榧树属植物资源研究进展
2024-12-27马长乐杨建欣桂晴龚买玉周龙飞刘佳


关键词:榧树属;植物资源;分类;演化;研究进展
榧树属Torreya Arn. 也称榧属,隶属红豆杉科Taxaceae,为古老珍稀的孑遗植物,均分布于北半球。榧树属植物共有10 个种(含2 变种),北美产2 种(‘佛罗里达榧’Torreya taxifolia 和‘加州榧’Torreya californica), 日本产1 种(‘ 日本榧’Torreya nucifera),其余7 种在中国自然分布(‘榧树’Torreya grandis、‘巴山榧’Torreyafargesii、‘长叶榧’Torreya jackii、‘云南榧’Torreyayunnanensis、‘四川榧’Torreya parvifolia、‘九龙山榧’Torreya grandis var. Jiulongshanensis、‘大盘山榧’Torreya dapanshanica)[1-6]。根据2021 年发布的《国家重点保护野生植物名录》(除2022年发表的新物种‘大盘山榧’外),我国榧树属全部野生种(包括变种)均为国家二级重点保护野生植物。相关资料显示,榧树属植物在中侏罗纪至第三纪上新世分布于北美、欧洲及亚洲,第四纪冰期后,欧洲范围内的分布区已然消失,现间断分布于东亚、北美东南部和西部[2]。由于生境破碎化程度日趋严重,榧树属的分布区范围愈加狭窄,种群衰退,个体数量逐渐减少。‘巴山榧’‘长叶榧’‘云南榧’和‘九龙山榧’等物种已上榜《中国物种红色名录》[7]。
榧树属植物枝叶繁密,树冠开展而整齐,树形优美,并能长期保持稳定;花、果、叶、姿等均具有较高观赏价值,适合园林造景;种子可供食用和药用;木材结构紧密,纹理细致,耐水湿,抗腐性强,是优良的建筑及家具用材,隋唐时期就作为珍贵树木在洛阳地区种植[8]。综合而言,榧树属植物是兼具药用、食用、观赏为一体的优良树种。本文从资源现状与地理分布、起源与系统演化、分类、群落结构特征、繁殖与遗传、生化成分等方面全面综述榧树属10 个物种的研究现状并提出研究展望,为更好地保护和利用榧树属植物资源提供参考。
1 种质资源与地理分布
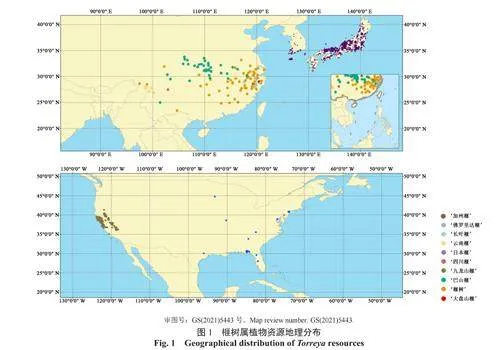
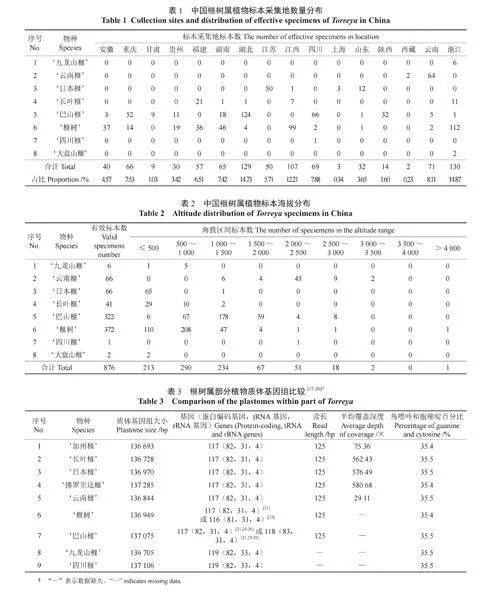
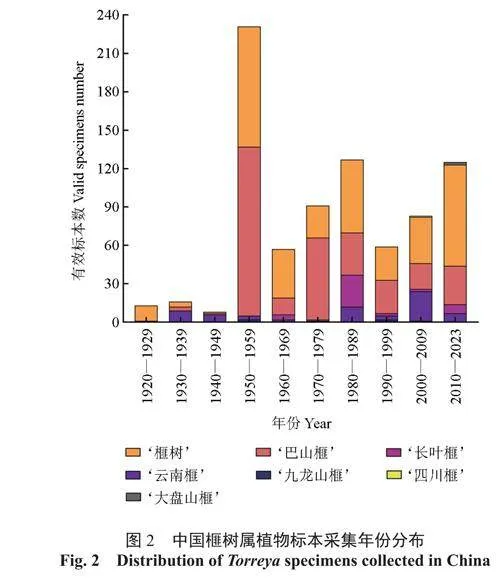
首先,采用R 语言“rgbif”程序包从世界生物多样性信息机构(https://www.gbif.org/,GBIF)检索获取榧树属植物分布数据,并从现有中英文文献、植物智(https://www.iplant.cn/)和全球数字化植物标本数据库(https://plants.jstor.org)等收集榧树属植物分布数据作为补充。对于只有地理位置但没有具体经纬度信息的分布数据,使用Excel插件DataMap For Excel(version 6.9.2),将地址转换为经纬度。然后,对相关信息进行核对和整理,删除重复及信息不完整的数据。最后,根据收集到的信息绘制成榧树属植物资源地理分布图(图1)中国榧树属植物标本采集年份分布图(图2)、采集地及数量分布表(表1)、海拔分布表(表2)。
从榧树属植物资源地理分布情况(图1)可知,榧树属植物具有分布区狭窄及不连续的共性,斑块化和岛屿化现象较为明显。‘榧树’和‘巴山榧’自然分布区的地理跨度相对较大[9],其他种均零星分布。‘榧树’片段化分布于浙江和福建等省区[10]。‘巴山榧’间断分布于湖北等省区。‘云南榧’仅分布于滇西北横断山区[11]。‘九龙山榧’主要分布于浙江九龙山[12]。‘四川榧’仅分布于四川布拖县临近金沙江位置[4]。‘长叶榧’仅分布于浙江、福建、江西、湖南和湖北[13-14]。‘日本榧’主要分布于日本和朝鲜半岛,在我国上海、青岛、南京等城市亦有引种栽培。‘加州榧’和‘佛罗里达榧’尚无在我国引种栽培的记载。随着全球气候变化和人类活动的影响,各物种的分布位置可能会发生迁移,消失或濒临消失,部分物种可能会逐步分布于海拔和温湿度更加适宜的区域[11,15]。
从中国榧树属植物标本采集年份分布图(图2)可知,自1920 年开始,榧树属植物标本就开始被逐步采集获取,其中1950 年至1959 年期间采集量最大,主要为‘榧树’和‘巴山榧’2 个物种。1960 年至今,伴随着植物学研究领域的兴起,榧树属标本采集数量保持稳定且呈现出逐步增加的状态,其中‘榧树’和‘巴山榧’仍然处于标本采集数量的前列。从1970 年开始,其他物种的标本数量逐渐增加,‘长叶榧’‘云南榧’‘四川榧’‘九龙山榧’‘大盘山榧’的标本逐步被采集获取,但由于后3 种植物的分布区域较为狭窄,采集到的标本数量十分有限。从标本采集地来看(表1),在浙江、江西、湖北、云南、四川等省采集到的标本数量较多,表明这些省份是榧树属植物的集中分布区域。从标本采集地的海拔范围来看(表2),在海拔1 500 m 以下获取的标本数量较多,表明榧树属植物主要分布于该海拔区间内。
2 起源、系统演化与分类
2.1 基因组结构
目前用于物种系统发育与演化历史研究的基因组学方法有叶绿体基因组、线粒体基因组和全基因组学等。上述方法在许多被子植物的研究中得到广泛应用并取得大量成果。虽然‘火炬松’Pinustaeda、‘苏铁’Cycas revoluta 全基因组测序和解析工作已经完成,但由于裸子植物具有基因组巨大、重复序列占比高等特征,导致裸子植物基因组学研究相对滞后,涉及到裸子植物类群复杂基因组结构的诸多科学问题仍有待深入探讨[16]。
相关研究表明(表3),榧树属植物的质体基因组大小处于136 693 ~ 137 412 bp,测序质体中GC含量相似(35.4% ~ 35.5%)[17-26]。‘加州榧’‘佛罗里达榧’‘长叶榧’‘云南榧’‘榧树’‘日本榧’‘巴山榧’等7 个物种包含117 个特有基因,其中有82 个蛋白编码基因、31 个tRNA 基因和4个rRNA 基因[19-20],但也有研究认为‘榧树’和‘巴山榧’的蛋白编码基因数量与其他物种不同[21-26]。‘九龙山榧’和‘四川榧’包含119 个特有基因,其中有82 个蛋白编码基因、33 个tRNA 基因和4个rRNA 基因[22-23]。尚未查到关于‘大盘山榧’质体基因组的相关研究。关于‘日本榧’,有研究发现其蛋白编码基因与tRNA 基因数量与其他物种不同[22]。有研究证实,榧树属植物的3 个tRNA基因(trnL-CAU、trnQ-UUG 和trnN-GUU) 具有双拷贝,且质体缺失典型的反向重复区(invertedrepeat,IR),导致长单拷贝区和短单拷贝区边界不明[19]。截至目前,这一特性已经在除‘九龙山榧’‘四川榧’和‘大盘山榧’之外的7 个物种中得到证实。但根据现有研究可以推断,包括‘九龙山榧’‘四川榧’和‘大盘山榧’在内的该属所有物种的质体基因组均满足上述规律。榧树属因失去1 个IR 区,导致其同源重组能力下降,使得质体基因组的多样性降低。但关于丢失机制及其进化历程的研究目前尚无定论。目前,关于裸子植物基因组的研究大部分仅涉及叶绿体(质体)基因组,关于榧树属的基因组研究也仅见于绿体(质体)基因组[19-26]。虽然榧树属大多数物种的质体基因组信息已得到验证,但仍停留在基本特征分析,基因组结构与系统发育的内在联系缺乏深入解析,核基因或全基因组的相关研究有待今后进一步开展。随着分子生物学和测序技术的发展。该属植物全基因组测序工作已逐步开展。‘香榧’基因组测序工作揭示了裸子植物特有成分金松酸的生物合成机制[24],是裸子植物基因组研究工作的重要补充和突破性成果。
2.2 起源与系统演化
目前,榧树属各物种的起源及进化关系脉络尚未完全理清。由于研究方法和数据来源不同或物种错误识别等原因,各物种在系统发育中的地位存在多种不同意见甚至矛盾观点[19,25-29]。从物种间演化关系的角度认为,榧树属与穗花杉属Amentotaxus 的分化时间估计在54.74 Ma[21]。‘长叶榧’是最早分化出来的物种,与属内其他物种产生分化的时间约在6.94 Ma,但关于其亲缘关系的推断存有不同见解[21,28]。此外,‘长叶榧’的质体与核系统发育位置不一致,可能是由于基因渐渗所引起的[29]。通过质体进化和系统发育关系研究确定了‘长叶榧’的系统发育位置,并认为‘巴山榧’与‘日本榧’,‘榧树’与‘加州榧’互为姐妹关系,‘云南榧’‘巴山榧’和‘日本榧’为单系进化分支[21]。基于叶绿体数据,认为‘榧树’‘巴山榧’和‘云南榧’互为姐妹关系[28];‘日本榧’和‘榧树’,‘四川榧’‘巴山榧’和‘云南榧’分别位于一个二级进化分支(100% 支持,它们共同构成同一个一级进化分支);‘长叶榧’和‘九龙山榧’则位于另一个一级进化分支(100%支持),但这两个一级进化分支间仅有94% 支持[30-31]。基于内转录间隔区序列的数据,对于‘榧树’与‘长叶榧’的系统地位持有不同观点。有研究认为‘榧树’与除‘长叶榧’之外的其他物种互为姐妹关系[28];另有研究则认为‘榧树’和‘长叶榧’互为姐妹关系[26],‘日本榧’‘巴山榧’和‘云南榧’互为姐妹关系[27,32],‘九龙山榧’是‘榧树’和‘长叶榧’的自然杂交种[19,29]。关于‘日本榧’和‘云南榧’在系统发育中的地位也一直没有定论[26,32]。
从生物地理的研究角度认为,榧树属植物的多样化与新近纪(中新世和上新世)的气候变化有关。目前的生物地理格局可能是受晚第三纪及以后频繁气候振荡和环境变化的影响[27]。系统发育祖先性状重构软件RASP 4 分析表明,在榧树属内部共发生过4 次扩散事件和3 次地理隔离事件。中国和北美东部最有可能是现存榧树属分布的祖先地区。中新世至上新世的重要扩散事件和随后第四纪的地理隔离形成了目前的生物地理分布格局。部分物种可能在“白令陆桥”消失之前成功完成扩散和迁徙。第四纪的气候变化为海洋两侧的物种创造了屏障,促进榧树属异地物种的形成。上新世和更新世的气候变化导致‘加州榧’和‘佛罗里达榧’的分布范围逐渐收缩,出现地理隔离,形成了与其它亚洲物种隔离的独立分支[33-34]。
整合系统基因组、形态学和化石证据等数据,认为榧树属在侏罗纪早期(186.4 Ma)[26] 或在54.74 Ma[21] 与穗花杉属从一个共同的祖先分离出来,在侏罗纪时期(201.3 ~ 145.0 Ma)进一步分化形成多个谱系,形成北美和东亚两个进化分支。在北美进化分支中,‘加州榧’和‘佛罗里达榧’的分化时间可追溯到渐新世时期(26.4 Ma)。在东亚进化分支中,‘巴山榧’和‘云南榧’在渐新世(37.1 Ma)与‘榧树’分离,在中新世中期(13.7 Ma)两者随之发生分化,形成了相互分离的单系群;‘长叶榧’在始新世至渐新世早期(34.4 Ma)从‘榧树’-‘日本榧’支系分化而来;‘榧树’则在中新世早期(25.8 Ma)从‘日本榧’分化出来[26](图3)。
2.3 分类
对榧树属的分类研究始于19 世纪30 年代。该属最初由英国植物学家George Arnort 所创立,并确定原产北美的‘佛罗里达榧’为模式种[1]。其后,诸多植物学先驱对榧树属物种进行鉴定和分类[2,35-36]。1846 年,‘日本榧’被正式归入榧树属。1927 年,胡先骕教授率先在国内开展榧树属分类研究,并发表4 个变种和2 个变型[35]。他在编撰《中国植物志》和《中国树木志》时,将‘长叶榧’‘榧树’‘巴山榧’‘日本榧’和‘云南榧’独立出来[1,37]。其后,Silba 将‘巴山榧’改为‘榧树’的变种,并将‘云南榧’与之合并[38-39]。而在《Floraof China》,‘云南榧’被视为‘巴山榧’的变种[3]。近些年,‘四川榧’和‘大盘山榧’2 个新物种先后被发现[4,6]。《中国生物物种名录》[40] 全面收录了榧树属全部物种,将‘四川榧’作为‘巴山榧’的亚种(表4)。
胡先骕教授研究认为,根据榧树属种子胚乳周围向内褶皱的程度(微皱和深皱),可将其分为榧树组(Sect. Torreya /Sect. Nuciferae)和皱乳榧树组(Sect. Ruminatae);前者包括‘加州榧’‘榧树’2 种,后者包括‘佛罗里达榧’‘巴山榧’‘长叶榧’和‘云南榧’4 种[2]。根据形态特征,本文认为此后发表的‘四川榧’应归入皱乳榧树组,‘九龙山榧’‘日本榧’和‘大盘山榧’则应归入榧树组。然而,近年来部分学者的系统发育研究结果并不支持这一传统的分组方法[25,27,29]。
在《中国植物志》和《Flora of China》的分类中[1,3],红豆杉科包括红豆杉属Taxus、白豆杉属Pseudotaxus、穗花杉属和榧树属4 个属,但其系统位置和科属关系等方面仍存在较大争议。核心问题在于红豆杉科的范围、属间关系及其与三尖杉科Cephalotaxaceae 的关系。关于红豆杉科和三尖杉科的分类也有诸多相互矛盾的观点,两者之间的明确划分还没有完全解决,长期以来争论的焦点是两者是否应该合并。主要观点有以下几点:1)认为穗花杉属与三尖杉属Cephalotaxus 亲缘关系较近;2)胚胎学研究结果建议将红豆杉科分为穗花杉族(含穗花杉属、南紫杉属Austrotaxus)、红豆杉族(含红豆杉属和白豆杉属)及榧树族(含榧树属)3 个族;3)根据双黄酮成分在红豆杉科各属的分布情况,将穗花杉属和白豆杉属归为红豆杉科还存在疑问;4)根据花粉形态及花粉外壁的超微结构,则建议将红豆杉科分为红豆杉族(含红豆杉属和榧树属)和白豆杉族(含白豆杉属)[41-42]。对于穗花杉属和榧树属的归属关系也未得到统一结论。虽然主要观点认为两者属于红豆杉科,但其与三尖杉科在形态学、解剖学、胚胎学和系统发育学上都有相似之处,因此有学者建议将三尖杉科并入红豆杉科[43],或将穗花杉属和榧树属归入三尖杉科[44]。也有观点认为穗花杉属和榧树属在叶绿体matk 和核rDNA ITS 区上与红豆杉科的亲缘关系较近,与三尖杉科的亲缘关系较远[45-46]。综上,目前还难以对以上分歧作出清晰的解释。
3 群体结构与遗传多样性
植物遗传多样性主要受植物群体结构、繁育系统、基因流、遗传漂变和自然选择等多种因素的影响[47],其分布格局和遗传结构是气候变化、地质事件和人类活动共同作用的结果[48]。全面掌握榧树属植物的种群结构特征、种内遗传变异情况、时空分布格局及其与环境条件的相互关系,能够为分类、进化、栽培、繁殖及遗传改良等方面的研究工作提供参考。对‘榧树’‘云南榧’‘巴山榧’和‘佛罗里达榧’的种群生态学研究表明,榧树属植物虽然有着较强的资源利用能力,但生态位宽度较小[49],年龄结构不合理,幼苗极度缺乏,中龄个体偏少,自然更新能力后劲不足,并持续受到人类活动的干扰和各类病害的侵扰,生存和繁衍受到不同程度的威胁[50-53]。目前相关研究较少,只有关于少数物种的零星报道,难以全面揭示该属植物的群落结构特征和空间分布格局。
研究发现,植物核基因组保持较高遗传多样性的原因可能是在进化过程中保留了广泛的遗传基础,也可能是后期自然选择产生了丰富的遗传变异[54-56]。‘榧树’为雌雄异株的二倍体裸子植物,与‘银杉’Cathaya argyrophylla 等其他裸子植物一样,叶绿体基因组为父系遗传;若居群中有相等的性比,则叶绿体基因组有效居群大小只有核基因组的1/4,因此叶绿体基因组对等位基因的丧失相对于核基因组更为敏感[54-56]。这一特殊的遗传模式导致其更容易流失突变导致的基因组重复区域,进而影响到其他基因组水平的演化。因此,叶绿体基因组的遗传多样性是瓶颈效应、奠基者效应和遗传漂变的指示器[55]。研究表明,‘榧树’‘巴山榧’‘长叶榧’居群均保持着较丰富的遗传多样性,遗传差异主要分布在居群内[57-58],其原因可能与进化历史有关。其中,‘长叶榧’群体由于受第四纪冰川的影响,其数量急剧下降,自然分布范围缩小,大量个体灭绝,幸存个体在一些冰期避难所中保留下来[59],其祖先广泛的遗传基础在不同的避难所中得以保留;但‘香榧’群体水平的遗传多样性较低,遗传衰退的主要原因有选择作用、遗传漂变以及自交现象等[58]。此外,由于雄性‘榧树’表型性状在居群内具有更为丰富的遗传多样性,因此在后期遗传改良中应重视群体内优良单株的选择,而球花大小、花粉生活力和花粉得率是‘榧树’雄株进行优株初选的重要指标[60]。总体而言,目前关于榧树属遗传多样性的研究仅见于少数物种,属内全部物种遗传多样性特征是否存在相同或相似的规律还有待进一步研究证实。当前,针对物种的遗传多样性特点,宜采取不同措施进行种质资源保护:就地保护时应当对尽可能多的群体实施保护;迁地保护时应当从所有群体采集样本,尽量保存遗传资源,避免在群体间实施种质迁移[57-58]。
4 栽培与繁殖
榧树属种群分布区域狭窄、分布密度小、开花结实率低、种子后熟、天然下种后出苗慢、成苗率低、幼苗生长较慢、更新困难、生殖周期冗长、生殖过程复杂、雌雄生殖和发育系统差异化[5]的特点是导致榧树属逐渐趋于濒危的主要原因。榧树属植物的种子一般较难获取,除栽培品种‘香榧’T. grandis ‘Merrillii’外,其他物种实生后代均难以健壮生长,优良性状不能稳定传递[61-62]。然而,美国农业部的一项调查数据显示,‘加州榧’和‘佛罗里达榧’在火灾后能很好地萌发[63],说明其具备良好的自然更新能力,对繁殖栽培和生境恢复具有一定启示。
关于榧树属植物栽培繁殖的研究主要聚焦于扦插繁殖、嫁接繁殖和组织培养等方面。但大多数研究都是以‘香榧’为材料,属内其他珍稀濒危物种的繁育尚未见系统的研究。榧树属的嫁接是当前相对成熟的技术措施,但关于砧木和接穗的年龄的影响有不同观点:1)砧木年龄越小活力越强,越易嫁接成功[61];2)砧木年龄对嫁接苗成活的影响不明显,但对成活后接穗的生长具有显著影响[62]。嫁接季节和嫁接方法对‘香榧’嫁接成活率的影响目前尚无定论[64]。扦插繁殖和组织培养仍处于实践探索阶段。最新研究表明,夏季扦插的‘香榧’雄株比雌株具有更高的扦插成活率[65],但多项技术难题仍未得到有效解决。
5 植物化学成分
榧树属植物中含有丰富的挥发油(精油)、萜类、黄酮类、抗疟活性物质和植物毒素等活性成分,具有良好的药用价值、营养价值和经济价值。当前对榧树属生化成分的研究报道较多,研究对象主要为人工培育品种‘香榧’。也有关于‘云南榧’[66-68]、‘长叶榧’[69]、‘日本榧’[70-71] 和‘佛罗里达榧’[61] 的少量研究。
5.1 脂肪酸及氨基酸
脂肪酸包括饱和脂肪酸和不饱和脂肪酸,主要来源于榧树属植物的种子和假种皮[72-73]。不饱和脂肪酸(主要为亚油酸、油酸、金松酸)具有一定的抗氧化活性,对羟基自由基的清除能力强于超氧阴离子自由基,对心脑血管疾病的防治具有较好的协同增效作用[72]。研究发现,‘香榧’种子含有32 种不饱和脂肪酸、12 种饱和脂肪酸[73];‘云南榧’种子鉴定出14 种脂肪酸,其中不饱和脂肪酸12 种[71]。关于榧树属植物种子的研究发现其含有17 种或19 种常见氨基酸,总氨基酸及氨基酸组分含量不同[74]。品种间总氨基酸含量高低顺序与必需氨基酸、味觉类氨基酸含量的高低顺序一致,其中天门冬氨酸、谷氨酸和精氨酸的含量居于前列,人体必需氨基酸(总数为7 种)的占比优于WHO/FAO 理想营养模式谱[75]。研究证实,在不同发育阶段‘香榧’果仁中氨基酸含量差异很大。目前,氨基酸生物合成的分子基础和调控机制仍不清楚[76],仅有关于金松酸这一榧籽油的特征性脂肪酸的研究报道[24,77]。
5.2 精油及萜类物质
在榧树属植物茎、叶、种子和假种皮中均含有丰富的精油,且其组分复杂;最主要的活性成分是萜类,具有良好的抗氧化活性,如α - 蒎烯、β - 蒎烯、柠檬烯、苎烯、香叶烯等;其中叶片和假种皮中α - 蒎烯和柠檬烯含量最高[78-79]。在‘香榧’假种皮中检测出挥发油47 种(萜烯类21 种、萜醇类15 种、萜烯酯类3 种、其他化合物8 种)[80]。柠檬烯和蒎烯为‘长叶榧’茎部的主要成分;其叶片中还具有另外13 种化合物,其中石竹烯、罗勒烯、β - 月桂烯、反式α - 蒎烯、顺式α - 蒎烯含量最高[69,81]。此外,榧树属精油中还存在榧树醇、榧树醛、香榧酯和紫杉醇等重要化合物,尤其是紫杉醇的分离和定量一度成为研究热点[82]。但目前关于‘香榧’假种皮中是否含有紫杉醇及其含量多少以及是否具备合成紫杉醇的能力等相关结论仍然有较大争议[82]。
5.3 黄酮类化合物
黄酮类化合物主要存在于绿色植物的花、树皮、叶片、果实和根部,具有清除自由基、抗氧化、抗肿瘤、抗病毒及清热解毒等多种功效[83]。研究发现,榧树属植物中黄酮类化合物主要有榧黄素(双黄酮类化合物)、托亚埃Ⅰ号、托亚埃Ⅱ号和四甲基阿曼托黄素等[84]。其中,榧黄素为红豆杉科植物的特征成分,已在‘长叶榧’‘日本榧’‘云南榧’和‘榧树’中分离得到;托亚埃Ⅰ号和托亚埃Ⅱ号(取代黄酮类化合物)已从‘香榧’枝叶中分离得到。此外,转录组测序和代谢组学分析表明‘香榧’果仁含有124 种黄酮类化合物,其中9 种黄酮类化合物与抗氧化活性高度相关,来自MYB、bHLH 和bZIP 家族的几种转录因子可能调节其生物合成[85]。总体而言,黄酮类化合物的含量与树龄有关,并受到脱落酸和赤霉素的调控[86],有着低含量、低溶解性和低生物利用度等特征,因此如何发掘其生物利用度是进一步发挥其食用和药用价值的基础。
5.4 其他物质
除上述主要成分外,榧树属植物中还存在木脂素、维生素、糖类和矿物质等[87]。现已知的木脂素类仅有3 种,其中松脂素具有抗骨质疏松的作用[83]。‘香榧’的木脂素类成分主要分布于叶片和假种皮,其种子中含有烟酸、叶酸、维生素B2、维生素D3 及19 种矿质元素;‘云南榧’的树皮、木质部和叶片中均含有人体必需矿质元素。但相关物质的具体种类和含量在不同物种和部位间有较大差异。不可忽视的是,榧树属植物化学成分中还存在一定量的微量重金属元素[68],在药用和食用价值开发时应加以重视。
6 研究展望
目前关于榧树属的相关研究主要集中在‘榧树’及其栽培品种,其余物种的研究较为匮乏。为了更好地保护和利用好榧树属植物资源,提出以下研究展望:
1)深入开展榧树属各物种的生物学及生态学特性研究,明确其种群濒危的生理和生态学机制,加强对种群及栖息地的就地保护,减缓或防止栖息地生境及生态系统的破坏。采用谱系地理学方法研究种群分布格局的形成机制及其演化过程,通过其空间分布格局来验证与之相关的地质事件,追溯和解释种群分化的历史原因,并将化石数据整合到系统发育和生物地理分析中,验证和解释其发育和起源问题。综合应用形态学、细胞学、地理学、分类学、遗传学等多学科的研究方法,整合属内全部物种,逐步澄清属内系统分类问题。
2)利用分子生物学和基因测序技术开展遗传多样性研究,探明榧树属自然群体的遗传结构、遗传多样性水平和群体间遗传分化程度,明确各物种在特定范围内的生存潜力和濒危程度。采用遗传连锁图谱构建技术,深入研究遗传机制,为高质量解析基因组和探讨不同物种的遗传变异奠定基础。充分整合基因组学、转录组学、蛋白组学和代谢组学等多组学技术手段,探讨遗传进化机制和濒危机理,推测冰期避难所及冰期后的迁移路线,进一步明确榧树属物种起源与系统演化关系,为种质资源保育提供科学依据。
3)榧树属植物目前除了其栽培品种‘香榧’在江浙地区和皖南山区得到规模化培育以外[88],大多数物种仍处于自然野生状态。建议今后加强在良种选育、园林应用等方面的实践与探索。同时,应在‘香榧’研究成果的基础上,采用转录组测序和代谢组学分析的方法,深入挖掘属内其他物种在食用和药用价值开发利用中的关键化学物质分子基础和调控机制,为该属植物潜在的经济价值开发提供更加科学的基础资料。
