种子发芽过程本身是一个极复杂的生理生化过程[13]。空间飞行和60Co-γ射线处理均能提高苦荬菜M1代的发芽势和发芽率。
2)作为青绿饲料之一的苦荬菜的株高和产量研究较少,但紫花苜蓿的报道认为,在产草量上要有突破,应重点选择植株高大、茎粗、侧枝数多的材料[14]。苜蓿产草量与植株高度呈正相关[15]。因此,试验表明,空间飞行和60Co-γ射线处理均能提高苦荬菜M1代的株高和产量。
3)饲草粗蛋白含量是影响饲用价值的重要指标,蛋白质含量越高,饲用价值就愈大,能够提高.饲草中粗脂肪含量高,其适口性好,营养价值也高。饲草中粗纤维含量直接影响家畜对饲草的采食和消化率。干物质中粗纤维含量愈高,各类养分的消化率愈低反刍家畜的生产性能[16]。试验表明,空间飞行和60Co-γ射线辐射影响了苦荬菜的品质构成因子。
4)通过空间飞行(AF)和60Co-γ射线辐射处理,苦荬菜M1代突变群体的变异主要通过产量因子、品质因子2主成分体现出来,因此本试验根据主成分分析对突变株系的评价,以高产为主,综合性状好的品种作为苦荬菜辐射诱变后代群体的选择依据,选用空间飞行(AF)和60Co-γ射线20 Gy处理的后代作为育种材料,以此进行培育新品种。
[1]Mei M,Qiu Y,He Y,et al.Mutational effects of spaceflight on Zea mays seeds[J].Advances in Space Research,1994,14(10):33-39.
[2]李金国,蒋兴村,王长城.空间条件对几种粮食作物的同工酶和细胞学的影响[J].遗传学报,1996,23(1):48-55.
[3]刘丽,唐凤兰,张月学,等.空间飞行和60Co-γ射线辐照对苦荬菜种子的细胞学效应研究[J].草地学报,2007,15(5):469-472.
[4]王琳清.我国辐射育成的农作物品种[J].原子能农业应用,1985,(1):1-8.
[5]王培英,王连铮.大豆诱变育种及龙辐73-8955突变系的选育[J].大豆科学,1982,1(1):77-83.
[6]蒋昌顺,马欣荣,邹冬梅,等.应用微卫星标记分析柱花草的遗传多样性[J].高技术通讯,2004,(4):25-30
[7]鹿金颖,刘敏,薛淮,等.俄罗斯“和平”号空间站搭载的番茄随机扩增多态性DNA分析[J].航天医学与医学工程,2005,18(1):72-74.
[8]霍建泰,张廷纲,包文生,等.利用太空诱变、AFLP技术及日光温室加代选育航椒3号辣椒一代杂种[J].干旱地区农业研究,2007,25(5):85-88.
[9]王长山,徐香玲,张月学,等.不同熟期苦荬菜的染色体数目及核型分析[J].草业科学,2006,23(4):32-36.
[10]张月学,李成权,申忠宝,等.太空环境诱变苦荬菜的细胞学效应研究[J].草业科学,2007,24(9):38-41.
[11]韩建国.牧草种子学[M].北京:中国农业出版社,2000.
[12]杨胜.饲料分析及饲料质量检测技术[M].北京:农业大学出版社,1993:19-21,56-63.
[13]王赞,李源,吴欣明,高洪文,等.PEG渗透胁迫下鸭茅种子萌发特性及抗旱性鉴定[J].中国草地学报,2008,30(1):50-55.
[14]韩路,贾志宽,韩清芳,等.紫花苜蓿主要性状的对应分析[J].中国草地,2003,25(5):38-42.
[15]孙建华,王彦荣,余玲.紫花苜蓿生长特性及产量性状相关性研究[J].草业学报,2004,13(4):80-86.
[16]李孟良,郑琳,杨安中,等.播期、密度对“双低油菜”菜苔营养成分及菜籽产量的影响[J].草业学报,2008,17(3):137-141.
[17]黄迎新,周道玮,岳秀,等.不同苜蓿品种再生特性的研究[J].草业学报,2007,16(6):14-22.
[18]郭艳丽,郝正里,曹致中,等.不同生育期和不同品种苜蓿的果胶含量及与其他营养素的相互关系[J].草业学报,2006,15(2):74-78.
