围栏封育对藏北高寒草地植物多样性与生态系统多功能性的影响
2025-03-06李振威缪雨珏宗宁





摘要:本研究选择藏北高原降雨梯度带四种类型高寒草地(高寒草甸、高寒草甸草原、高寒草原、高寒荒漠草原),通过测定围栏封育与自由放牧样地中与养分循环和牧草供给等功能密切相关的指标,利用平均值法和多阈值法来探讨围栏封育工程对生态系统多功能性的影响。结果表明:高寒草地植物多样性、物种丰富度和生态系统多功能性都会随降水量减少而降低(Plt;0.05)。围栏封育显著提高植物多样性、物种丰富度以及地上生物量。进一步分析发现,生态系统多功能性与Margalef指数、Simpson指数和Shannon-Weiner指数、物种丰富度均呈显著正相关(Plt;0.05),而与Pielou指数不相关。多阈值法显示围栏封育和自由放牧样地物种丰富度对生态系统多功能性的有效驱动分别在1%~84%和5%~82%阈值区间内,最大效应值分别是0.69和0.70。综上,围栏封育会使植物多样性发生变化,进而影响生态系统多功能性,保护物种多样性对于维持生态系统多功能性具有重要意义。
关键词:生态系统多功能性;植物多样性;藏北高寒草地;围栏封育与自由放牧;平均值法;多阈值法
中图分类号:S812""""""" 文献标识码:A""""""" 文章编号:1007-0435(2025)02-0596-13
Effects of Grazing Exclusion on Plant Diversity and Ecosystem Multifunctionality of Alpine Grasslands in Northern Xizang
LI Zhen-wei1,2, MIAO Yu-jue1,2, ZONG Ning1*
(1.Key Laboratory of Ecosystem Network Observation and Modelling, Institute of Geographic Sciences and Natural Resources Research,
Chinese Academy of Sciences, Beijing 100101, China; 2.University of Chinese Academy of Sciences, Beijing 100049, China)
Abstract:To investigate the impacts of grazing exclusion on the multifunctionality of ecosystems, 11 indicators related to nutrient cycling and productivity, including aboveground biomass, soil pH, soil bulk density, moisture content, total soil carbon, total soil phosphorus, available phosphorus, total soil nitrogen, inorganic nitrogen, ammonium nitrogen, and nitrate nitrogen, were selected and measured from grazing exclusion and free grazing sample plots of four grassland types (alpine meadow, alpine meadow steppe, alpine steppe, and alpine desert steppe) in the rainfall gradient zone in Northern Xizang. The average value method and multi-threshold method were utilized to explore the effects of grazing exclusion on the multifunctionality of ecosystems. The results showed that plant diversity, species richness and ecosystem multifunctionality of alpine grasslands all decreased significantly with decreasing precipitation (Plt;0.05). Grazing exclusion increased plant diversity and richness as well as aboveground biomass. Ecosystem multifunctionality was significantly correlated with Margalef richness index, Simpson dominance index, Shannon-Weiner diversity index and species richness (Plt;0.05), while ecosystem multifunctionality showed a non-significant correlation with Pielou evenness index. Multi-threshold method demonstrated that species richness drove the ecosystem multifunctionality within the thresholds of 1%-84% and 5%-82% inside and outside the fence effectively, respectively, and the maximum effect values were 0.69 and 0.70, respectively. In conclusion, plant diversity might alter under fence closure, which in turn affected ecosystem multifunctionality. Thus, the protection of species diversity is important for the maintenance of ecosystem multifunctionality in alpine ecosystems.
Key words:Ecosystem multifunctionality;Plant diversity;Alpine grassland in northern Xizang;Grazing exclusion and free grazing;Mean value method;Multi-threshold method
青藏高原拥有我国面积最大的草地资源,其天然草原面积达到了13.2亿亩,是我国的五大牧区之一[1]。青藏高原高寒草地在调节气候、水土保持、保护生物多样性等方面发挥着重要的作用,是我国重要的生态安全屏障[2-3]。然而,青藏高原的原生草地在气候变化和人类活动(如过度放牧等)的影响下,有19%~60%的草地出现了不同程度的退化[4-5]。青藏高原草地的退化不仅造成了植被生产力的下降、生物多样性的损失、土壤养分和有机质的降低,还对牧民的生计、当地的经济效益和社会发展产生了严重的影响[5-6]。长久以来,如何进行退化高寒草地的恢复一直是科学家和牧民关注的问题。围栏封育被认为是一种成本低但效果显著的恢复退化草地的重要措施[7-8]。围栏封育可以暂时的隔绝人类活动对草地的干扰,从而有效的改善过度放牧给草地带来的负面影响。围栏封育对于草地植被群落的生物量和土壤中的养分含量都有着显著提升的作用[9],对于草地中的物种多样性方面也有不同程度的提升作用[10-11]。鉴于草原在生态系统服务功能中发挥的重要作用,因此我们需要更好的探究围栏对于草地生态系统功能的影响。
生态系统功能(Ecosystem function)即生态系统的不同生境、生物学及其系统的性质或过程[12-13]。近几十年来,相关学者致力于生态系统功能的研究,尤其是生物多样性与生态系统功能之间的关系成为研究热点[14-15]。随着研究的深入,许多关于生物多样性与生态系统功能之间关系的成果被发表[16-20]。然而,现有研究大多只关注生产力、凋落物分解、营养循环、土壤养分等单一或少数生态系统功能与生物多样性之间的关系[21]。通过研究单个生态系统功能代表整体的功能会忽略生物多样性对其他生态系统功能产生的影响,无法全面反映生物多样性对生态系统整体的影响[21-23]。因此,生态系统多功能性(Ecosystem multifunctionality,EMF)的概念被提出。自2007年Hector等[24]首次量化生态系统多功能性后,对生物多样性与生态系统多功能性之间关系的研究逐渐成为生态学领域的热点议题[16,25-26]。此后,EMF被作为评估草地生态系统多功能性的指标,因为它可以将生态系统的整体功能综合为一个简单的、可以用来对比的指标[27-28]。目前对于生态系统多功能性的计算方法主要有以下四种:单一功能法、功能-物种替代法、平均值法、阈值法。其中使用较多的是平均值法,但这一方法忽略了各个生态指标之间所占权重的不同[29-31]。因此,采用不同方法进行生态系统多功能性的计算显得尤为重要。
目前,前人的研究大都集中在同一草地类型上围栏内外的差异,而对于不同草地类型之间的研究相对较少。然而围栏对不同的草地类型的影响各不相同。此外,前人对于生态系统功能的研究也主要集中在单一功能上。因此,本文基于藏北高原降雨梯度带四种类型高寒草地(高寒草甸、高寒草甸草原、高寒草原、高寒荒漠草原)围栏内外的群落,选择土壤养分循环和储存能力、牧草供给能力、土壤水源涵养能力等多方面的指标,使用平均值法和多阈值法来量化围栏封育工程对高寒生态系统多功能性的影响。通过分析围栏封育工程对不同草地类型生物多样性与生态系统多功能性关系的影响,以期为生态工程实施下不同类型高寒草地生态系统的管理提供科学依据。
1 材料与方法
1.1 研究区概况
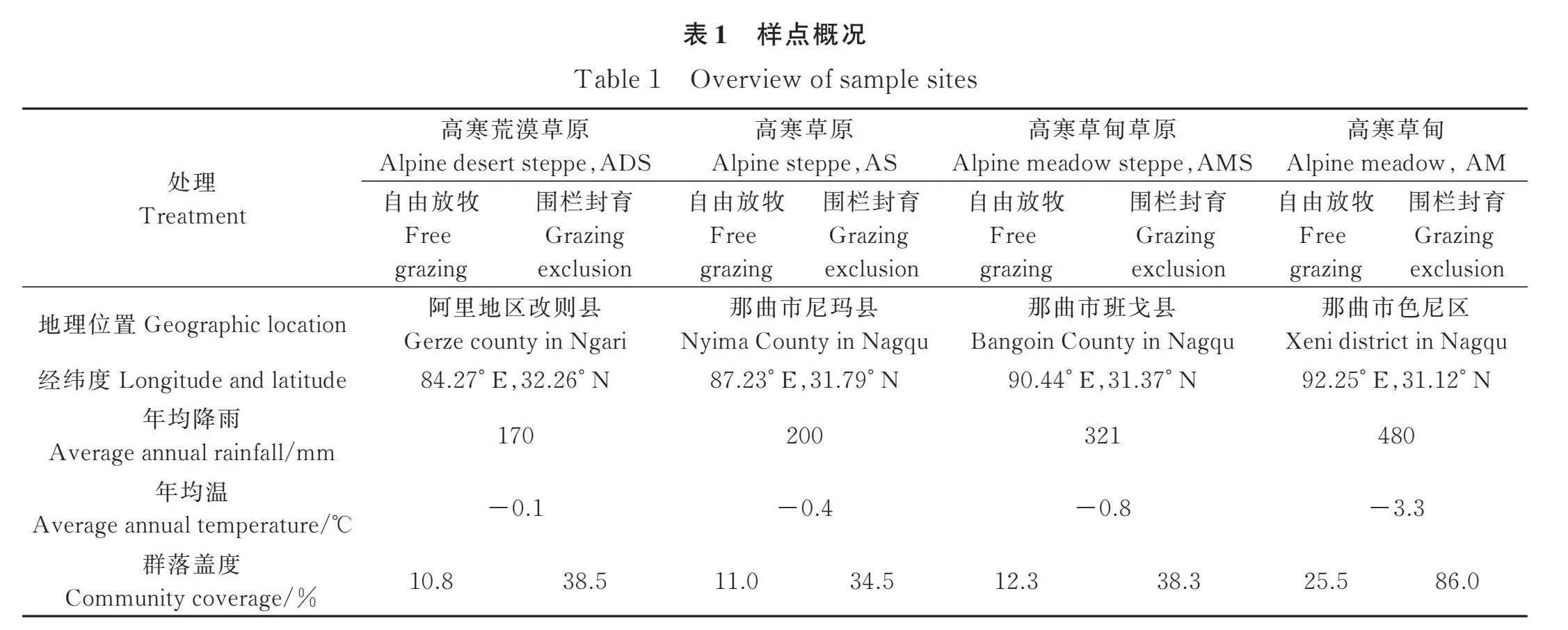
本文的采样地点自东向西依次位于藏北高原的那曲市色尼区、班戈县、尼玛县以及阿里地区改则县(31.119°~32.259° N,84.273°~92.253° E),海拔在4400~4700 m。从东向西年降水量自那曲地区的480 mm降低至阿里地区的170 mm,其中85%以上的降雨分布在6—8月,年平均气温在-3.3~-0.1℃。研究区域草地类型从东向西依次为高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原[32]。东部高寒草甸的主要优势物种为高山嵩草(Carex parvula O.),伴生种有二裂委陵菜(Potentilla bifurca Linn.)、窄叶苔草(Carex montiseverestii Kük.)等,而中西部高寒草甸草原、高寒草原和高寒荒漠草原的优势种主要是紫花针茅(Stipa purpurea Griseb.),但各个草地类型的伴生种都各不相同:高寒草甸草原的伴生种主要是羊茅(Festuca ovina L.)、冷地早熟禾(Poa crymophila Keng.)、线叶苔草(Carex capilliformis Franch.)等;高寒草原的伴生种主要是丛生黄芪(Astragalus confertus Benth.)、青藏狗娃花(Aster boweri Hemsl.)等;高寒荒漠草原的伴生种主要是小叶棘豆(Oxytropis microphylla Bunge.)、燥原荠(Stevenia canescens DC.)等。随着植被类型的变化,土壤类型也呈现梯度变化特征,高寒草甸土壤类型为高山草甸土(黑毡土),高寒草甸草原和高寒草原土壤类型为高山草原土(莎嘎土),高寒荒漠草原土壤类型为高山荒漠化草原土(高山草原土的亚类,也称寒漠土)[33]。
1.2 试验设计与植被调查
在2023年8月上旬植株生长旺盛期,在研究区域内进行植被调查和样品采集。根据我们前期的观测和相关的研究表明,自由放牧样地放牧强度在轻度到中度之间,围栏封育从2012年开始[34]。在4种类型高寒草地中的围栏内外分别设置1个样地,共设置8个样地。每个样地中沿着围栏设置4个样方作为重复,样方之间间隔在10 m左右。采用样方法(0.5 m×0.5 m)测量样方中的植被总盖度、物种数量、物种盖度以及每个物种的个体数。完成植被调查后将植物地上部分齐地剪下放置于信封中,带回实验室进行地上生物量的测量。将植物材料置于65 ℃烘箱中烘干48小时至恒重后称重,换算到每平米作为地上生物量。用直径为3.8 cm的土钻获取样方内0~10 cm,10~30 cm两层土壤,每个样方内取五钻(分别在样方的四角和中心)混合为一个土壤样品。土壤过2 mm筛后分为两份,一份带回实验室风干,一份冷藏保存,留作进一步指标测定与分析。
1.3 植被与土壤因子测定与分析
关于生态系统功能的测定共选择了11个指标:地上生物量(Aboveground biomass,AGB)、土壤pH值、土壤容重(Bulk density,BD)、土壤含水量(Soil moisture, SM)、土壤全碳(Total carbon,TC)、土壤全磷(Total phosphorus,TP)、土壤速效磷(Soil available phosphorus,AP)、土壤全氮(Soil total nitrogen,TN)、土壤无机氮(Soil inorganic nitrogen,SIN)、土壤铵态氮(Soil ammonium nitrogen,NH4+-N)、土壤硝态氮含量(Soil nitrate nitrogen,NO3--N)[35]。使用酸度计测量土壤pH,土水比为1∶2.5。土壤含水量采用烘干法,新鲜土壤置于烘箱中105 ℃烘干48小时至恒重后称重。土壤容重采用环刀法测定。土壤全氮和全碳含量采用元素分析仪测定。土壤全磷用HClO4-H2SO4消解-钼锑抗比色法测定(GB7852-87)测定。土壤无机氮(包括铵态氮和硝态氮)采用流动分析仪测定。土壤速效磷含量采用NaHCO3浸提-钼锑抗比色法测定。上述指标综合了土壤养分循环、土壤涵养水源能力、牧草供应能力等多种生态系统功能。
1.4 指标计算方法
对于植物物种多样性的计算选取Margalef丰富度指数、Simpson优势度指数、Shannon-Weiner多样性指数、Pielou均匀度指数和物种丰富度五个指标来表征。计算公式如下[35]:
(1)Margalef丰富度指数:
M=(W-1)/ln N
(2)Simpson优势度指数:
S=1-∑_(i=1)^WQ_i^2
(3)Shannon-Weiner多样性指数:
D=-∑Q_i lnQ_i
(4)Pielou均匀度指数:
P=H/lnS
(5)物种丰富度指数:
R=W
式中,W为样方内物种数;N为样方内全部物种个数;i为第i个物种;Qi为样方内第i个物种的个体数所占所有物种的个体数的比例。
选取地上生物量、土壤pH值、土壤容重、土壤含水率、土壤全碳、土壤全磷、土壤速效磷、土壤全氮、土壤无机氮、土壤铵态氮、土壤硝态氮含量11种指标来计算生态系统多功能性。对于生态系统多功能性的计算选择了平均值法和多阈值法两种方法。平均值法首先是将选取的生态系统多功能指标参数值标准化,然后再使用Z得分法计算11个生态系统多功能指标的Z分数,最后求得Z分数的平均值即为EMF值。Z得分法的公式和EMF值的计算公式如下[33]:
Z_ab=(F_ab-μ_b)/σ_b
T_EMF=∑_(b=1)^11〖Z_ab/10〗
式中,Zab为样地a第b种生态系统功能参数的Z分数;Fab为样地a第b种生态系统功能参数的数值;μb为第b种生态系统功能参数全部样地间的平均值;σb为第b种生态系统功能参数在全部样地间的标准差。
多阈值法是指通过设置阈值,计算超过阈值的功能数来求得多功能性指数。计算公式如下[29]:
MF_t=∑_(i=1)^F〖(r_i (f_i )gt;t_i)〗
式中,F是功能总数,fi是生态系统中某一功能i的值,ti表示阈值。该方法中各项指标含义如下:
(1)最小阈值(Tmin):多样性开始产生影响的最低阈值(即显著大于或小于0的斜率)。
(2)最大阈值(Tmax):斜率首先下降到与0没有显著差异的阈值。
(3)最大多样性效应阈值(Tmde):多样性具有最强正或负影响的阈值。
(4)多样性最大效应(Rmde):多样性具有最强的积极或消极影响的关系强度(即斜率)。
(5)最小阈值处的多功能性(Mmin):达到Tmin时的功能数。
(6)最高阈值处的多功能性(Mmax):达到Tmax时的功能数。
(7)多样性最大化时的多功能性(Mmde):达到Tmde时的功能数。
(8)最大可能多样性效应的百分比(Pmde):Rmde除以实验中可能出现的最大斜率(总功能数除以总物种数),表示多样性对多功能性的相对重要性在实验中实现的百分比。
1.5 统计与分析
使用Excel 2019进行数据整理和计算,包括Margalef丰富度指数、Simpson优势度指数、Shannon-Weiner多样性指数、Pielou均匀度指数和重要值。之后再用Origin 2024进行画图。利用SPSS 27.0.1进行方差齐性检验、T检验和ANOVA单因素方差分析。后利用R 4.3.3软件的multifunc包对生态系统多功能性进行多阈值法分析并画图。本文所有分析的显著性水平为Plt;0.05。
2 结果与分析
2.1 围栏封育对不同类型高寒草地物种多样性和地上生物量的影响
由图1可知,围栏封育对不同类型高寒草地的影响不同。在高寒草甸和高寒荒漠草原中,围栏封育显著提高了地上群落生物量(Plt;0.05),而物种多样性的各项指数并没有显著变化。在高寒草甸草原中,围栏封育显著提高了物种丰富度、Margalef丰富度指数和群落地上生物量(Plt;0.05)。在高寒草原中,围栏封育使物种丰富度、Margalef丰富度指数、Simpson优势度指数和Shannon-Weiner指数均出现显著增加的趋势(Plt;0.05)。
围栏封育与自由放牧区域不同类型的高寒草地物种丰富度和多样性指数呈现不同的规律。在自由放牧区域,物种丰富度、Margalef丰富度指数、Simpson优势度指数、Shannon-Weiner指数和Pielou均匀度指数的最大值均出现在高寒草甸草原,其次是高寒草甸。相对于高寒草甸和高寒草甸草原的物种多样性,高寒草原和高寒荒漠草原的物种多样性则显著降低(Plt;0.05)。对于群落地上生物量,高寒草甸相较于其他三种草地类型显著增加(Plt;0.05)。在围栏封育区域,物种丰富度、Margalef丰富度指数、Simpson优势度指数、Shannon-Weiner指数、Pielou均匀度指数都以高寒草甸草原最高。相较于高寒草甸草原,物种丰富度、Margalef丰富度指数、Simpson优势度指数、Shannon-Weiner指数、Pielou均匀度指数在高寒草甸呈现下降的趋势但并不显著,而在高寒荒漠草原和高寒草原中,除Pielou均匀度指数外都显著下降(Plt;0.05);群落地上生物量则以高寒草甸最高,且与其他草地类型都有显著差异(图1)。
2.2 围栏封育对不同类型高寒草地土壤理化性质的影响
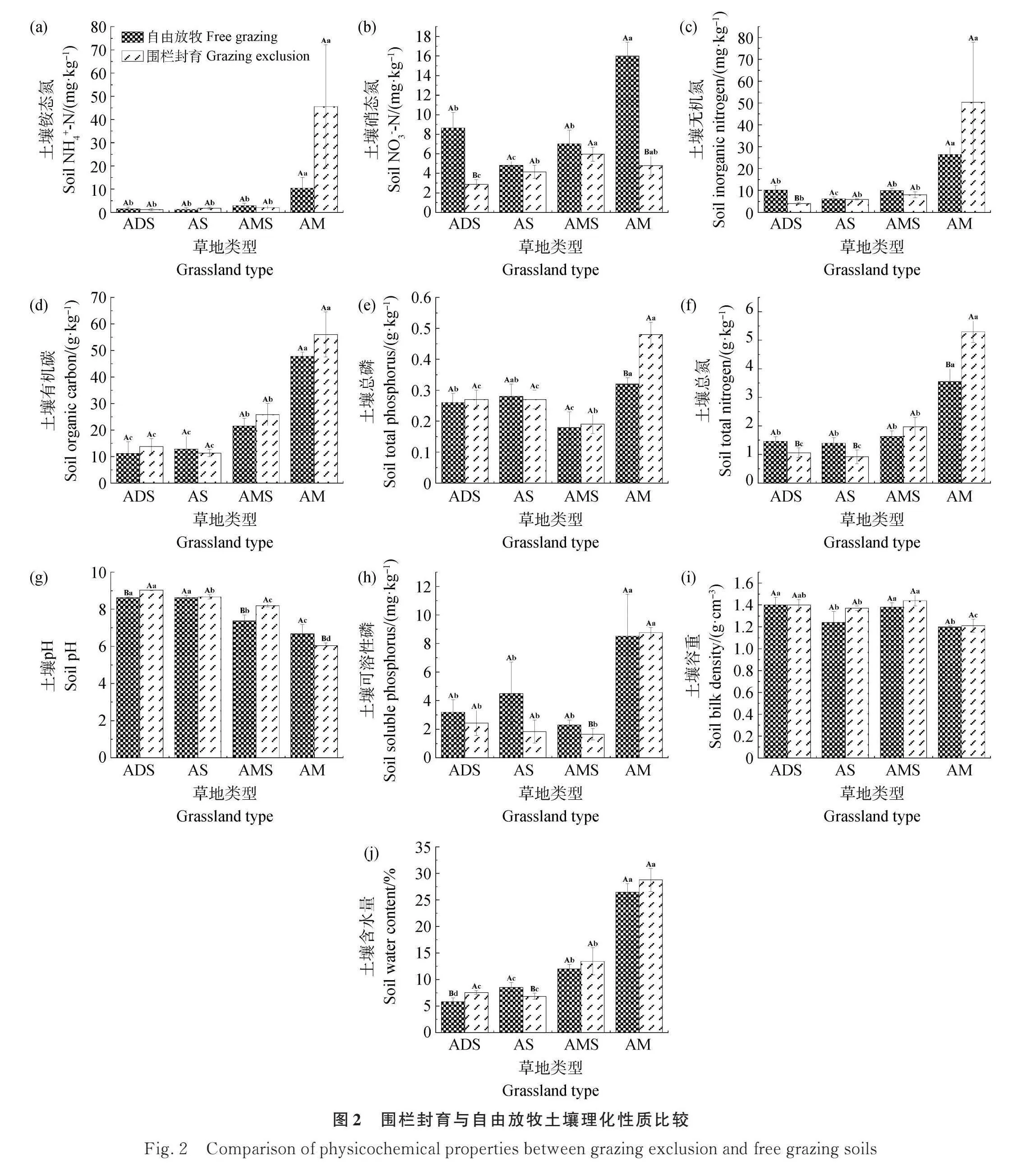
由图2可以看出,围栏封育对不同类型高寒草地土壤理化性质的影响不同。在高寒草甸中,围栏封育显著提高土壤全磷和全碳含量,而降低土壤硝态氮和pH值(Plt;0.05)。在高寒草甸草原中,围栏封育显著降低土壤速效磷的含量,但显著提高土壤pH值(Plt;0.05)。在高寒草原中,围栏封育显著降低土壤全氮和土壤含水率(Plt;0.05)。在高寒荒漠草原中,围栏封育显著降低了土壤硝态氮、土壤无机氮、土壤全氮和土壤含水量,但显著提高土壤pH(Plt;0.05)。
在自由放牧区域,沿降雨梯度的下降土壤养分含量呈降低趋势。具体来说,土壤铵态氮、硝态氮、无机氮、全碳、全磷、全氮、速效磷的含量和土壤含水率的最高值均出现在高寒草甸中,而最低值均出现在高寒荒漠草原或高寒草原中,且高寒草甸与高寒荒漠草原、高寒草原均存在显著差异(Plt;0.05),而土壤pH则正好相反(Plt;0.05)。土壤容重在高寒荒漠草原和高寒草甸草原中最高,在高寒草原和高寒草甸中最低,且前两种与后两种草地类型之间差异显著(图2,Plt;0.05)。
土壤养分含量在围栏封育区域的变化趋势同自由放牧区域相似。高寒草甸的土壤铵态氮、土壤无机氮、土壤全碳、土壤全氮、土壤全磷、土壤速效磷含量和土壤含水率最高,而土壤容重和pH值则是所有草地类型中最低的,且上述土壤理化性质与其他类型草地皆有显著差异(Plt;0.05)。高寒草甸草原的土壤硝态氮含量和土壤容重最高;在高寒荒漠草原和高寒草原中则出现土壤pH值的最大值,而其他土壤理化性质与高寒草甸相比基本都有显著差异(图2,Plt;0.05)。
2.3 围栏封育对高寒草地生态系统多功能性的影响
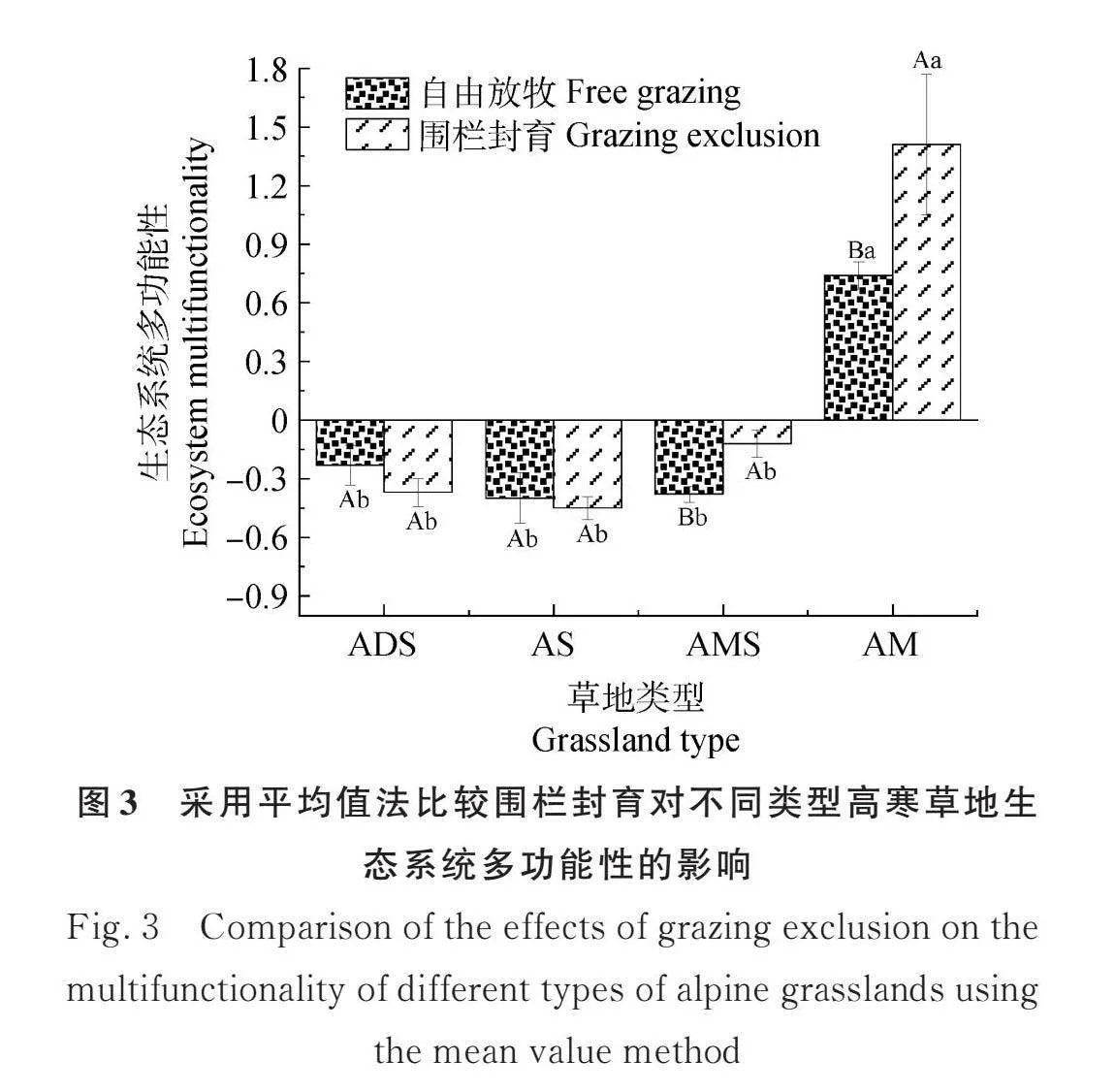
采用平均值法计算生态系统多功能性指数,对比不同草地类型围栏封育和自由放牧的生态系统多功能指数。由图3可知,在高寒草甸和高寒草甸草原中,围栏封育区域的生态系统多功能性指数分别为1.41和-0.12,二者均显著高于自由放牧区域的生态系统多功能性指数(Plt;0.05)。但在高寒草原和高寒荒漠草原中,围栏封育区域的生态系统多功能性指数为-0.45和-0.37,与自由放牧区域的-0.40和-0.23没有显著差异。对比不同类型高寒草地的生态系统多功能性指数,无论围栏内外,高寒草甸的生态系统多功能性指数都以唯一的正值位列第一,而最低值则出现在高寒草原,围栏封育和自由放牧分别为-0.45和-0.37。
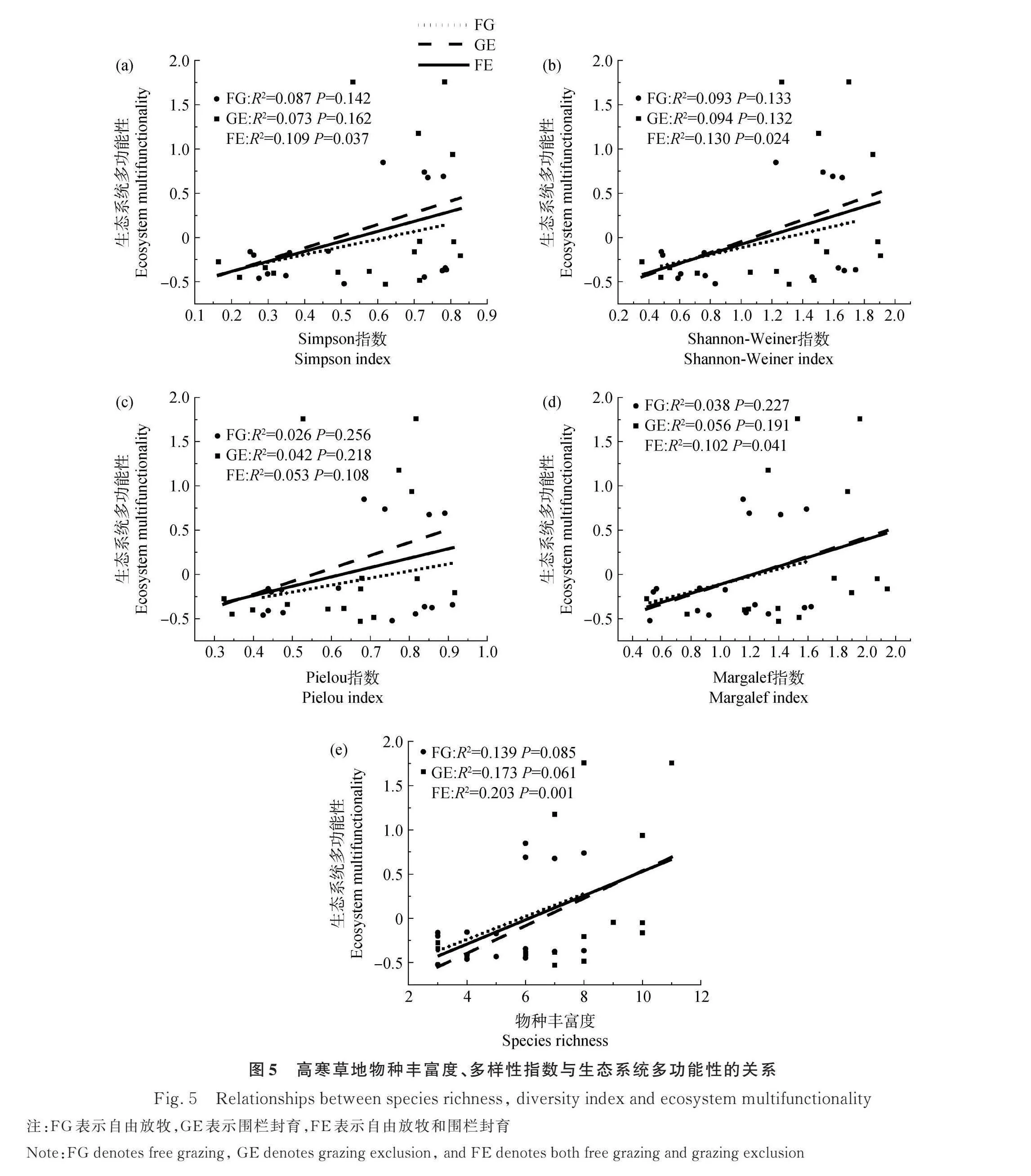
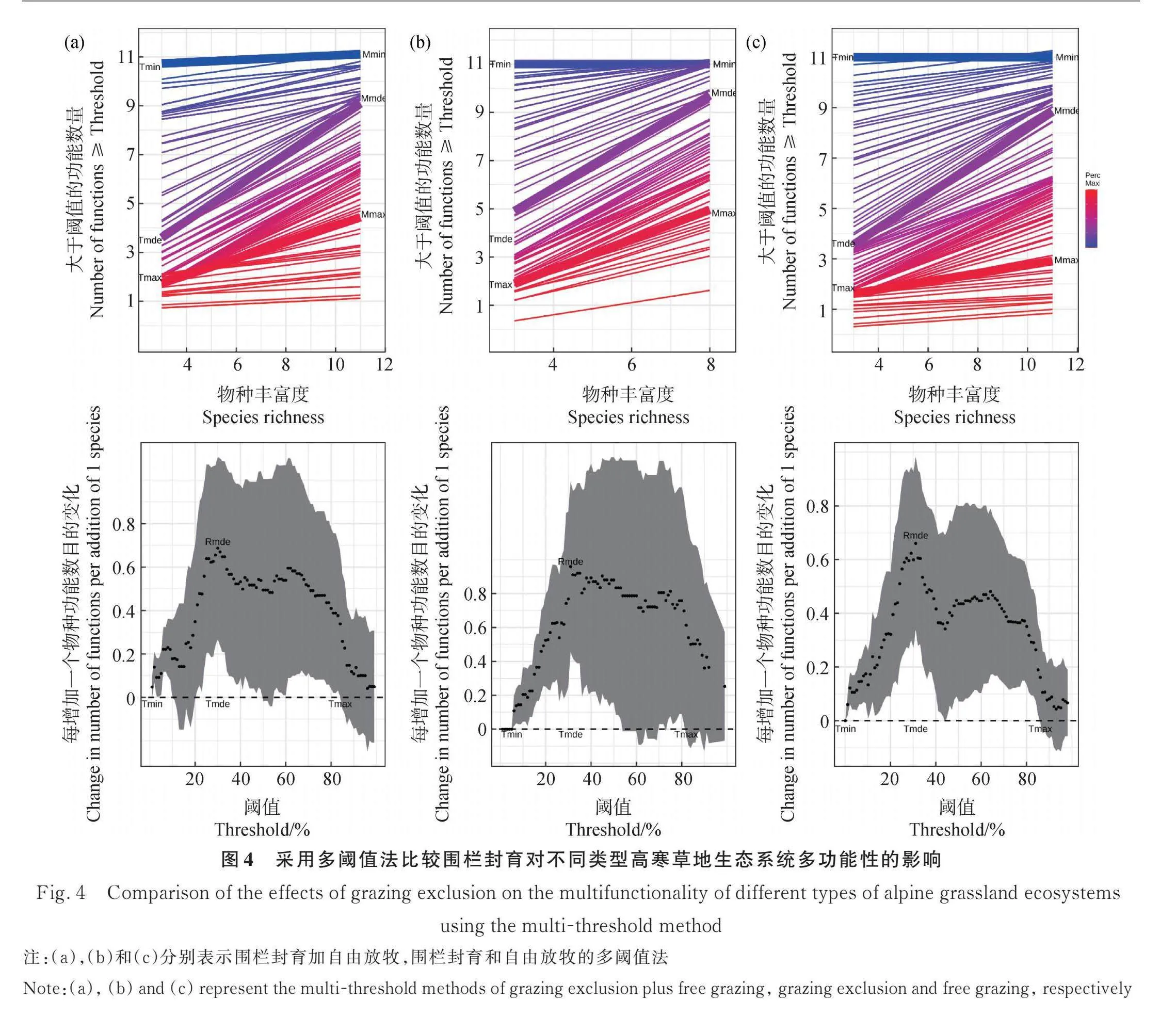
围栏封育的多阈值法结果表明,在阈值为1%(Tmin)和84%(Tmax)之间,物种丰富度和多功能性之间呈正相关。在阈值为30%(Tmde)时,这种正相关关系达到顶峰,斜率为0.68,这意味着每增加一个物种,功能数增加0.68。对于本研究的多样性水平范围(3-11个物种)和测量的生态系统多功能性指标(11),物种丰富度与大于阈值的多功能性指标之间关系的最大可能斜率为1。因此在关系最强的情况下,在本研究中物种多样性对多功能性最大的可能影响为69%(Pmde)(图4)。
自由放牧的多阈值法结果表明,在阈值为5%(Tmin)和82%(Tmax)之间,物种丰富度和多功能性之间呈正相关。在阈值为31%(Tmde)时,这种正相关关系达到顶峰,斜率为0.96,每个物种可增加0.96个功能。对于本研究的多样性水平范围(3-8个物种)和测量的生态系统多功能性指标(11),物种丰富度与大于阈值的多功能性指标之间关系的最大可能斜率为0.73。因此在关系最强的情况下,本研究中物种多样性对多功能性最大的可能影响为70%(Pmde)(图4)。
将围栏封育和自由放牧综合起来进行分析发现,在阈值为1%~86%的范围里,物种丰富度和多功能性之间呈正相关关系(图4)。在阈值为31%时(Tmde),物种丰富度和多功能性之间的正相关关系达到最高,斜率为0.66,这说明每个物种可增加0.66个功能数。对于本研究的多样性水平范围(3~11个物种)和测量的生态系统多功能性指标(11),物种丰富度与大于阈值的多功能性指标之间关系的最大可能斜率为1。因此在关系最强的情况下,多样性在本研究中对多功能性最大的可能影响为66%(Pmde)。
2.4 物种多样性与生态系统多功能性的关系
由图5可知,无论是在自由放牧还是围栏封育处理,物种多样性与生态系统多功能性之间的关系均不显著。综合两种处理的样点发现,物种丰富度、Margalef丰富度指数、Simpson优势度指数和Shannon-Weiner多样性指数均与生态系统多功能性呈现显著正相关关系(Plt;0.05),其中物种丰富度的拟合效果最好,解释量达到20.3%。Pielou均匀度指数则与EMF没有显著相关关系(P=0.108)。由此可以看出,物种丰富度越高,高寒草地生态系统多功能性就越高。
3 讨论
3.1 围栏封育对不同类型高寒草地植物群落组成、土壤理化性质和生态系统多功能性的影响
本研究在青藏高原上选取了四种草地类型(高寒草甸、高寒草甸草原、高寒草原、高寒荒漠草原)进行植物群落组成、土壤理化性质和生态系统多功能性的分析。研究发现,降水增加显著的促进了植物的物种多样性和生态系统多功能性。这与胡亚等[36-37]研究结果一致。降水通过影响植物物种多样性和土壤理化性质来改变生态系统多功能性[38-39]。一方面,在干旱环境下,水分胁迫是限制植物生长的主要因素,土壤水分的不足会限制土壤微生物的正常活动,从而影响植物对于土壤养分的吸收和利用。另一方面,降水会改变土壤理化性质和土壤环境,促进生态位的分化,使更多不同的物种(例如更适应湿润环境的植物)可以有更好的生长空间,从而控制群落演替的进程[40]。研究表明,降水的增加使土壤容重和pH降低,而土壤养分和含水率均会增加。Chytry等[41]的研究表明土壤pH与降水量呈现负相关的关系,郭志霞等[42]研究发现随着降水的增加,土壤容重会下降,而土壤养分相对增加,这与我们的研究结果一致。降水的增加促进了土壤淋溶,土壤中的可溶性物质被淋溶到更深层的土壤中,从而改变了表层土壤中的离子平衡,降低了土壤pH。土壤pH的变化也会因为降水导致非酸阳离子浸出增加[41]。土壤养分含量的增加可能是因为降水促进干旱土壤中的植物生物量增加,也促进了降水对于植物凋落物的物理破碎过程。另一方面,降水促进了土壤微生物对凋落物的分解,从而增加土壤养分含量[43]。部分前人的研究证明降水量增加可能会降低土壤养分含量,这可能是因为随着降水的过多导致土壤养分淋溶增加引起的[44]。
围栏封育是一种应用广泛且常见的退化草地恢复技术,它可以有效的减少过度放牧的影响,有利于草地的自然恢复[45]。本研究表明,与自由放牧草地相比,围栏封育草地的地上生物量有显著的增加。草地围栏封育会有效的减少人类活动和放牧对于草地的影响,使牧草能有充分恢复和生长的时间。围栏的使用会减少牲畜对于草地的采食和踩踏,使得群落植被盖度显著增加[46]。本研究围栏封育处理中Margalef丰富度指数、Shannon-Wiener指数和物种丰富度均出现不同程度的增加,这说明围栏封育使群落结构的复杂性增加,这是因为围封之后植物群落得以在稳定的环境中生长。此外,一项关于围栏封育对草地植物多样性影响的Meta分析也表明,短期围栏封育对于草地物种丰富度有促进作用[47],但围栏封育对于草地物种多样性的促进作用随时间的增加而减少,整体呈现先促进后降低的过程[48-49]。由于我们只进行了一次采样,并不能观察到物种多样性随时间的变化,未来的研究会针对长期围栏封育对物种多样性的影响进行研究。任毅华等[50]的研究表明,围栏内Margalef丰富度指数、Shannon-Wiener指数和Simpson指数较围栏外有增加的趋势,但Pielou均匀度指数会下降,这与我们的结果略有不同。这是因为任毅华等[50]选择的研究区域,植物群落之间存在较大的竞争,而围栏封育之后优势物种的竞争力增强,主导了该区域初级生产力的分配,从而抑制了其他物种的生长[51]。
土壤理化性质改善是草地恢复的重要指标。对比围栏封育和自由放牧土壤的各项理化性质,围栏封育处理土壤含水率和容重都有增加的趋势,在高寒草原和高寒草甸草原更显著,这与周天阳等[52]关于8年围栏封育对青藏高原东南缘高山草地的综合评价结果一致。这主要有两个原因,一是因为围栏的使用最大程度的隔绝了牲畜对于土壤的影响,使土壤可以进行自我恢复[52];二是围栏封育后新增的植被覆盖在土壤上,从而减少土壤水分的蒸发[53]。在高寒荒漠草原、高寒草原、高寒草甸草原三种草地类型中,围栏封育处理的土壤硝态氮、土壤无机氮、土壤有效磷都出现下降的趋势,这是因为围栏隔绝家畜对草地的影响,减少了家畜粪便中氮素的输入,从而降低了土壤中氮的含量[54];而土壤有效磷的下降则是因为由于围栏封育促使植物生物量的增加,从而促进了植物对于磷的吸收[54]。在这三种草地类型中,围栏封育处理的土壤pH呈现增加的趋势,因为随着植被的增加,土壤水分的蒸发减少,土壤中的盐基离子随着水分而积累在土壤表层从而使pH增加。土壤铵态氮在高寒荒漠草原和高寒草甸草原的围栏内略微下降,但在高寒草原和高寒草甸的围栏内有所增加,这是由多方面原因造成的:一方面随着土壤pH的增加,铵态氮相对更加稳定;另一方面在不同的草地类型中,土壤微生物的组成存在差异[55],对于铵态氮的转化能力也不同。土壤全碳在高寒荒漠草原和高寒草甸草原围栏封育处理更高,在高寒草原自由放牧处理更高。土壤全氮在高寒荒漠草原和高寒草原围栏封育处理都有下降的趋势,在高寒草甸草原中有增加的趋势。这是因为在不同草地类型中物种组成不同,对养分的种类和需求也不同[56]。我们的研究与前人关于围栏对土壤理化性质的影响的研究有一定的差异性。刘建康等[57]研究发现,由于围封的原因,土壤植被逐渐恢复,增加了植被对降水的拦截作用,从而降低了土壤被风蚀的情况,使围封草地的各个土壤养分指标均高于自由放牧草地。
平均值法的结果表明,在高寒草甸和高寒草甸草原中围栏对于生态系统多功能性有显著的提高,但在高寒荒漠草原和高寒草原无显著影响。这与Yu等[58]的研究结果一致,只有在干旱程度较低的草地上,围栏封育才会促进多功能性,这主要与干旱条件下围栏封育限制了植物生物多样性维持生态系统多种功能的能力有关。同时,在内蒙古典型草原的研究结果也表明,围栏封育通过提高养分含量来增强生态系统多功能性[59],这与我们的研究结果相一致。阈值法的结果表明,无论围栏封育还是自由放牧处理通过增加物种多样性不可能同时使所有功能都达到最理想的程度,这与德国BIODEPTH站点的研究结果一致[30]。虽然生物多样性确实会增加生态系统多功能性,并且在较高的阈值下也有显著的促进作用,然而在高阈值下斜率呈下降趋势,这表明高物种丰富度并不能保证所有功能表现在最高水平。围栏封育使物种丰富度对生态系统多功能性产生效应的阈值范围略有缩小,这是因为围栏的使用使物种的多样性会增加,增加的物种中对于生态系统有负面作用的物种也会增加,从而导致物种丰富度对生态系统多功能性产生效应的阈值范围缩小[47]。但物种丰富度对生态系统功能产生的最大效应有显著的增加,这是因为围栏封育使群落的组成发生改变,使能最大程度促进生态系统功能的优势物种增加[50]。
3.2 植物群落组成与生态系统多功能性的关系
在青藏高原的各种草地类型,不同的物种多样性指数与生态系统多功能性指数关系存在差异。本研究发现,除Pielou均匀度指数外,物种丰富度、Margalef丰富度指数、Simpson优势度指数和Shannon-Weiner多样性指数均与生态系统多功能性呈现显著正相关关系,其中物种丰富度与生态系统多功能性指数的线性回归关系最为显著(图5)。这表明,在青藏高原高寒草地中物种数量是影响生态系统多功能性的主要因素,物种的丰富度、优势度和多样性对生态系统的多功能性也具有促进作用,但物种均匀度对生态系统多功能性并没有显著的作用。本研究结果与张文馨等[60]、熊定鹏等[33]和Vander Plas等[61]已有报道一致。以上这些研究都说明了物种多样性对于生态系统功能具有一定程度的正效应。基于物种之间的互补效应,物种多样性的增加会促进生态位的分化,从而降低物种之间的竞争关系,使更多功能互补的物种在环境中发挥作用。
此外,本研究发现围栏封育12年后生态系统多功能性的提升主要集中在降雨较多的区域(高寒草甸和高寒草甸草原),而在相对较干旱的西部地区(高寒荒漠草原和高寒草原)没有明显的效果。这是由于虽然围栏封育对所有类型的高寒草地地上生物量均有显著的促进作用,但对物种丰富度的提升主要发生在高原东部较湿润的区域(图1)。由于物种多样性与丰富度对生态系统多功能性的显著促进作用,物种丰富度未发生显著改变的区域生态系统多功能性也不会发生显著提升。
4 结论
本研究表明,在藏北高原随降雨量的减少,植物的物种多样性、物种丰富度和生态系统多功能性逐渐降低,土壤养分等理化性质也逐渐降低,而土壤pH值和容重逐渐增加。12年围栏封育对高寒草地地上生物量有显著的促进作用,但对物种丰富度的提升主要发生在高原东部较湿润的区域。在高寒草甸中,12年的围栏封育对土壤碳、磷均显著提升,而对土壤pH值有降低作用;但在其他三种类型草地中,围栏封育对于土壤养分含量没有显著影响,而对土壤pH值有提高作用。12年的围栏封育对于生态系统多功能性的提升主要集中在降雨较多的区域,高寒草甸和高寒草甸草原生态系统多功能性分别提升91.0%和69.9%,而在干旱的西部地区(高寒荒漠草原和高寒草原)没有显著的效果。此外,高寒草地生态系统多功能性与物种多样性和丰富度(除Pielou均匀度指数外)均呈显著的正相关,表明维持较高的生态系统多功能性则需要更高的物种多样性和丰富度,这也解释了为什么围栏封育对生态系统多功能性的提升主要在高原东部较湿润的区域。然而,本研究在每种草地类型选取了一个样点,未来的研究会在每种类型高寒草地中增加实验样点来增加结果的可靠程度。
参考文献
[1]"""""" 杨君辉,程方方. 西藏全面加强草原生态保护建设综述[J]. 畜牧兽医科学,2018(1):59
[2]"""""" 张江,袁旻舒,张婧,等. 近30年来青藏高原高寒草地NDVI动态变化对自然及人为因子的响应[J]. 生态学报,2020,40(18):6269-6281
[3]"""""" 孙鸿烈,郑度,姚檀栋,等. 青藏高原国家生态安全屏障保护与建设[J]. 地理学报,2012,67(1):3-12
[4]"""""" 崔庆虎,蒋志刚,刘季科,等. 青藏高原草地退化原因述评[J]. 草业科学,2007,24(5):20-26
[5]"""""" WANG Y F,LV W W,XUE K,et al. Grassland changes and adaptive management on the Qinghai-Tibetan Plateau[J]. Nature Reviews Earth amp; Environment,2022(3):668-683
[6]"""""" 张骞,马丽,张中华,等. 青藏高寒区退化草地生态恢复:退化现状、恢复措施、效应与展望[J]. 生态学报,2019,39(20):7441-7451
[7]"""""" 范毅,加马力丁·吾拉扎汗,郭继敏. 围栏封育对草原植被恢复的作用[J]. 农村科学实验,2021(6):22-23
[8]"""""" 汪海霞,吴彤,禄树晖. 我国围栏封育的研究进展[J]. 黑龙江畜牧兽医,2016(9):89-92
[9]"""""" 程雨婷. 围栏封育后我国草地植被与土壤恢复的Meta分析研究[D]. 上海:华东师范大学,2020:19-39
[10]"""" 樊丹丹,孔维栋. 围栏对青藏高原不同类型草地土壤原核微生物多样性的影响[J]. 生态学报,2024,44(2):651-661
[11]"""" 左万庆,王玉辉,王风玉,等. 围栏封育措施对退化羊草草原植物群落特征影响研究[J]. 草业学报,2009,18(3):12-19
[12]"""" GALSTON A. Fundamentals of Ecology[J]. Yale Journal of Biology amp; Medicine,1972,45: 605-605
[13]"""" 冯剑丰,李宇,朱琳. 生态系统功能与生态系统服务的概念辨析[J]. 生态环境学报,2009,18(4):1599-1603
[14]"""" CARDINALE B J,DUFFY J E,GONZALEZ A,et al. Biodiversity loss and its impact on humanity[J]. Nature,2012,486(7401):59-67
[15]"""" TILMAN D,ISBELL F,COWLES J M. Biodiversity and ecosystem functioning[J]. Annual Review of Ecology, Evolution, and Systematics,2014(45):471-493
[16]"""" VAN DER PLAS F. Biodiversity and ecosystem functioning in naturally assembled communities[J]. Biological reviews of the Cambridge Philosophical Society,2019,94(4):1220-1245
[17]"""" HAZARD C,KRUITBOS L,DAVIDSON H,et al. Contrasting effects of intra- and interspecific identity and richness of ectomycorrhizal fungi on host plants, nutrient retention and multifunctionality[J]. The New phytologist,2017,213(2):852-863
[18]"""" 张全国,张大勇. 生物多样性与生态系统功能:最新的进展与动向[J]. 生物多样性, 2003, 11(5):351-363
[19]"""" 张全国,张大勇. 生物多样性与生态系统功能:进展与争论[J]. 生物多样性,2002,10(1):49-60
[20]"""" 江小雷,岳静,张卫国,等. 生物多样性,生态系统功能与时空尺度[J]. 草业学报,2010,19(1):219-225
[21]"""" GAMFELDT L,HILLEBRAND H,JONSSON P R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning[J]. Ecology,2008,89(5):1223-1231
[22]"""" 徐炜,马志远,井新,等. 生物多样性与生态系统多功能性:进展与展望[J]. 生物多样性,2016,24(1):55-71
[23]"""" 王凯,王聪,冯晓明,等. 生物多样性与生态系统多功能性的关系研究进展[J]. 生态学报,2022,42(1):11-23
[24]"""" HECTOR A,BAGCHI R. Biodiversity and ecosystem multifunctionality[J]. Nature,2007,448:188-190
[25]"""" MANNING P,VAN DER PLAS F,SOLIVERES S,et al. Redefining ecosystem multifunctionality[J]. Nature ecology evolution,2018,2(3):427-436
[26]"""" JING X,PRAGER CM,CLASSEN AT,et al. Variation in the methods leads to variation in the interpretation of biodiversity-ecosystem multifunctionality relationships[J]. Journal of Plant Ecology,2020,13(4):431-441
[27]"""" GARLAND G,BANERJEE S,EDLINGER A,et al. A closer look at the functions behind ecosystem multifunctionality: A review[J]. Journal of Ecology,2020,109(2):600-613
[28]"""" HOOPER D U,ADAIR E C,CARDINALE B J,et al. A global synthesis reveals biodiversity loss as a major driver of ecosystem change[J]. Nature,2012,486(7401):105-108
[29]"""" 徐炜,井新,马志远,等. 生态系统多功能性的测度方法[J]. 生物多样性,2016,24(1):72-84
[30]"""" BYRNES J E K,GAMFELDT L,ISBELL F,et al. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions[J]. Methods in Ecology and Evolution,2014,5(2):111-124
[31]"""" 李静鹏,郑志荣,赵念席,等. 刈割、围封、放牧三种利用方式下草原生态系统的多功能性与植物物种多样性之间的关系[J]. 植物生态学报,2016,40(8):735-747
[32]"""" 武建双,李晓佳,沈振西,等. 藏北高寒草地样带物种多样性沿降水梯度的分布格局[J]. 草业学报,2012, 21(3):17-25
[33]"""" 熊定鹏,赵广帅,武建双,等. 羌塘高寒草地物种多样性与生态系统多功能关系格局[J]. 生态学报,2016,36(11):3362-3371
[34]"""" ZHU J T,ZHANG Y J,WU J S,et al. Herbivore exclusion stabilizes alpine grassland biomass production across spatial scales[J]. Global Change Biology,2024,30(1):e17155
[35]"""" 周宸宇,杨晓渊,邵新庆,等. 不同退化程度高寒草甸植物物种多样性与生态系统多功能性关系[J]. 草地学报,2022,30(12):3410-3422
[36]"""" 胡亚. 水氮变化对荒漠草原植物群落多样性与生态系统多功能性关系的影响[D]. 北京:中国科学院大学,2021:64-81
[37]"""" HU Y,LI X Y,GUO A X,et al. Species diversity is a strong predictor of ecosystem multifunctionality under altered precipitation in desert steppes[J]. Ecological Indicators,2022(137):108762
[38]"""" FRY E L,SAVAGE J,HALL A L,et al. Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland[J]. Ecology, 2018, 99(10):2260-2271
[39]"""" YAN Y Z,ZHANG Q,BUYANTUEV A,et al. Plant functional β diversity is an important mediator of effects of aridity on soil multifunctionality[J]. Science of The Total Environment,2020(726):138529
[40]"""" 杨兆平,欧阳华,宋明华,等. 青藏高原多年冻土区高寒植被物种多样性和地上生物量[J]. 生态学杂志,2010,29(4):617-623
[41]"""" CHYTRY M,DANIHELKA J,ERMAKOV N,et al. Plant species richness in continental southern Siberia: effects of pH and climate in the context of the species pool hypothesis[J]. Global ecology and biogeography,2007,16(5):668-678
[42]"""" 郭志霞,刘任涛,冯永宏,等. 不同降水对荒漠灌丛土壤理化性质和地表植被分布的影响[J]. 水土保持通报,2021,41(1):56-65
[43]"""" 韦昌林,李毅,单立山,等. 降水变化对典型荒漠植物凋落物分解的影响[J]. 草地学报,2022,30(5):1280-1289
[44]"""" 刘晨,康慧,孙海莲,等. 模拟降水对短花针茅荒漠草原土壤养分的影响[J]. 草原与草业,2021,33(4):22-26
[45]"""" 贺金生,刘志鹏,姚拓,等. 青藏高原退化草地恢复的制约因子及修复技术[J]. 科技导报,2020,38(17):66-80
[46]"""" 姚喜喜,才华,李长慧. 封育和放牧对高寒草甸植被群落特征和土壤特性的影响[J]. 草地学报,2021, 29(S1):128-136
[47]"""" XIONG D P,SHI P L,ZHANG X Z,et al. Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China A meta-analysis[J]. Ecological engineering,2016(94):647-655
[48]"""" 聂莹莹,杜广明,王国庆,等. 围栏封育对呼伦贝尔草甸草原群落物种多样性的影响[J]. 中国草地学报,2016,38(6):106-110
[49]"""" 贾晓妮,程积民,万惠娥. 云雾山本氏针茅草地群落恢复演替过程中的物种多样性变化动态[J]. 草业学报,2008,17(4):12-18
[50]"""" 任毅华,周尧治,井向前,等. 围栏封育对西藏退化高寒草甸物种多样性及生产力的影响[J]. 贵州农业科学,2015,43(10):166-169
[51]"""" 黎松松,王宏洋,林华,等. 围栏封育对喀尔里克高寒草甸优势种数量特征和物种多样性的影响[J]. 草食家畜,2023(3):26-30
[52]"""" 周天阳,高景,王金牛,等. 基于群落结构及土壤理化性质对围封7年青藏高原东南缘高山草地的综合评价[J]. 草业学报,2018,27(12):1-11
[53]"""" BAI G,BAO Y Y,DU G X,et al. Arbuscular mycorrhizal fungi associated with vegetation and soil parameters under rest grazing management in a desert steppe ecosystem[J]. Mycorrhiza,2013,23(4):289-301
[54]"""" 杜陈军. 围封对退化高寒草甸生态系统CO2交换的影响[D]. 北京:中国科学院大学,2019:32-38
[55]"""" 康宝天,侯扶江,SAMAN B. 祁连山高寒草甸和荒漠草原土壤细菌群落的结构特征[J]. 草业科学,2020,37(1):10-19
[56]"""" 赵帅,张静妮,赖欣,等. 放牧与围封对呼伦贝尔针茅草原土壤酶活性及理化性质的影响[J]. 中国草地学报,2011,33(1):71-76
[57]"""" 刘建康,张克斌,冯湘,等. 封育方式对荒漠草原群落特征及土壤理化性质的影响[J]. 生态环境学报,2021,30(3):445-452
[58]"""" YU L F,SUN W J,ZHANG H Y,et al. Grazing exclusion jeopardizes plant biodiversity effect but enhances dryness effect on multifunctionality in arid grasslands[J]. Agriculture Ecosystems amp; Environment,2024,363:108883
[59]"""" ZHENG J H,ZHANG B,ZHANG F,et al. Effects of fencing on near-term ecosystem multifunctionality in a typical steppe in Inner Mongolia[J]. Agriculture Ecosystems amp; Environment,2023(342):108238
[60]"""" 张文馨,范小莉,王强,等. 黄河三角洲植物多样性与生态系统多功能性间的关系[J]. 山东大学学报(理学版),2020,55(1):110-116
[61]"""" VAN DER PLAS F,MANNING P,ALLAN E,et al. Jack-of-all-trades effects drive biodiversity-ecosystem multifunctionality relationships in European forests[J]. Nature communications,2016,7(1):1-11
(责任编辑" 付宸)
