‘热研2号’柱花草对不同邻近植物的识别与生物响应差异
2024-11-05李雪枫陈七双许超肖云霈罗瑛王坚





收稿日期:2024-02-25;修回日期:2024-04-16
基金项目:国家自然科学基金项目(32260344);海南省自然科学基金项目(324RC444、321RC471)资助
作者简介:李雪枫(1974-),女,汉族,宁夏隆德人,博士,主要从事草地生态、植物资源与环境研究,E-mail:xfli0922@163.com;*通信作者Author for correspondence,E-mail:wangjian901@163.com
摘要:为明确‘热研2号’柱花草(Stylosanthes guianensis‘Reyan No. 2’)对邻近植物的识别与生物响应对策,研究了其分别与自我、近亲、远亲和非亲4个亲缘关系组的10种植物伴生时的生长情况。结果表明:亲缘关系对‘热研2号’柱花草的株高和分枝数均值无显著影响,但近亲组伴生使其总生物量和地上生物量均值显著增加而根冠比均值显著减小(P<0.05),远亲和非亲组伴生使其根长、根冠比、生物量均值均显著降低而侧根数均值显著增加(P<0.05)。生长指标变化结合主成分分析表明:与‘热研21号’(S. guianensis ‘Reyan No. 21’)、‘热研24号’(S. guianensis ‘Reyan No. 24’)、‘热研5号’(S. guianensis ‘Reyan No. 5’)和鬼针草(Bidens pilosa)为邻株时,‘热研2号’柱花草的地上部生长显著增强,且程度依次减弱;与西卡柱花草(S. scabra)、细茎柱花草(S. gracilis)、蒺藜草(Cenchrus echinatus)、羽芒菊(Tridax procumbens)和大叶柱花草(S. grandifolia)为邻株时,其地上部和地下部生长均变差,且程度依次增强。说明‘热研2号’柱花草与近亲缘品种伴生时地上部生长相对较强,与远亲缘和非亲缘种伴生时地上、地下生长均减弱。
关键词:邻居识别;响应策略;侧根数;分枝数;生物量分配
中图分类号:S541.9 文献标识码:A 文章编号:1007-0435(2024)10-3225-08
Studies on the Discrepancy of Recognition and Biological Response of Stylosanthes guianensis ‘Reyan No.2’ to Different Neighbor Plants
LI Xue-feng, CHEN Qi-shuang, XU Chao, XIAO Yun-pei, LUO Ying, WANG Jian*
(College of Tropical Agriculture and Forestry, Hainan University/ Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Haikou, Hainan Province 570228, China)
Abstract:In order to clarify the recognition and biological response strategies of Stylosanthes guianensis ‘Reyan No. 2’ to different neighbor plants,this experiment was conducted to study the growth changes of S. guianensis ‘Reyan No.2’ when it was respectively grew adjacent to 10 plant species belonging to four genetic treatment groups:self,close relative,distant relative and non-relative. The results showed that the relative relationship had no significant effects on the mean plant height and branch number of S. guianensis ‘Reyan No.2’. Compared with the self-treatment group,the mean values of total biomass and aboveground biomass in the close-relative treatment group increased significantly,while the mean value of root-shoot ratio decreased significantly (P<0.05). The mean values of root length,root-shoot ratio and biomass of the distant relative treatment group and the non-relative treatment group significantly decreased (P<0.05),but the mean value of the number of lateral roots significantly increased (P<0.05). The changes of each growth index combined with the results of principal component analysis showed that when S. guianensis ‘Reyan No.2’ neighbored with S. guianensis ‘Reyan No.21’,S. guianensis ‘Reyan No. 24’,S. guianensis ‘Reyan No. 5’ and Bidens pilosa,respectively,the growth of the aboveground part significantly enhanced,and the degree of response weakened in order. When S. guianensis ‘Reyan No. 2’ neighbored with S. scabra,S. gracilis,Cenchrus echinatus,Tridax procumbens and S. grandifolia, respectively,the growth of aboveground and underground parts became worse,and the degree increased in order. These results indicated that the aboveground growth of S. guianensis ‘Reyan No. 2’ was relatively strong when its close relative was its neighbor,and the aboveground and underground growth was weak when its distant relatives and non-relatives were its neighbor.
Key words:Neighbor recognition;Response strategy;Number of lateral root;Number of branch;Biomass distribution
植物可以接收和识别不同环境中的物理、化学或生物信号,并根据这些信号作出相应的响应[1]。通过改变对自然资源竞争具有直接影响的形态和生物量分配等生物响应指标来适应周围微环境条件变化的能力是植物生态研究的重要内容之一[2-3]。由于植物本身不能像动物那样自由移动,相对较远的个体对于植株体彼此之间地上部遮阴、地下部吸收养分和水分等互作过程的影响较小,植物与它相邻生长的植物(邻近植物)之间的相互作用是决定小尺度空间结构的重要因素[4-5]。相邻生长的植物之间可能存在争夺土壤、光照、水分等自然生态因子而进行的生长策略调整[6],也可能存在邻近植物所释放次生代谢物的化感作用所导致的生长策略变化[7]。养分利用能力强、具有较强表型可塑性的植物必然在与邻近植物的相互干扰中占据主导地位[6,8],在多变的环境中获得生存优势[9]。
植物与邻近植物之间因相互识别、相互干扰而进行的生长策略调整是影响植物与邻近植物种间或种内关系的重要因素[7,10],特别是自然植物种群中,由于种子的随机传播,邻居之间的亲缘关系无法预测,亲缘识别可以使植物以较小的利他主义成本实施利于邻近亲属生长的竞争策略[11],主要通过形态、分配、叶和根空间构型特征影响彼此的竞争能力[12]。如菟丝子(Cuscuta chinensis)能识别寄主植物释放的挥发性物质从而朝向寄主植物番茄(Lycopersicon esculentum)所在的一侧生长而趋避非寄主植物小麦(Triticum aestivum)一侧,使番茄易于被其寄生受害而小麦难以被其危害[13];大豆(Glycine max)与种内品种伴生时的叶生物量无显著变化,与野大豆(Glycine soja)伴生时却显著增加[14];紫茎泽兰(Ageratina adenophora)与小藜(Ageratina adenophora)为邻时的生物量比其与自身种植株为邻时显著升高,而小藜的生物量却显著下降[15]。活性炭吸附拟南芥(Arabidopsis thaliana)根系分泌物的试验也证明即便邻近植物是自身品种,其分泌物也会影响根系形态,使拟南芥侧根数减少、侧根长增加[16]。这些研究结果说明植物最终的生长策略和生物响应结果取决于邻近植物的具体特性[9,17],也表明植物能够非加性地整合营养物质与邻近植物的信息[18],从而调整其生长、繁殖和防御策略,优化资源分配和利用以提高个体适合度[7]。因此,研究植物对其邻近植物的识别与响应效应对构建可持续发展的农林生态系统具有重要意义。
柱花草(Stylosanthes spp.)是豆科(Leguminosae)笔花豆属(Stylosanthes Sw.)多年生草本或亚灌木,广泛分布于美洲、非洲和亚洲的热带和亚热带地区[19],具有粗蛋白含量高、品质好、抗旱能力强、生产力高和适应性强等特点,在我国南方地区可用于饲料作物、绿肥覆盖、水土保持以及草地改良等[19-20]。‘热研2号’柱花草(S. guianensis ‘Reyan No.2’)是由中国热带农业科学院选育出的优良栽培品种,是目前最适合南方地区种植的热带豆科牧草品种之一[21]。课题组前期研究已发现,在植株体没有直接接触的情况下,‘热研2号’柱花草对不同亲缘关系植物的根系分泌物有识别与响应差异,近亲缘植物的根系分泌物处理时其生长无显著变化,远亲缘植物根系分泌物作用下其生长变化较为激烈[22];但作为混播邻居实际伴生生长时,‘热研2号’柱花草的对邻居的识别与响应情况尚不明确。张德等[23]已指出种间相互作用不可避免地会使植物在地上部和地下部产生不同生长策略。柱花草在田间栽培时不但可能与不同亲缘关系品种混播,还会面临杂草侵入危害这一现状,面对不同种类邻居,‘热研2号’柱花草的生长对策及其地上部与地下部对邻近植物的识别与响应差异对决定其与邻近植物的群落状态具有重要生态意义。因此,本试验通过在土培条件下研究‘热研2号’柱花草分别与不同亲缘品种/种及不同种杂草为邻近植物时表型特征参数和生物量分配策略的变化,揭示‘热研2号’柱花草对不同邻近植物的识别与生物响应策略,探究其在不同邻近植物影响下地上部与地下部的响应策略差异,为柱花草多品种混播栽培及柱花草草地杂草管理提供科学参考。
1 材料与方法
1.1 试验材料
共选取10 种植物种子,包括‘热研2号’、‘热研5号’(S. guianensis ‘Reyan No. 5’)、‘热研21号’(S. guianensis ‘Reyan No. 21’)、‘热研24号’(S. guianensis ‘Reyan No. 24’)、大叶柱花草(S. grandifolia)、西卡柱花草(S. scabra)和细茎柱花草(S. gracilis)共7种豆科牧草种子及鬼针草(Bidens pilosa)、蒺藜草(Cenchrus echinatus)和羽芒菊(Tridax procumbens)共3种杂草种子进行试验。7种柱花草种子均由中国热带农业科学院热带作物品种资源研究所热带牧草中期备份库提供,3种杂草种子采集自海南大学儋州校区教学基地及附近。
供试土壤选取于儋州校区农科基地0~30 cm表层的砖红壤。将土壤阴凉风干后过200目土壤筛装盆备用。土壤的基本理化性质为[24]:pH值为5.34、土壤有机质含量为10.91 g·kg-1,全氮含量为0.52 g·kg-1,速效磷含量为41.70 mg·kg-1,速效钾含量为79.82 mg·kg-1。
1.2 试验方法
1.2.1 试验设计 在海南大学儋州校区农科基地(19°30′N,109°29′E)温室大棚内进行试验。试验期间大棚内平均气温28℃左右,平均湿度85%左右。
以‘热研2号’柱花草为主要研究对象,参考王俐媛等[22]对‘热研2号’柱花草的不同亲缘植物的界定,‘热研2号’柱花草与其自身品种伴生为自我处理,即对照组,其他9种植物分别为‘热研2号’柱花草的邻居植物,共组成“自我(对照)、近亲、远亲、非亲”4个邻居关系处理组。其中:‘热研5号’、‘热研21号’和‘热研24号’为‘热研2号’柱花草的种内近缘亲属品种,简称近亲;大叶、西卡和细茎柱花草为‘热研2号’柱花草的种间远缘亲属品种,简称远亲;鬼针草、蒺藜草和羽芒菊为‘热研2号’柱花草的种间非亲属品种,简称非亲。参考赵元辰等[24]和李洁等[25]的试验方法,采用土壤盆栽法,将‘热研2号’柱花草分别与10 种植物配对,两种植物各1株、株距6.0 cm移栽于规格为14 cm(上径)×12 cm(下径)×16 cm(高)花盆中心线两侧,两植株之间分别距花盆中心线3.0 cm。每个组合3个重复。试验处理编号及邻居关系详见表1。
1.2.2 试验方法与指标测定 柱花草的种子在播种前用80℃热水浸种3~5 min,鬼针草、蒺藜草和羽芒菊种子保持干燥,然后将10种种子分别播于育苗盘,待幼苗长至株高约15 cm时,挑选株高基本一致的植株按试验设计配对移栽。整个生长过程中及时拔除长出的杂草以消除其他植物对试验的影响。每2 d按土壤最大持水量的60%浇水1次,并随机转动花盆以消除光照、边际等环境影响。试验期间不施肥。
待各移栽植株缓苗5 d后开始计数试验天数。生长30 d后,先测量每株植物的伸直株高,再记录每株植株的一级分枝数,然后将盆栽内的植株连同土壤一起小心取出,用清水冲洗干净根系,将每株植物(根、冠连在一起)做好标记带回实验室。在室内测量主根长度和一级侧根数,再分开每株植物的地上部和地下部,置于105℃鼓风干燥箱杀青30 min后在70℃烘干至恒重,用感量为0.1 mg的分析天平称量各部分干重,分别记录每株植物根生物量、茎叶生物量,计算各株植物的总生物量和根冠比。
1.3 数据处理与分析
使用Microsoft Excel进行数据整理,对不同邻居关系处理组之间和不同邻近植物伴生下‘热研2号’柱花草的各检测指标进行单因素方差分析(one-way ANOVA),用Duncan法在P<0.05水平进行多重比较。用主成分分析法(Principal component analysis,PCA)综合评判‘热研2号’柱花草占据主导地位的表型响应指标及对9种邻居植物表型响应强弱。首先将各处理组数据相对自我处理组的变化百分率进行Z-score法标准化处理,然后通过KOM检验法和Bartlett球体检验法进行因子分析的适用性检验,再计算主成分权重系数,根据主成分权重系数和各主成分对应的特征值计算因子得分和综合得分,获得综合排名。使用GraphPad Prism 8(GraphPad Software Inc.,San Diego,CA,USA)制作图表。统计分析使用IBM SPSS Statistics 22.0(SPSS Inc.,Chicago,IL,USA)。
2 结果与分析
2.1 邻近植物对‘热研2号’柱花草地上部生物响应的影响
2.1.1 邻近植物对‘热研2号’柱花草株高的影响 自我、近亲、远亲、非亲这4个处理组之间,‘热研2号’柱花草的株高均值均无显著差异(图1)。以‘热研5号’柱花草为邻时,‘热研2号’柱花草的株高显著降低20.18%(P<0.05),以其他邻居植物为邻时,‘热研2号’柱花草的株高均无显著变化(图1)。
2.1.2 邻近植物对‘热研2号’柱花草分枝数的影响 与自我处理组的分枝数均值相比,虽然近亲处理组、远亲处理组和非亲处理组‘热研2号’柱花草分枝数的均值呈现逐渐减少的趋势,但4个处理组之间差异均不显著(图2)。与大叶柱花草、蒺藜草和羽芒菊为邻的‘热研2号’柱花草分枝数无显著差异,比以自我为邻时显著降低(降幅均值21.70%)(P<0.05);与‘热研5号’‘热研21号’‘热研24号’、西卡柱花草、细茎柱花草和鬼针草为邻时,‘热研2号’柱花草分枝数与以自我为邻时均无显著差异(图2)。
2.2 邻近植物对‘热研2号’柱花草根系生长的影响
2.2.1 邻近植物对‘热研2号’柱花草根长的影响 近亲处理组‘热研2号’柱花草的根长均值与自我处理组无显著差异,分别比远亲处理组和非亲处理组的均值显著高出39.78%和46.61%(P<0.05);远亲处理组与非亲处理组‘热研2号’柱花草的根长均值间差异不显著,二者分别比自我处理组的根长均值显著降低32.61%和40.22%(P<0.05)(图3)。与‘热研5号’为邻时‘热研2号’柱花草的根长无显著变化;与‘热研21号’和‘热研24号’为邻时的根长间无显著差异,平均比与自我为邻时显著增加17.32%(P<0.05);与细茎柱花草和羽芒菊为邻时的根长间也无显著差异,平均比以自我为邻时显著降低26.13%(P<0.05);与大叶、西卡和蒺藜草为邻时的根长间也无显著差异,平均比以自我为邻时显著降低37.55%(P<0.05);与鬼针草为邻时根长显著降低幅度最大(53.60%)(P<0.05)(图3)。
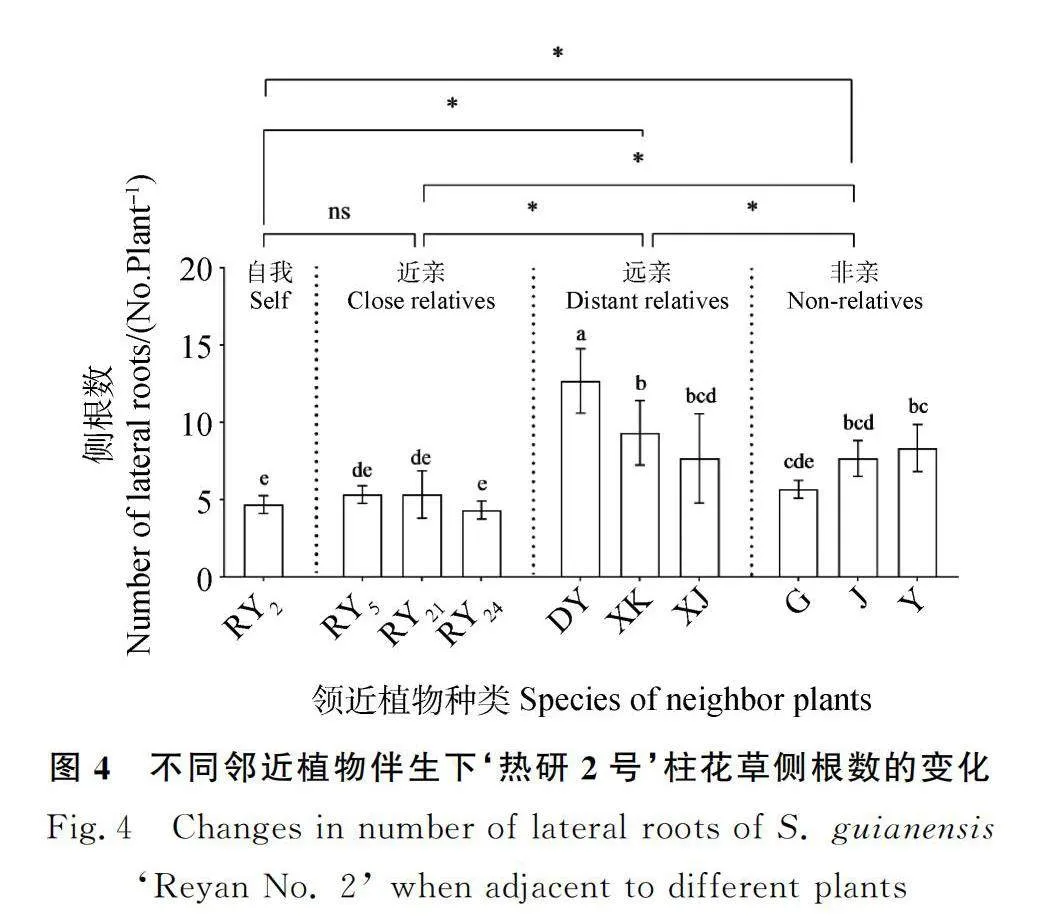
2.2.2 邻近植物对‘热研2号’柱花草侧根数的影响 近亲处理组与自我处理组的侧根数均值无显著差异;远亲处理组的侧根数均值显著大于其他处理组的对应值(P<0.05),分别比自我处理组、近亲处理组和非亲处理组的侧根数均值显著增大111.78%,97.92%和36.98%;非亲处理组的侧根数均值分别比自我处理组和近亲处理组的侧根数均值显著增大54.68%和44.49%(P<0.05)(图4)。与大叶柱花草为邻时‘热研2号’柱花草的侧根数最多,比与自我为邻时显著增加171.31%(P<0.05);与西卡柱花草、细茎柱花草、蒺藜草和羽芒菊为邻时的侧根数间无显著差异,平均比以自我为邻时显著增加76.65%(P<0.05);与‘热研5号’‘热研21号’‘热研24号’和鬼针草为邻时的侧根数与以自我为邻时均无显著差异(图4)。
2.3 邻近植物对‘热研2号’柱花草生物量积累和分配的影响
2.3.1 邻近植物对‘热研2号’柱花草生物量的影响 除近亲处理组的根生物量均值与自我处理组的根生物量均值无显著差异之外,自我、近亲、远亲和非亲4个处理组的茎叶生物量均值、根生物量均值和总生物量均值的对应值之间均存在显著差异(P<0.05)。其中近亲处理组的各生物量均值最大,分别比自我处理组的对应均值高43.30%,9.23%和33.76%(P<0.05);远亲处理组的茎叶生物量和总生物量均值最低,分别比自我处理组的对应均值显著降低45.83%和55.15%(P<0.05);非亲处理组各生物量均值分别比自我处理组对应均值显著降低23.22%,79.14%和34.41%(P<0.05),但其根生物量均值与远亲处理组的对应均值无显著差异,茎叶生物量均值和总生物量均值分别比远亲处理组的对应均值显著增加41.74%和46.23%(P<0.05)(图5)。
‘热研2号’柱花草的总生物量在以‘热研21号’‘热研24号’、鬼针草和‘热研5号’为邻时均显著增加,且增加量依次显著减小(P<0.05);以大叶柱花草、西卡柱花草、细茎柱花草、蒺藜草和羽芒菊为邻时总生物量均显著降低(P<0.05),其中以西卡柱花草为邻时降幅最小,以羽芒菊为邻时降幅最大,与其他3种植物为邻时的降幅间无显著差异(图5)。
根生物量仅在与‘热研21号’为邻时显著增加25.02%(P<0.05);与‘热研5号’和‘热研24号’为邻时均与以自我为邻无显著差异;与其它6种植物为邻时根生物量均显著降低(P<0.05),其中与鬼针草为邻时降低28.39%,其他5种彼此无显著差异(图5)。茎叶生物量在与‘热研5号’‘热研21号’‘热研24号’和鬼针草为邻时均显著增加(P<0.05),其中与‘热研21号’为邻时增幅最大,与‘热研24号’和鬼针草为邻时无显著差异,与‘热研5号’为邻时增幅最小;与其他5种植物为邻时茎叶生物量均显著降低(P<0.05),其中与西卡柱花草为邻时降幅最小,与羽芒菊为邻时降幅最大,其他3种植物为邻时彼此无显著差异(图5)。
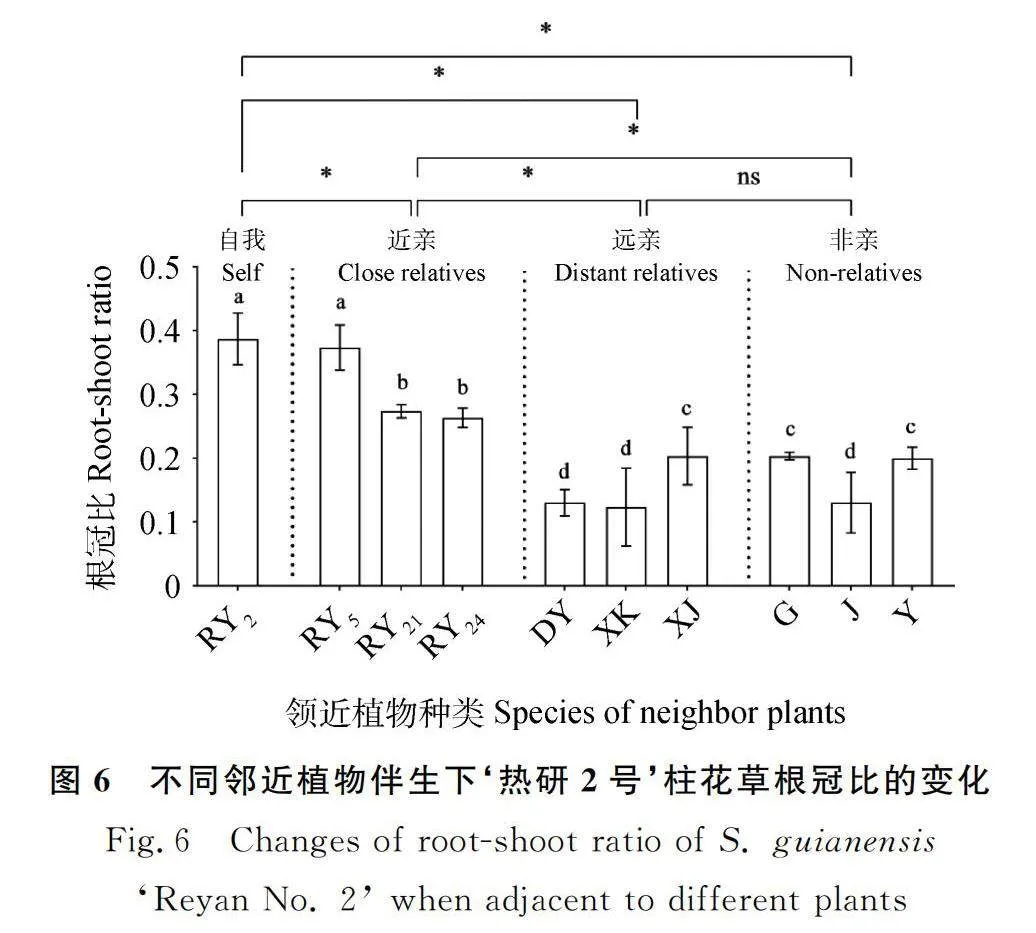
2.3.2 邻近植物对‘热研2号’柱花草根冠比的影响 近亲处理组、远亲处理组和非亲处理组的根冠比均值分别比自我处理组的对应均值显著降低21.57%,61.21%和54.30%(P<0.05),且远亲处理组和非亲处理组的根冠比均值之间无显著差异;而远亲处理组和非亲处理组的根冠比均值分别比近亲处理组的对应均值显著降低50.54%和41.74%(P<0.05)(图6)。与‘热研5号’为邻时‘热研2号’柱花草的根冠比与自我为邻时无显著差异;与其他8种植物为邻时的根冠比均显著降低(P<0.05),其中与‘热研21号’和‘热研24号’为邻时根冠比间无显著差异,平均比与自我为邻时显著降低30.75%(P<0.05);与细茎柱花草、鬼针草和羽芒菊为邻时根冠比间也无显著差异,平均显著降低47.80%(P<0.05);与大叶柱花草、西卡柱花草和蒺藜草为邻时根冠比间也无显著差异,平均显著降低66.93%(P<0.05)(图6)。
2.4 综合分析
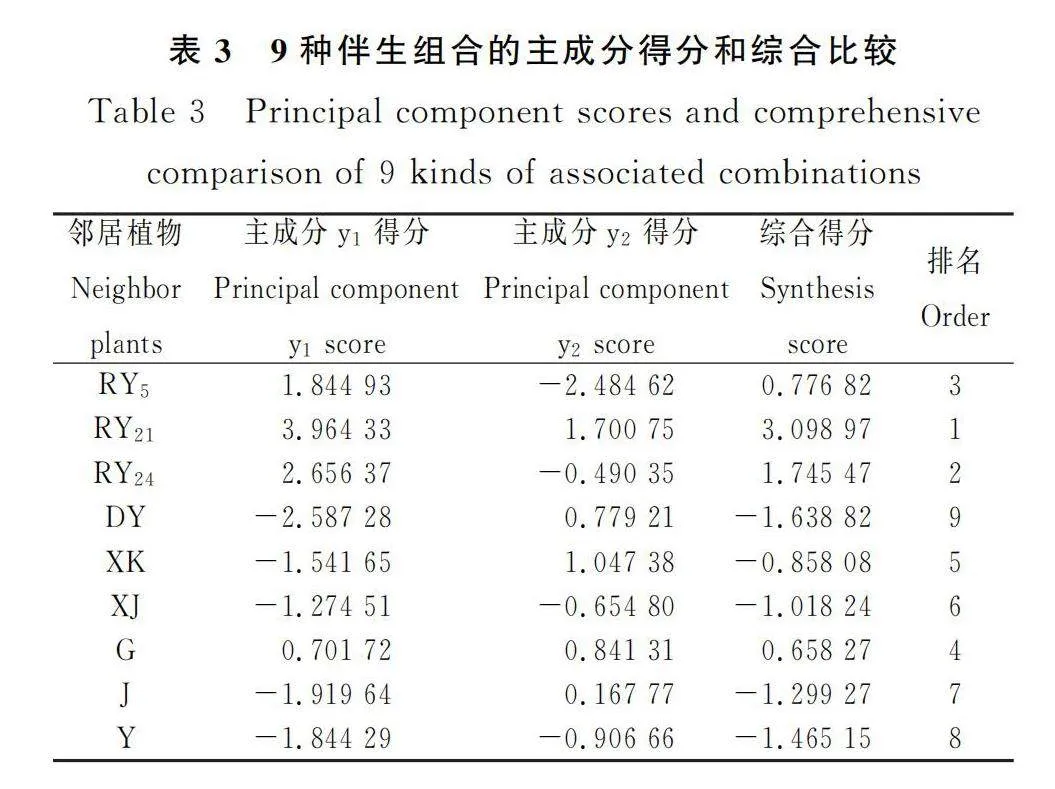
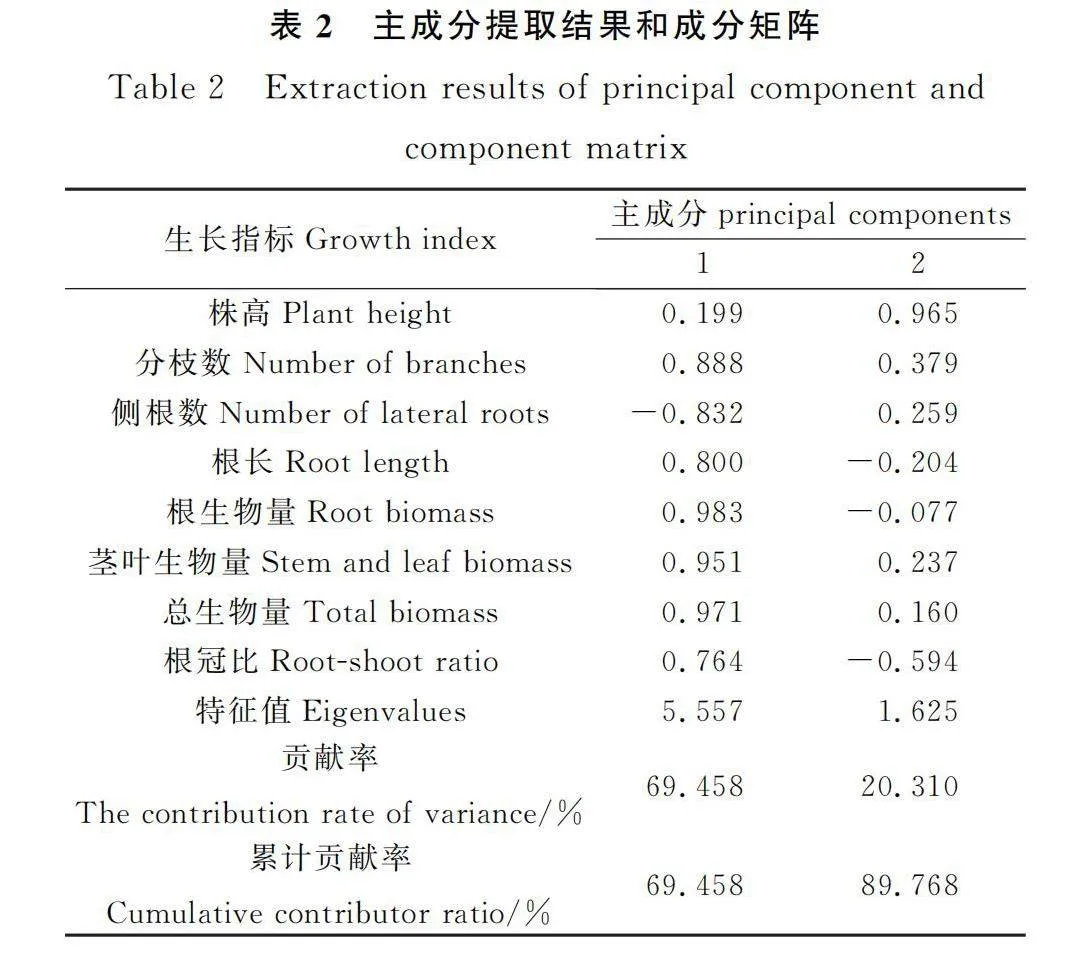
对9个邻居植物伴生下‘热研2号’柱花草的8个生长指标进行主成分分析可知:前2个主成分的累计贡献率达到89.768%(表2),其中第1主成分的贡献率最大,达69.458%,与第1主成分相关性相对较大(特征向量较大)的生物响应指标为根生物量、总生物量和茎叶生物量;第2主成分的贡献率20.310%,与第2主成分相关性相对较大的表型指标为株高、分枝数和侧根数。通过计算主成分得分和综合得分发现(表3),‘热研2号’柱花草对9种邻居植物植物表型响应得分由大到小依次为:‘热研21号’>‘热研24号’>‘热研5号’>鬼针草>西卡柱花草>细茎柱花草>蒺藜草>羽芒菊>大叶柱花草。
3 讨论
为了获取更多的资源以实现个体生长发育和种群繁衍,植物不但具有识别邻近植物的能力,还能够根据邻近植物的具体身份选择合适的生长策略[17],一般尽量减少与亲缘植株之间的竞争来获得更大的广义适合度[25]。本研究的4类邻居关系处理组中,相对于自我处理组,与近亲品种为邻时‘热研2号’柱花草的总生物量和茎叶生物量均值有所增大、根冠比均值有所降低,其株高、分枝数、根长、侧根数和根生物量等均值均无显著变化。这一结果虽然与美洲海滩芥(Cakile edentula)[26]、大豆[14]等对近邻的识别与响应情况并不完全一致,但都支持了植物的亲缘竞争回避策略[27-28],即识别到邻近植株为近亲缘植物后,植物根系部分基本维持原本的生长状态以避免与亲缘植株彼此间的地下竞争,而将现有的生物量更多地分配到地上部进行营养生长或生殖生长,以促进自身和同一种群的发展。与远亲种和非亲种为邻时‘热研2号’柱花草均表现出了根长、生物量和根冠比均值显著降低、侧根数显著增加的生长响应方式,说明受亲缘关系较远甚至无亲缘关系的邻近植物的影响,‘热研2号’柱花草的地上部和地下部生长都显著减弱,特别是地下部生长变化更为激烈,通过控制主根深扎、增强侧根数量的策略以获取更多的养分资源。
相关研究已表明,植物主要通过邻近植物的根系分泌物来识别邻近植物与自己的亲缘关系,从而决策生长响应方式[22,29],但具体响应结果除了与物种特性有关外,还受生态因子等的制约,一般表现为与陌生种一起生长时,在光照有限的环境中增加地上竞争性状,在养分有限的环境中增加地下竞争性状[26,30]。‘热研2号’柱花草对邻近植物的生长响应趋势与Dudley等[26]报道美洲海滩芥与陌生种为邻时根系生物量高于其与近亲种为邻时的生长响应趋势相反。这可能是亲缘识别与响应的效应与决定竞争结果的其他因素的作用效果相比较小,被掩盖或被忽略了[31],需要进行更精确、更全面的验证。面对4类邻近植物的生长策略变化,也说明‘热研2号’柱花草地下部比地上部的可塑性更强,更容易受到邻近植物竞争的影响,其主要的响应参数在于根系构型和根系生物量,这与主成分分析结果基本一致,也支持了Li等[32]根系构型在植物种间相互作用中具有重要意义这一观点。
在各处理组内部,不同品种/种植物与‘热研2号’柱花草为邻时,‘热研2号’的生长情况也各有差异。近亲组内,‘热研2号’和‘热研5号’均为CIAT184柱花草的常规选育品种[33],‘热研21号’和‘热研24号’均为太空诱变品种[34-35],由育成途径可推测‘热研2号’与‘热研5号’的亲缘关系相对最近,与‘热研21号’和‘热研24号’相对较远。本次近邻识别与响应结果中‘热研2号’柱花草对‘热研5号’的生长响应最平缓,对‘热研21号’的响应最强,‘热研24号’居中,说明我们对于亲缘关系的推测是可靠的,也进一步佐证了通过减少亲缘植株竞争的有害后果、利于提高群体广义适应的亲缘选择理论[36]。远亲组内,与大叶柱花草为邻时‘热研2号’的生长变化最激烈,主要表现在侧根数显著增加而根长和分枝数显著减小。主成分分析表明与大叶柱花草为邻时的综合得分为绝对值最大的负值,表明‘热研2号’受到强烈竞争影响,生长表现最差;与西卡柱花草和细茎柱花草为邻时的综合得分虽然也是负值,但其绝对值相对较小,由此可推测大叶柱花草与‘热研2号’柱花草的亲缘关系较远,细茎、西卡柱花草和‘热研2号’柱花草的亲缘关系相对较近。这一结论与本课题组王俐媛等[22]仅用含有亲缘植物根系分泌物的营养液培养‘热研2号’柱花草得到的生长响应情况并不完全一致,其原因可能在于根系分泌物组分在土壤因子的作用下发生了一定变化[24,37],从而影响了其与近邻间的识别与响应效果,还需要在后续进行进一步试验验证。
非亲组内,与鬼针草为邻时‘热研2号’柱花草表现出正向响应结果(综合得分为正),在与鬼针草的竞争中保持一定的地上生长优势,使非亲处理组的茎叶生物量和总生物量均值显著高于远亲处理组的对应均值,其原因可能在于柱花草的根系分泌物对鬼针草有一定的生长抑制作用[38]。与蒺藜草和羽芒菊为邻时,‘热研2号’柱花草的生长响应为负效应(综合得分为负值),说明其在生长竞争中处于劣势,生物量积累减少。柱花草与非亲植物为邻时生长无显著规律性的原因可能在于3种非亲植物种间差异太大,种间响应结果主要由彼此在资源分配获取能力方面的差异决定,而不是亲缘选择效应决定[39]。鬼针草、蒺藜草和羽芒菊均为海南省的常见农林杂草,因此田间栽培柱花草时要及时清除杂草,选择亲缘品种混播以提高柱花草的产量,但幼苗之间的竞争或促进等相互作用结果是可变的,受复杂环境因素的影响[40],本试验仅比较了幼苗期对邻近植物识别与响应情况,今后要进一步研究柱花草在营养生长旺盛期和生殖生长过程中的响应情况来确定其完整的响应对策。
4 结论
与近亲缘植物为邻时,‘热研2号’柱花草的生物量显著增加,地上部正生长响应强烈;与远亲缘植物和非亲缘植物为邻时,‘热研2号’柱花草的根系构型变化大,生物量显著降低,整体生长变差。与同一处理组内部不同种/品种植物为邻时‘热研2号’柱花草的生长响应也有一定差异,主成分分析表明使‘热研2号’地上生长具有优势的邻近植物依次为‘热研21号’>‘热研24号’>‘热研5号’>鬼针草,生长变差的邻近植物依次为大叶柱花草>羽芒菊>蒺藜草>细茎柱花草>西卡柱花草。说明‘热研2号’柱花草能通过识别邻近植物的亲缘关系调整自身生长策略,可选择近亲缘品种混播来提高柱花草的产量,但苗期生长中柱花草对杂草不具有竞争优势,需要及时清除。
参考文献
[1]陈仁飞,姬明飞,关佳威,等. 植物对称性竞争与非对称性竞争研究进展及展望[J]. 植物生态学报,2015,39(5):530-540
[2]陈柯岐,邓星光,林宏辉. 植物响应非生物胁迫的分子机制[J]. 生物学杂志,2021,38(6):1-8
[3]DE KROON H,HUBER H,STUEFER J F,et al. A modular concept of phenotypic plasticity in plants[J]. New Phytologist,2005,166(1):73-82
[4]LAW R,PURVES D W,MURRELL D J,et al. Causes and effects of small-scale spatial structure in plant populations[R]. IIASA Interim Report,Blackwell Science,Oxford,2001:21-44
[5]BENOT M L,BITTEBIERE A K,ERNOULT A,et al. Fine-scale spatial patterns in grassland communities depend on species clonal dispersal ability and interactions with neighbours[J]. Journal of Ecology,2013,101(3):626-636
[6]朱润军,杨巧,李仕杰,等. 植物表型可塑性对环境因子的响应研究进展[J]. 西南林业大学学报(自然科学),2021,41(1):183-187
[7]孔垂华. 植物种间和种内的化学作用[J]. 应用生态学报,2020,31(7):2141-2150
[8]熊韫琦,赵彩云. 表型可塑性与外来植物的成功入侵[J]. 生态学杂志,2020,39(11):3853-3864
[9]KAUR A,KAUR S,SINGH H P,et al. Phenotypic variations alter the ecological impact of invasive alien species:lessons from Parthenium hysterophorus[J]. Journal of Environmental Management,2019,241:187-197
[10]徐满厚,刘敏,翟大彤,等. 植物种间联结研究内容与方法评述[J]. 生态学报,2016,36(24):8224-8233
[11]MURPHY G P,SWANTON C J,VAN ACKER R C,et al. Kin recognition,multilevel selection and altruism in crop sustainability[J]. Journal of Ecology,2017,105(4):930-934
[12]DUDLEY S A,MURPHY G P,FILE A L. Kin recognition and competition in plants[J]. Functional Ecology,2013,27(4):898-906
[13]RUNYON J B,MESCHER M C,MORAES C M.Volatile chemical cues guide host location and host selection by parasitic plants[J]. Science,2006,313(5795):1964-1967
[14]MURPHY G P,VAN ACKER R,RAJCAN I,et al. Identity recognition in response to different levels of genetic relatedness in commercial soya bean[J]. Royal Society Open Science,2017,4(1):e160879
[15]雷桂生,王五云,蒋智林,等. 紫茎泽兰与伴生植物小藜的竞争效应及其生理生化特征[J]. 生态环境学报,2014,23(1):16-21
[16]DE KROON H. How do roots interact?[J]. Science,2007,318(5856):1562-1563
[17]KONG C H,ZHANG S Z,LI Y H,et al. Plant neighbor detection and allelochemical response are driven by root-secreted signaling chemicals[J]. Nature Communications,2018,9(1):3867
[18]CAHILL J F,MCNICKLE G G,HAAG J J,et al. Plants integrate information about nutrients and neighbors[J]. Science,2010,328(5986):1657-1657
[19]蒋亚君,申晴,丁西朋,等. 柱花草种质资源表型性状的多样性分析[J]. 草业科学,2017,34(5):1032-1041
[20]李粤,翁绍捷,邹冬梅. 热研2号柱花草干燥特性及粗蛋白质变化的试验研究[J]. 农业工程学报,2006,22(3):125-128
[21]杨帆,张宇,余爱,等. 配方施肥对热研2号柱花草产量和品质的影响[J]. 草业学报,2012,21(2):264-270
[22]王俐媛,王坚,赵元辰,等. ‘热研2号’柱花草对不同亲缘植物根系分泌物的生长响应[J]. 草业科学,2022,39(2):309-317
[23]张德,龙会英. 种间相互作用对牧草间作体系生产力的影响[J]. 干旱地区农业研究,2017,35(2):234-239
[24]赵元辰,李雪枫,王俐媛,等. 根系分隔方式对‘热研5号’圭亚那柱花草亲缘识别与选择的影响[J]. 草业科学,2023,40(2):491-501
[25]李洁,庞蕊,徐兴良,等. 高粱对不同身份邻居的形态学与生理学响应[J]. 应用与环境生物学报,2017,23(5):800-805
[26]DUDLEY S A,FILE A L. Kin recognition in an annual plant[J]. Biology Letters,2007,3(4):435-438
[27]肖万欣,王延波,张书萍. 临近根系间的亲缘识别、行为应对及其功能性状可塑性进化[J]. 辽宁农业科学,2021(2):48-53
[28]林威鹏,彭莉,肖桃艳,等. 植物亲缘识别的研究进展[J]. 植物生态学报,2015,39(11):1110-1121
[29]BIEDRZYCKI M L,JILANY T A,DUDLEY S A,et al. Root exudates mediate kin recognition in plants[J]. Communicative & Integrative Biology,2010,3(1):28-35
[30]MURPHY G P,DUDLEY S A. Kin recognition:competition and cooperation in Impatiens (Balsaminaceae)[J].American Journal of Botany,2009,96(11):1990-1996
[31]GRIFFIN A S,WEST S A. Kin selection:fact and fiction[J]. Trends in Ecology & Evolution,2002,17(1):15-21
[32]LI L,SUN J H,ZHANG F S,et al. Root distribution and interactions between intercropped species[J]. Oecologia,2006(147):280-290
[33]刘国道,白昌军,何华玄,等. 热研5号柱花草选育研究[J]. 草地学报,2001,9(1):1-7
[34]白昌军,刘国道. 柱花草良种繁育技术与管理[M]. 南京:江苏科学技术出版社,2013:37-38
[35]金沙蝴蝶豆、热研24号柱花草、黔北蕉芋、滇东北薏苡4个新品种通过国家牧草品种审定[J]. 世界热带农业信息,2020(2):3
[36]李洁,孙庚,胡霞,等. 植物的亲缘选择[J]. 生态学报,2014,34(14):3827-3838
[37]吴清莹,林宇龙,孙一航,等. 根系分泌物对植物生长和土壤养分吸收的影响研究进展[J]. 中国草地学报,2021,43(11):97-104
[38]王俐媛,王坚,李雪枫,等. 根系分泌物在‘热研2号’柱花草与鬼针草生长早期种间干扰中的作用[J]. 草地学报,2022,30(11):3018-3025
[39]CHEPLICK G P,KANE K H. Genetic relatedness and competition in Triplasis purpurea (Poaceae):resource partitioning or kin selection?[J]. International Journal of Plant Sciences,2004,165(4):623-630
[40]PARISE A G,BERTOLI S C,SOUZA G M. Belowground interactions affect shoot growth in Eucalyptus urophylla under restrictive conditions[J]. Plant Signaling & Behavior,2021,16(9):1927589
(责任编辑 闵芝智)
