一个番茄细胞分裂素受体类激酶基因的生物信息学分析
2009-01-27李爱芹李利斌刘国明
李爱芹 李利斌 刘国明
摘要:利用生物信息学的方法从番茄的基因组中解析得到一个与拟南芥渗透胁迫相关的细胞分裂素受体类激酶HK1高度同源的基因,命名为LeHK1,并对它的序列结构、蛋白基序、启动子区的顺式元件和基因的遗传进化进行了分析。 结果显示,番茄基因LeHK1编码的蛋白具有组氨酸蛋白激酶的结构特征,而且在进化上与拟南芥的渗透胁迫感受基因AtHK1分属同一个类群。从基因顺式反应元件分析来看,番茄基因LeHK1具有多个响应不同环境信号如热、干旱、激素信号的顺式元件,预示这个基因在环境响应和植物生长发育过程中具有重要功能。
关键词:番茄; HK1; 基因和蛋白结构; 顺式反应元件; 遗传进化
中图分类号:Q75文献标识码:ADOI编码:10.3969/j.issn.1006-6500.2009.05.001
Bioinformatics of A Cytokinin Receptor Like Kinase Gene in Tomato(Lycopersicum esculentum L.)
LI Ai-qin1, LI Li-bin2, LIU Guo-ming3
(1. High and New Technology Center of Shandong Academy of Agricultural Sciences,Jinan, Shandong 250100, China; 2. Vegetable Research Institute, Shandong Academy of Agricultural Sciences, Jinan, Shandong250100, China; 3.Taitou Town Comprehensive Agricultural Servicce Station of Shouguang City, Shouguang , Shandong 262735,China)
Abstract:Through bioinformatic method and datamining, a tomato homolog of arabidopsis histidine kinase gene HK1 which encodes a cytokinin recepter like protein, was isolated and named LeHK1. The structure ofLeHK1 sequence and its putative coding protein, aslo its cis-elements in promoter regioin and phylogenetic relasionship with other plants HKs was analyzed. The results showed that LeHK1 possesed the character of histidine kinase, and it was clusteredin the same group with AtHK1. According to the result of cis-elements analysis,tomato HK1 has muptiple cis-elements responsive to different environmental factors such as heat, drought, and hormones. This suggests that LeHK1 may play key roles in response to stresses and development.
Key words: Lycopersicum esculentum; HK1; gene and protein strtuctre ; cis-element; phylogeny
组氨酸蛋白激酶家族在植物乙烯、细胞分裂素和光信号响应过程中具有重要功能,参与和调节植物生长发育和对环境因子响应的众多过程。组氨酸蛋白激酶基因在模式植物拟南芥和水稻中得到了广泛和深入的研究[1-12], 但是在西红柿中的研究,尤其是对与细胞分裂素信号相关的组氨酸激酶基因的研究还很不充分。现有研究表明,细胞分裂素受体类蛋白在植物生长发育和逆境响应过程中具有极其重要的功能[1-3,5-15]。番茄作为世界上种植面积最大、也是我国最重要的蔬菜之一,在农业生产和人们的生活中具有不可替代的作用。因此,研究番茄细胞分裂素受体类蛋白基因对于改良番茄的抗逆性,以适应全球气候变暖和水资源日益匮乏,以及由于各种人为原因造成的土壤盐渍化等局面,力求生产更多的蔬菜,满足人们的生活需求具有重要意义。拟南芥渗透胁迫感受基因AtHK1(细胞分裂素受体类蛋白基因)已被克隆并得到功能验证[1,7],其直系同源基因在杨树、长春花中都已得到克隆和研究[13,16], 但在西红柿中还未见报道。因此,本研究拟利用拟南芥基因AtHK1的信息来解析番茄的同源基因序列。

1材料和方法
利用拟南芥的渗透胁迫感受基因AtHK1序列在Genbank中搜索相应的番茄基因组序列[17],根据内含子的边界特征解析得到编码区序列,利用编码区和基因组序列进行结构展示[18],并对预测编码的蛋白进行结构特征分析[19]和亚细胞定位预测[20]。根据基因组序列得到上游启动子区的序列,并对启动子序列中的顺式反应元件进行预测分析[21]。遗传进化分析利用MEGA4.1来进行,采用Bootstrap test-Neighbor Joining 方法,重复500次运算。
2结果与分析
2.1番茄LeHK1的序列和结构特征
本研究通过比较基因组学和生物信息学的方法从番茄基因组中得到一个与拟南芥HK1同源的基因组序列,命名为LeHK1。其预测编码的蛋白与拟南芥HK1的序列一致性为60.81%。LeHK1基因组序列长为5 261 bp,编码区为3 738 bp,预测编码一个含有1 245个氨基酸残基,分子量为138.86 kD的蛋白(表1)。这个基因位于番茄2号染色体的BAC克隆AC215474中,含有13个外显子,其外显子和内含子的结构如图1所示。

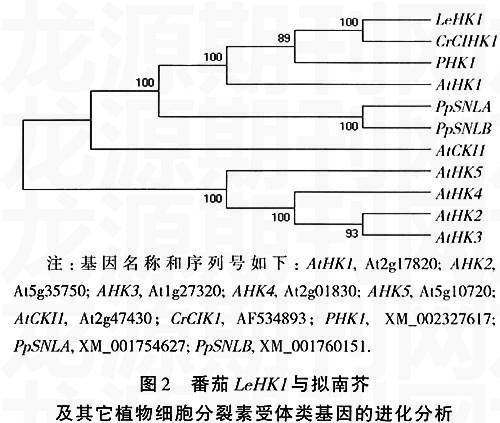
2.2 番茄LeHK1的进化分析
番茄LeHK1与拟南芥细胞分裂素受体相关的蛋白基因家族的进化分析表明(图2),LeHK1与AtHK1的关系最近, 推测与AtHK1有类似的功能,在渗透胁迫反应过程中具有重要作用。另外,LeHK1类基因在苔藓、杨树、长春花中都已得到克隆,而且在进化上分为同一个类群。长春花的CIHK1基因受冷胁迫诱导[16],因此,推测LeHK1可能在冷胁迫响应中也具有某种功能。功能性失活和获得研究表明,AtHK1能够正向调节植物对干旱、盐的响应和ABA的信号转导[1,2,7]。基因芯片分析的结果显示,突变体athk1中多个受逆境/ABA诱导表达的基因表达明显下降,这包括AREB1、ANAC和DREB2A及其调控的下游基因[1]。这说明,AtHK1基因作用于AREB1、ANAC和DREB2A的上游,可以通过依赖和不依赖ABA的信号途径正向调节逆境响应。另外,研究表明AtHK1在植物生长和种子成熟中也具有重要功能[1,7]。
2.3番茄LeHK1基因的顺式反应元件
通过分析启动子及其上游序列的顺式反应元件可以预测基因的功能。因此,本研究在得到LeHK1的基因组序列以后,取起始密码子上游2 000 bp的序列进行顺式元件分析,结果见表2。在LeHK1起始密码子上游2 000 bp的范围内共有10类顺式反应元件,包括赤霉素响应元件GARE和TATC-box,热激响应元件HSE,能和Myb蛋白结合的干旱诱导的顺式反应元件mybs,富含TC的逆境响应元件,昼夜节律响应元件,胚乳特异表达的顺式元件Skn-1,与防卫反应有关的顺式元件W-box和EIRE,还有Box-W1。这些结果表明,LeHK1基因可能不仅能够响应某些激素、热等非生物因子,而且与防卫反应和种子发育也有关系,值得深入探讨。
2.4番茄LeHK1蛋白的结构预测
为了研究LeHK1的功能,在解析得到它的编码序列后,对其预测编码的蛋白进行了结构分析。结果显示,LeHK1蛋白含有磷酸结合结构域、反应调节结构域、组氨酸激酶结构域、跨膜螺旋结构域和潜在的豆蔻酰化位点。这说明LeHK1蛋白具有组氨酸激酶的结构特征,而且有可能被豆蔻酰化。蛋白豆蔻酰化有利于与细胞膜的结合。亚细胞定位预测表明,LeHK1蛋白可能定位在细胞膜上,这与结构分析的结果相一致。
3讨论
细胞分裂素受体类激酶在植物逆境响应和生长发育过程具有重要功能。本研究解析的番茄基因LeHK1编码的蛋白具有组氨酸蛋白激酶的结构特征,而且在进化上与拟南芥的渗透胁迫感受基因HK1分属同一个类群,二者蛋白序列一致性达60.81%,而且亚细胞定位预测的结果表明LeHK1蛋白可能定位在细胞膜上。从这些分析结果来看,番茄基因LeHK1应为细胞分裂素受体类激酶基因,可能与拟南芥的渗透胁迫感受基因AtHK1具有相同或者相似的功能。另外,从基因顺式反应元件分析来看,番茄基因LeHK1 具有多个能响应不同环境信号如热、干旱、昼夜节律、病原物侵染,以及激素信号如赤霉素等的顺式反应元件,可对多种信号作出响应,预示这个基因在环境信号响应和植物生长发育过程中具有重要功能。至于番茄基因LeHK1对各种环境和激素信号的响应模式,在逆境响应和生长发育中的具体功能以及与细胞分裂素信号的关系需要深入探讨。本研究为进一歩研究番茄LeHK1基因的功能奠定了基础。
参考文献:
[1] Tran L P, Urao T, Qin F, et al . Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis[J].PNAS ,2007, 104: 20623-20628.
[2] Urao T, Yakubov B, Satoh R, et al. A transmembrane hybrid-type histidine kinase in arabidopsis functions as an osmosensor[J].The Plant Cell, 1999(11): 1743-1754.
[3] Higuchi M, Pischke M S, Mǎhǒnen, et al. In planta functions of the Arabidopsis cytokinin receptor family[J].PNAS, 2004,101: 8821-8826.
[4] Hwang I, Chen H C, Sheen J. Two-component signal transduction pathways in Arabidopsis[J].Plant Physiol, 2002,129:500-515.
[5] Nishimura C, Ohashi Y,Sato S, et al. Genetic analysis of Arabidopsis histidine kinase genes encoding cytokinin receptors reveals their overlapping biological functions in the regulation of shoot and root growth in Arabidopsis thaliana[J].Plant Cell ,2004(16):1365-1377.
[6] Desikan R, Horàk J, Chaban C, et al. The histidine kinase ahk5 integrates endogenous and environmental signals in Arabidopsis guard cells[J].PLoS ONE, 2008, 3(6): 2491.
[7] Wohlbach D J, Quirino B F, Sussman M R. Analysis of the Arabidopsis histidine kinase athk1 reveals a connection between vegetative osmotic stress sensing and seed maturation[J].Plant Cell, 2008(20):1101-1117.
[8] Iwama A, Yamashino T, Tanaka Y, et al. AHK5 histidine kinase regulates root elongation through an ETR1-dependent abscisic acid and ethylene signaling pathway in Arabidopsis thaliana[J].Plant Cell Physiol, 2007,48(2):375-380.
[9] Pareek A, Singh A, Kumar M, et al. Whole-genome analysis of oryza sativa reveals similar architecture of two-component signaling machinery with arabidopsis[J].Plant Physiology, 2006,142(2):380-397.
[10] Jain M, Tyagi K, Khurana J P. Differential gene expression of rice two-component signaling elements during reproductive development and regulation by abiotic stress[J].Functional & Integrative Genomics, 2008, 8(2): 175-180.
[11]Pareek A,Singh A, Kumar M, et al. Whole-genome analysis of oryza sativa reveals similar architecture of two-component signaling machinery with Arabidopsis[J].Plant Physiology,2006,142(2): 380-397.
[12] Ito Y, Kurata N. Identification and characterization of cytokinin-signalling gene families in rice[J].Gene, 2006(382):57-65.
[13] Chefdor F, Bénédetti H, Dpierreux C, et al. Osmotic stress sensing in populus: Osmoponents identification of a phosphorelay system[J].FEBS Letters, 2005(580): 77-81.
[14] Gonzalez-Rizzo S, Crespi M, Frugier F. The medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with sinorhizobium meliloti[J].The Plant Cell, 2004(18): 2680-2693.
[15] Coba de la Pe a T, Cárcamo C B, Almonacid L, et al. A salt stress-responsive cytokinin receptor homologue isolated from Medicago sativa nodules[J].Planta, 2008, 227(4):769-779.
[16] Papon N, Clastre M, Andreu F, et al. Expression analysis in plant and cell suspensions of CrCKR1, a cDNAencoding a histidine kinase receptor homologue in Catharanthus roseus (L.) G Don[J]. J Exp Bot,2002, 53 (376):1989-1990.
[17] Tatusova T A, Madden T L. Blast 2 sequences, a new tool for comparing protein and nucleotide sequences[EB/OL]. http://blast.ncbi.nlm.nih.gov/Blast.cgi,1999-01-01.
[18] Guo A Y, Zhu Q H, Chen X, et al. GSDS: A gene structure display server. [EB/OL]. http://gsds.cbi.pku.edu.cn/,2007-08-01.
[19] Gribskov M, Fana F, Harper J, et al. Plants P: A functional genomics database for plant phosphorylation[EB/OL]. http://plantsp.genomics.purdue.edu/html/feature_scan.html, 2001-01-01.
[20] Horton P, Park K J, Obayashi T, et al. Wolf psort: Protein localization predictor[EB/OL]. http://wolfpsort.org/, 2007-12-01.
[21] Lescot M, Déhais P, Thijs G, et al. Plant CARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences[EB/OL]. http://bioinformatics. psb.ugent.be/webtools/ plantcare/html/,2002-12-01.
