白玉兰二次开花诱导与生理调控研究
2024-10-09陈香波张冬梅傅仁杰张浪申亚梅罗玉兰尹丽娟


摘要:【目的】通过人工诱导实现白玉兰(Yulania denudata)二次开花,以探明白玉兰花芽分化与开花诱导的生理调控机制。【方法】采用施用营养肥与激素处理白玉兰植株,进行花芽发育石蜡切片观察,并测定不同时期花芽内源激素含量。【结果】5月上旬随着营养生长趋于减缓,白玉兰起始花芽分化,至6月上旬花芽分化基本完成;单独采用外源激素(6-BA、GA3)或配合施用营养肥(N、P、K、糖),可诱导白玉兰当年二次开花,二次开花株率33%~100%,单株最大开花量22;花芽分化成熟过程中,内源GA3与ABA含量升高,而打破休眠、开花的花芽内脱落酸(ABA)含量下降,内源生长素类激素(IAA、ME-IAA)含量升高;较未开花对照,外源细胞分裂素处理株花芽内ME-IAA、IPA、JA-Ile、JA含量较高,外源赤霉素(GA3)处理株内源tZR、GA1、GA3含量较高。【结论】外源激素可不依赖于营养处理直接诱导白玉兰二次开花,不同激素诱导白玉兰二次开花可能存在不同的开花调控通路。
关键词:白玉兰;花芽分化;二次开花;生理调控;营养;激素
中图分类号:S718;S685"""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)05-0097-08
Induction and physiological regulatory mechanism of secondary flowering in Yulannia denudata
CHEN Xiangbo1, ZHANG Dongmei1*, FU Renjie1, ZHANG Lang1, SHEN Yamei2, LUO Yulan1, YIN Lijuan1
(1.Shanghai Academy of Landscape Architecture Science and Planning, Shanghai Engineering Research Center of Landscaping on Challenging Urban Sites, Shanghai 200232, China; 2.School of Landscape Architecture, Zhejiang Agriculture and Forestry University, Hangzhou 311302, China)
Abstract: 【Objective】The aim of this"" study was to achieve the secondary flowering of Yulannia denudata through artificial induction and to explore the physiological regulation mechanism of flower bud differentiation and flowering induction in Y. denudata.【Method】Y. denudata plants were treated with nutrients and hormones. Flower buds were embedded in paraffin and sectioned to observe flower bud development, and endogenous hormone contents in flower buds at different stages were measured.【Result】Flower bud differentiation of Y. denudata began in early May when vegetative growth slowed down and was completed by early June. Secondary flowering within the same year was induced via treatment with exogenous hormones [6-benzylaminopurine (6-BA) and gibberellin A3 (GA3)] alone or in combination with nutrient applications [nitrogen (N), phosphorus (P)," potassium (K) and sugars]. The secondary flowering rate ranged from 33% to 100%, with" the" maximum of 22 flowers per plant. During the maturation of flower bud differentiation, the levels of endogenous GA3 and abscisic acid (ABA) increased. The ABA content in flower buds decreased while breaking dormancy and flowering, whereas the content of endogenous auxins (indole-3-acetic acid (IAA) and methylindole-3-acetic acid (ME-IAA) increased. Flower buds treated with exogenous cytokinins had higher levels of ME-IAA, IPA, jasmonoyl-l-isoleucine (JA-Ile)" and jasmonate (JA), while flower buds treated with exogenous gibberellins (GA3) had higher levels of endogenous trans-zeatin riboside (tZR), gibberellin A1 (GA1), and GA3. 【Conclusion】Exogenous hormones directly induce secondary flowering in Y. denudata without relying on nutrient treatments. Thus, different hormones may induce secondary flowering through various regulatory pathways.
Keywords:Yulannia denudata; differentiation of flower bud; secondary flowering; physiological regulation; nutrition; hormone
白玉兰(Yulania denudata)为木兰科(Magnoliaceae)玉兰属 (Yulania)开花乔木[1],春季先花后叶,花开满树、洁白高雅,被推选为上海市市花。上海地区白玉兰常年花期在2月下旬至3月上旬,随开花前气温变化会有所波动,早可在2月上旬、迟则至3月中旬,而开花延续长短也随着开花时节气温波动及雨水情况而变化。有研究表明,白玉兰始花期受当年1、2月的日照和温度影响最大,春季开花前1—2月的温度越高、光照越充足,白玉兰开花越早;温度低、光照少,则白玉兰开花延迟[2]。白玉兰花芽分化在前一年的夏初即已完成,每年只进行1次花芽分化,自4月底开始至6月中上旬结束,整个花芽分化期持续约40 d[3]。花芽分化完成至第2年早春开花期间,白玉兰花芽内部组织形态上只是经历花托伸长、拟花蕾稍膨大等微小变化,冬季落叶后进入休眠[4]。
玉兰属花芽分化与开花模式类型多样而复杂,既有白玉兰、紫玉兰(Y. liliflora)等花芽一次分化型,也有1年多次分化型,如紫玉兰‘红元宝’(Y. liliiflora ‘Hongyuanbao’)以及‘红元宝’杂交选育品种‘红笑星’(Yulania × Michelia ‘Star’)、‘红金星’(Yulania × Michelia ‘Sweet’)、‘红寿星’(Yulania × Michelia ‘Fairylake’)等,1年中会有2~3次开花,分别在3—4月、6—7月或8—10月开放,表现为二次开花或多季开花特性[5];此外玉兰属植物还存在因花芽分化与发育的不同步而表现出多次开花的类型,最典型的是‘常春’二乔玉兰(Yulania" × soulangeana ‘Changchun’),春季花后经过一段营养生长后分化花芽,部分花芽于6月膨大开花,但雌、雄蕊发育异常,另一部分花芽则继续缓慢发育至第2年春季开花,也有少量夏季开放花基部侧芽再次分化花芽而次年春季开花[6]。
白玉兰虽一年只进行1次花芽分化,但栽培中偶有发现个别白玉兰植株会在夏秋季节二次开花。目前,对于白玉兰花芽分化形态与生理调控机制的研究仍非常欠缺。本研究通过对白玉兰枝条、芽生长动态监测及花芽分化显微切片观察,掌握白玉兰营养与生殖生长规律,分阶段采用不同营养、激素处理白玉兰植株诱导实现二次开花,初步分析影响白玉兰花芽分化及开花的内源激素调控机制,为进一步揭示玉兰属开花调控的分子机理奠定基础。
1 材料与方法
试验于上海市园林科学规划研究院青松试验基地(121°26′E,31°12′N)进行。选取枝条开展、树形饱满的白玉兰植株为试验材料,规格为地径4.0~5.0 cm、株高1.8~2.0 m。于春季发芽前将白玉兰植株种植于装好园土、草炭、珍珠岩按2∶2∶1体积比例混合基质的无纺布种植袋中,常规除草管理。
1.1 处理方法
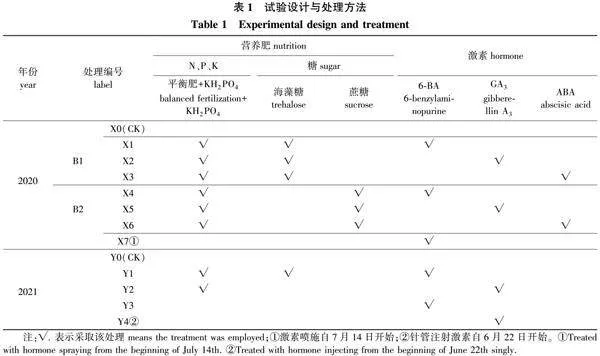
于2020年、2021年连续2年共进行11组处理及2组对照(每年1组对照),处理分3个阶段进行:第1阶段为白玉兰花后营养生长期,自4月上旬始至4月中旬止;第2阶段为生殖生长阶段,自4月下旬始至5月下旬止;第3阶段为花前准备期,自6月上旬始直至开花为止。
1)施肥处理。营养肥包括氮、磷、钾及糖等, 自4月上旬始至7月中旬进行N、P、K施肥:第1阶段根部浇施平衡肥[m(N)∶m(P2O5)∶m(K2O)=20∶20∶20],施用质量浓度1 g/L,每14 d浇施1次,第2、3阶段采用质量分数0.2%的磷酸二氢钾(KH2PO4)溶液每7 d喷施1次。糖类施用为在第1、2阶段对准地上部分树体叶片部位,用喷壶雾状喷施海藻糖或蔗糖溶液,质量浓度为0.06 mol/L,每周喷施1次。
2)激素处理。2020年采用树冠喷施的方法,自第3阶段起,对准地上部分树冠部位分别喷施6-苄基腺嘌呤(6-BA)、赤霉素(GA3)或脱落酸(ABA)溶液,质量浓度分别为0.5、0.5、0.3 g/L,每14 d喷施1次。2021年前期(6月4日—21日)采用树冠喷施的方式,激素施用质量浓度0.8 g/L,每7 d喷施1次,每株喷施量200 mL。因树冠喷施药剂损耗,后期改为针管注射方式施用;在主干树冠分枝处以下2~3 cm处上下错位插入3~4支注射针管深至树干木质部,每管注入激素溶液20 mL/次,每隔1~2 d 补液1次,共计注射6次,激素施用质量浓度为1.0 g/L。
以上施用方法凡浇施均每株施用量为1 L,喷施则每株施用量100 mL。对照为不施用任何营养肥或激素,仅日常浇水管理。每处理9株,为3个重复。前期置于露地,主要依靠自然降水,视土壤干燥情况适度补水。自第3阶段始将所有处理白玉兰植株移入遮雨棚下,同期适度控水,仅在土壤干透前稍加补水,以不萎蔫为度,具体处理方法见表1。
1.2 测试方法
1.2.1 开花统计
统计二次开花的白玉兰植株数,记录二次开花时间。计算开花株率(%,开花株占处理株数之百分比);观察并统计二次开花的花朵数,计算平均单株开花量(合计单株开花量与处理株数之比),取平均值。对两年数据采用DPS统计软件分别进行方差分析及LSD多重比较[7]。
1.2.2 花芽发育形态观测
自2020年4月下旬始至6月上旬止,每周选取10个位于树冠中上部枝条顶端的花芽测量芽长及芽宽[8]。同时每2周采集3个芽,剥去外部包裹的芽鳞片,取中心芽用体积分数25%戊二醛溶液固定。在第3阶段激素处理前,X1、X2、X3均为海藻糖喷施处理,前期处理方法相同,为减少重复采样,采用混合取芽方式、合并记为B1处理;同理,X4、X5、X6合并记为B2处理,分别对B1、B2、X0(对照)取芽进行花芽发育石蜡切片观察。
1.2.3 内源激素含量测定
于2021年白玉兰花芽分化不同时期:花芽分化起始期、完成期,花芽膨大裂苞期(开花前期)对Y1、Y2、Y4处理进行取样。其中花芽分化起始期(5月10日)取的芽处于营养生长期结束、启动花芽分化的成花转变时期,样品标号后缀记为“-V”(vegetative period),花芽分化完成期(6月4日)属生殖生长阶段,样品标号后缀记为“-R”(reproductive period),花芽膨大裂苞期是即将裂苞启动开花的时期,样品标号后缀记为“-F”(flowering period), 采取对照Y0处理未开花花芽,样品标号记为“Y0-N”作为未开花对照(取样时间7月2日)。取至少2 g的芽材料混合后作为1个样品待测,每处理3个重复,放入10 mL贴好标签的离心管中,并立即投入干冰中速冻,短期运输后置于-70 ℃冰箱保存待测。测前迅速液氮固定,提取测定激素。
1.2.4 数据分析
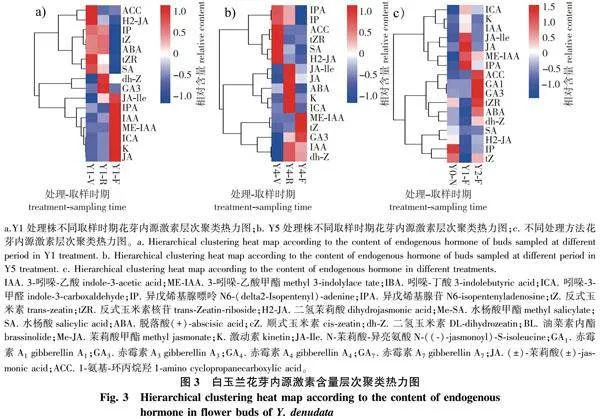
采用高通量靶标代谢组学检测24种植物内源激素,测试完成后对内源激素的定量值计算欧氏距离矩阵(Euclidean distance matrix),以完全连锁方法对内源激素代谢物聚类,并以热力图进行分析,绘制层次分析聚类热力图[9]。
2 结果与分析
2.1 白玉兰花芽发育与分化
白玉兰自花谢后即进入营养生长期,3月下旬—4月枝条快速生长,5月上旬开始,伴随枝条营养生长减缓,白玉兰进入生殖生长期、启动花芽分化。花芽分化前的白玉兰短枝顶芽细长瘦弱,自 5月上旬开始顶芽明显膨大变粗,尤其是5月下旬至6月初,白玉兰花芽进入快速膨大期,芽长、芽宽增长近1倍。石蜡切片显微观察发现,白玉兰花芽分化经历花芽分化起始期(5月上旬)、花瓣原基分化期(5月中旬)、雄蕊原基与雌蕊原基分化期(5月下旬),至6月上旬花芽分化趋于完成进入花芽分化完成期。观察发现,营养施肥处理株花芽较未处理对照有一定数量增多,但各处理花芽分化时期早晚差异不明显(图1)。
2.2 白玉兰二次开花诱导
通过施肥与激素处理等综合措施运用可使早春已开花白玉兰植株年内二次开花(表2),所有二次开花的处理均在激素施用处理中实现,其中细胞分裂素处理(X1)与赤霉素处理(X5、Y2)达到100%的处理株开花。不同激素诱导效果不同,细胞分裂素与赤霉素可促使白玉兰花芽膨大、启动开花,而脱落酸处理(X3、X6)未能诱导实现白玉兰二次开花。从开花株率与平均单株开花量看,细胞分裂素处理与赤霉素处理间差异未达显著水平(Pgt;0.05)。前期未施用营养肥,仅在后期花芽分化完成后树冠喷施细胞分裂素(6-BA)或赤霉素(GA3),即可实现花芽膨大开花 (X7、Y3、Y4处理),可见激素可独立诱导白玉兰成熟花芽进入开花程序,营养肥施用不是开花的必要条件。
不同于早春先花后叶,夏季二次开花的白玉兰是花叶同在(图2e—2f)。X1处理开花时间最早,为7月14日(图2d),此时正值上海夏季高温季节,日最高温度在30 ℃以上,单朵花从花瓣伸展绽放到凋落枯萎、持续2~3 d,较春季开花持续期缩短2~3 d。后期以激素注射方式代替喷施,单株开花量从2020年的2~6朵/株增加至2021年16(Y1,图2e)与22朵/株(Y2,图2f),植株整体开花效果明显提升。
从开花早晚看,细胞分裂素处理组(X1、X4)诱导二次开花时间略早于赤霉素处理组(X2、X5),海藻糖营养施用处理组(X1、X2)略早于蔗糖处理组(X4、X5)。由激素使用开始至第1朵花开放需(30±10)d,随气温高低及注射方式不同而有变化。针管注射激素吸收效率高,诱导二次开花时间间隔短(Y4)。将起始激素处理时间推迟到7月14日(X7),则至开花仅需20 d左右,通过延后处理的方式能够顺延白玉兰二次开花时间,由此,可精准实现7—8月白玉兰二次开花目标。
细胞分裂素与赤霉素除了诱导白玉兰花芽膨大而开花外,植株生长表现也会发生一些变化。经细胞分裂素处理的白玉兰植株,部分叶枯焦落叶,伴有枝条隐芽、不定芽大量诱导萌生,后续会抽枝发叶(图2g);而赤霉素处理株枝条夏季休眠被打破而发枝伸长(图2h),这在正常白玉兰生长中较为少见。
2.3 内源激素含量变化
2.3.1 不同发育时期花芽内源激素含量变化
由营养生长转入生殖生长、开始花芽分化直至花芽膨大裂苞、开花的不同发育阶段,芽内源激素代谢发生着变化(图3a、3b)。由营养生长转入花芽分化成熟,白玉兰花芽内源激素变化表现为:幼嫩组织激素如细胞分裂素类,IPA(异戊烯基腺苷)、IP(异戊烯基腺嘌呤)、tZR(反式玉米素核苷)降低,而与组织成熟有关的激素 ABA(脱落酸)、GA3(赤霉素)含量增加,同时与花器官发育相关的激素JA(茉莉酸)与JA-Ile(N-茉莉酸-异亮氨酸)也有所增加。白玉兰花芽分化完成后即进入休眠,而使用外源激素(赤霉素或细胞分裂素)可促使花芽打破休眠而启动开花,内源激素调控变化主要为生长素类激素IAA(吲哚-3-乙酸)或ME-IAA(吲哚-3-乙酸甲酯)增加,而与成熟及衰老相关的激素SA(水杨酸)、H2-JA(二氢茉莉酸)、ABA(脱落酸)则有所下降。发育成熟的花芽内源ABA含量较高,但经外源细胞分裂素(6-BA)与赤霉素(GA3)处理后,花芽内脱落酸(ABA)含量下降,内源生长素类激素(IAA、ME-IAA)则维持在较高水平,这些激素变化必然与打破花芽休眠促使花苞开裂而开花存在一定联系。
2.3.2 不同处理方法花芽内源激素含量变化
花芽分化完成后,外源激素(6-BA、GA3)处理阻断白玉兰已发育成熟花芽休眠而直接启动开花程序进而开花。较未开花对照(Y0-N),外源细胞分裂素(6-BA)处理植株花芽ME-IAA、IPA、JA-Ile、JA含量较高,而外源赤霉素类(GA3)处理植株花芽内源tZR、GA1、GA3含量较高(图3c)。
3 讨 论
3.1 开花诱导效应
自然条件下,白玉兰花芽分化完成后,即进入长达8个月的休眠期,至第2年早春随着气温回升,花芽开始膨大、启动开花。白玉兰花芽发育完全后,施用外源细胞分裂素或赤霉素,可诱导白玉兰成熟花芽即时启动开花程序,且不依赖于前期营养积累。激素适合的施用应在6月中、下旬白玉兰花芽分化完成以后进行,树干注射激素较叶面喷施对白玉兰二次开花效果更直接,这可能与喷施损耗大且易造成叶表面糖累积堵塞气孔而影响叶片吸收有关。
开花是一个能量消耗的过程,需要一定的营养储备。对‘常春’二乔玉兰研究中发现,春、夏两次成花过程中枝条可溶性糖含量均有不同程度的降低,可溶性糖利于开花的实现[6]。糖在植物体内特别是花芽中的含量与开花显著相关。糖作为一种信号因子调控植物内源激素水平从而调控开花[10],外施蔗糖或者海藻糖可加快‘红元宝’紫玉兰二次花芽分化的进程,尤其是第2次分化花芽中的海藻糖和蔗糖含量有显著提高,海藻糖可能参与了内源激素的运输[11-12],通过调控激素的合成与分解途径而影响内源激素水平[13],而在花芽分化与开花中扮演着重要角色。从本研究结果来看,外施营养元素可增进树体营养积累,促进枝条营养生长,一定程度促进白玉兰花芽分化,但不是开花诱导的必然条件。
后续观察发现,2020年夏季二次开花的白玉兰植株,次年(2021年)春季植株上留存的少量花芽会正常开花,花后继续花芽分化至第3年(2022年)春季开花,但第3年夏秋季赤霉素处理株(X2、X5)在无任何人为干预的情况下个别株“再次”二次开花。激素作用促使白玉兰二次开花,打破了植株正常的开花节律,并且在植株体内产生了“记忆”,亦可能是2022年夏季持续40 ℃极端高温带来的叠加效应,其内在原因值得进一步研究。据对2020、2021两年实验处理期间的气象因子对比,气温差别并不显著,二次开花的时间主要与激素处理时间有关,且激素的有效吸收直接关系到二次开花效果。
3.2 内源调控机制
3.2.1 不同发育时期内源激素调控
比较不同发育时期花芽内源激素含量变化,自花芽分化起始至花芽分化完成期间, 伴随花芽分化成熟,内源GA3含量明显增加,ABA有不同程度的增加, tZR、IP与IPA等细胞分裂素类激素均有所下降,其中tZR降低明显;后期自花芽分化完成期至花芽膨大裂苞期间,虽施用的是不同的外源激素,但花芽成熟裂苞、转而开花的内源激素变化存在共同点,即IAA、ME-IAA升高,而ABA降低,外源激素的施用影响到花芽内部激素的平衡。由此分析,在花芽分化起始到成熟过程中,GA3、ABA升高有助于花芽分化成熟,而打破花芽休眠、进入开花程序则需要较高水平的IAA以及低水平的ABA,这与肖爱华等[14]在红花玉兰(Magnolia wufengensis)以及牛辉陵等[15]在枣(Ziziphus jujuba)中的研究结果一致,GA3、ABA、IAA等植物内源激素参与了白玉兰花芽分化与开花调控。
3.2.2 不同处理方法内源激素调控
白玉兰植株在花芽分化完成至启动开花过程中,外源施用细胞分裂素诱导内源JA与JA-Ile增加,茉莉酸在植物衰老过程中发挥着重要作用,尤其是在促进植物叶片衰老方面,这也解释了6-BA处理植株出现部分叶枯焦并伴随落叶的现象,可能是白玉兰植株启动胁迫诱导反应的结果。除此以外,细胞分裂素处理诱导IPA增加,IPA为一种结合态细胞分裂素结合在tRNA上,构成tRNA的组成成分,可诱导植物细胞的分裂与增殖,在诱导芽的分化、叶绿体发育、养分运输及植物抗衰老等方面发挥着作用,白玉兰大量不定芽诱导增生可能与此有关。
外源赤霉素处理与细胞分裂素处理株内源激素调控差异较大,外施GA3导致内源赤霉素类物质GA1、GA3显著增加,从而诱导白玉兰植株开花,这与孙斌[16]、温璐华等[17]研究发现高水平GA3 可促进金银花(Lonicera japanica)、果梅(Prunus mume)等植物开花、使花期提前的结果一致。分子研究揭示:赤霉素可通过GAF1-TPR复合体抑制开花抑制因子而促进植物提早开花[18],赤霉素具有促使植物细胞伸长、茎伸长的作用[19],这也解释了为何外源赤霉素处理使得夏季本已停止生长的白玉兰植株枝条再次发枝伸长。Kinet[20]认为,低浓度IAA 含量为花芽孕育所必需,而打破花芽休眠开花则需要高浓度生长素IAA或GA3等赤霉素类内源激素的积累,在白玉兰开花的诱导调控中得到证实,并且内源激素间的平衡在植物开花调控中扮演着重要角色[21]。
3.2.3 开花诱导调控通路
植物开花调控是一个复杂的过程,受多种外界环境因子如营养条件、光照、温度、水分状况等影响[22], 而开花诱导存在多种调控通路,包括光周期(光依赖)、春化、自主、赤霉素、温度及年龄通路等[23-24],各通路间既彼此独立又互相交联,构成一个复杂的网络,实现对植物开花时间的精细调节[25]。白玉兰花芽对气温变化非常敏感,春季开花诱导主要受温度通路调控。赤霉素诱导白玉兰二次开花,证明白玉兰中存在赤霉素通路。细胞分裂素诱导白玉兰开花的同时出现枯叶、落叶现象,这与夏秋季节个别白玉兰植株提早落叶并开花现象极为类似。众所周知,叶枯萎、落叶往往与高温、低温或干旱脱水引起的胁迫反应有关,胁迫反应与激素调控存在某些交叉通路,比如在番茄研究中发现,受细胞分裂素诱导表达的下游基因也同时受低温冷胁迫、干旱脱水胁迫等的诱导[26]。模式植物拟南芥研究中发现,春化、温度、年龄与赤霉素途径是通过促进或抑制共同的下游开花整合基因SOC1基因,进而调控花分生组织基因AP1、CAL、FUL、LFY表达而诱导开花[27],SOC1基因的增强表达受高温和赤霉素共同诱导[28]。同样,转录组分析发现光周期、春化、年龄和赤霉素通路均参与到对‘红元宝’紫玉兰两次花芽分化的成花调控中[14]。温度、赤霉素共同参与了白玉兰的开花诱导调控,并可能与胁迫反应存在交叉通路,这再次证明了植物开花遗传调控通路的错综复杂性[29]。有关激素、营养与高温、干旱等环境因子诱导花芽分化及开花的交叉调控,还有待进一步从生理、分子生物学水平进行基因差异表达、代谢组学等研究分析,以揭示白玉兰花芽诱导及开花诱导与调控机制。
参考文献(reference):
[1]中国科学院中国植物志编辑委员会.刘玉壶.中国植物志.第30(1)卷.被子植物门.双子叶植物纲.防已科 木兰科[M].北京:科学出版社,1996. Chinese Academy of Sciences, Editorial Committee of Flora of China, LIU Y H. Flora of China.Vol. 30(1). Angiospermae. Dicotyledoneae. Menispermaceae. Magnoliaceae[M]." Beijing: Science Press,1996.
[2]舒素芳,毛俊萱,蔡敏.白玉兰始花期与气象因子的关系分析[J].浙江农业学报,2013,25(2):248-251.SHU S F,MAO J X,CAI M.Analysis on relationships between first-flowering dates of Magnolia denudata and meteorological factors[J].Acta Agric Zhejiangensis,2013,25(2):248-251.DOI: 10.3969/j.issn.1004-1524.2013.02.08.
[3]范李节,陈梦倩,王宁杭,等.3种木兰属植物花芽分化时期及形态变化[J].东北林业大学学报,2018,46(1):27-30,39.FAN L J,CHEN M Q,WANG N H,et al.Flower bud differentiation of three species of Magnolia[J].J Northeast For Univ,2018,46(1):27-30,39.DOI: 10.3969/j.issn.1000-5382.2018.01.006.
[4]武祖发,胡建军,斯华文,等.玉兰花芽分化观察[J].中国中药杂志,1991,16(2):79-81,126.WU Z F,HU J J,SI H W,et al.An observation on differentiation of Magnolia denudata Dess flower buds[J].China J Chin Mater Med,1991,16(2):79-81,126.
[5]张杰,张春英,王亚玲,等. 木兰珍品:多季开花木兰[J].园林,2018,11:56-59. ZHANG J,ZHANG C Y,WANG Y L.Magnolia treasures:multi-season flowering Magnolia[J]. Garden,2018,11:56-59.
[6]蒋政,孙李勇,刘旭,等.常春二乔玉兰春夏季开花节律及营养效应研究[J].植物研究,2019,39(2):192-199.JIANG Z,SUN L Y,LIU X,et al.Nutritional effect and rhythm of spring and summer flowering in Magnolia soulangeana ‘Changchun’[J].Bull Bot Res,2019,39(2):192-199.DOI: 10.7525/j.issn.1673-5102.2019.02.004.
[7]唐启义, 冯明光. DPS数据处理系统:实验设计、统计分析及数据挖掘[M]. 北京: 科学出版社,2007. TANG Q Y, FENG M G. DPS data processing system:experimental design, statistical analysis and data mining[M]. Beijing: Science Press,2007.
[8]陈建业,鲁国荣,宁玉霞,等.木兰属等3属植物的芽种类、结构与成枝规律研究[J].中国农学通报,2012,28(34):46-52.CHEN J Y,LU G R,NING Y X,et al.Study on the bud species,anatomical structure and the growth rule of branch forming in Magnolia Linn.,Yulania spach and Michelia Linn[J].Chin Agric Sci Bull,2012,28(34):46-52.DOI: 10.3969/j.issn.1000-6850.2012.34.009.
[9]KOLDE R. Pheatmap: pretty heatmaps [Z]. R package version, 2012.
[10]MATSOUKAS I G.Interplay between sugar and hormone signaling pathways modulate floral signal transduction[J].Front Genet,2014,5:218.DOI: 10.3389/fgene.2014.00218.
[11]程少禹.‘红元宝’紫玉兰两次花芽分化糖含量变化及转录组分析[D].杭州:浙江农林大学,2021.CHENG S Y.Sugar content and transcriptome analysis during two flower bud differentiation of Magnolia liliflora‘Hongyuanbao’[D].Hangzhou:Zhejiang A amp; F University,2021.
[12]程少禹,宣铃娟,董彬,等.‘红元宝’紫玉兰两次花芽分化差异代谢通路及关键调控基因筛选[J].园艺学报,2020,47(8):1490-1504.CHENG S Y,XUAN L J,DONG B,et al.Identification of differential metabolic pathways and key regulatory genes in the two flower bud differentiation processes of Magnolia liliiflora[J].Acta Hortic Sin,2020,47(8):1490-1504.DOI: 10.16420/j.issn.0513-353x.2019-0963.
[13]赵慧慧.高温胁迫下小麦中植物激素对外源海藻糖的响应[D].上海:华东师范大学,2016.ZHAO H H.The response of plant hormones to exogenously-supplied trehalose in wheat under high temperature stress[D].Shanghai:East China Normal University,2016.[14]肖爱华,陈发菊,贾忠奎,等.梯度洗脱高效液相色谱法测定红花玉兰中4种植物激素[J].分析试验室,2020,39(3):249-254.XIAO A H,CHEN F J,JIA Z K,et al.Determination of 4 plant hormones in Magnolia wufengensis by gradient elution high performance liquid chromatography[J].Chin J Anal Lab,2020,39(3):249-254.DOI: 10.13595/j.cnki.issn1000-0720.2019.051504.
[15]牛辉陵,张洪武,边媛,等.枣花分化发育过程及其内源激素动态研究[J].园艺学报,2015,42(4):655-664.NIU H L,ZHANG H W,BIAN Y,et al.Flower formation and endogenous hormones dynamic in Chinese jujube[J].Acta Hortic Sin,2015,42(4):655-664.DOI: 10.16420/j.issn.0513-353x.2014-1085.
[16]孙斌.盆栽金银花花期调控初步研究[J].山东林业科技,2015,45(6):41-43.SUN B.Preliminary research on the flowering regulation of the potted Lonicera japonica[J].J Shandong For Sci Technol,2015,45(6):41-43.DOI: 10.3969/j.issn.1002-2724.2015.06.010.
[17]温璐华,王真,庄维兵,等.外源GA4处理解除果梅花芽休眠的生理效应研究[J].中国南方果树,2015,44(5):16-22.WEN L H,WANG Z,ZHUANG W B,et al.Physiological effect of exogenous GA4 on the dormancy release of Japanese apricot flower buds[J].South China Fruits,2015,44(5):16-22.DOI: 10.13938/j.issn.1007-1431.20140474.
[18]FUKAZAWA J,OHASHI Y,TAKAHASHI R,et al.DELLA degradation by gibberellin promotes flowering via GAF1-TPR-dependent repression of floral repressors in Arabidopsis[J].Plant Cell,2021,33(7):2258-2272.DOI: 10.1093/plcell/koab102.
[19]潘瑞炽,董愚得.植物生理学[M].北京:高等教育出版社, 1998.PAN" R Z, DONG" Y D. Plant physiology[M]. Beijing: Higher Education Press,1998.
[20]KINET M J.Environmental,chemical,and genetic control of flowering[J].Hortic Rev,1993:279-334.
[21]肖安琪.三角梅花芽分化的内源激素变化与花期调控研究[D].广州:华南农业大学,2016.XIAO A Q.Study on the change of endogenous hormones during flower bud formation and regulation of blooming time for Bougainvillea[D].Guangzhou:South China Agricultural University,2016.
[22]MOURADOV A,CREMER F,COUPLAND G.Control of flowering time:interacting pathways as a basis for diversity[J].Plant Cell,2002,14(S1):111-130.DOI: 10.1105/tpc.001362.
[23]FORNARA F,DE MONTAIGU A,COUPLAND G.SnapShot:control of flowering in Arabidopsis[J].Cell,2010,141(3):550.DOI: 10.1016/j.cell.2010.04.024.
[24]KINOSHITA A,RICHTER R.Genetic and molecular basis of floral induction in Arabidopsis thaliana[J].J Exp Bot,2020,71(9):2490-2504.DOI: 10.1093/jxb/eraa057.
[25]孙丽,徐启江.高等植物开花诱导途径信号整合的分子机制[J].生物技术通讯,2009,20(6):885-890.SUN L,XU Q J.Molecular mechanisms of the signals integration of floral inductive pathways in high plants[J].Lett Biotechnol,2009,20(6):885-890.DOI: 10.3969/j.issn.1009-0002.2009.06.037.
[26]SHI X L,GUPTA S,RASHOTTE A M.Characterization of two tomato AP2/ERF genes,SlCRF1 and SlCRF2 in hormone and stress responses[J].Plant Cell Rep,2014,33(1):35-45.DOI: 10.1007/s00299-013-1510-6.
[27]BANTA J A,PURUGGANAN M D.The genetics and evolution of flowering time variation in plants:identifying genes that control a key life history transition[M]//FLATT T,HEYLAND A.Mechanisms of Life History Evolution.Oxford:Oxford University Press,2011:114-126.DOI: 10.1093/acprof:oso/9780199568765.003.0009.
[28]LEE H N,YOO S J,LEE J H,et al.Genetic framework for flowering-time regulation by ambient temperature-responsive miRNAs in Arabidopsis[J].Nucleic Acids Res,2010,38(9):3081-3093.DOI: 10.1093/nar/gkp1240.
[29]杨小凤,李小蒙,廖万金.植物开花时间的遗传调控通路研究进展[J].生物多样性,2021,29(6):825-842.YANG X F,LI X M,LIAO W J.Advances in the genetic regulating pathways of plant flowering time[J].Biodivers Sci,2021,29(6):825-842.DOI: 10.17520/biods.2020370.
(责任编辑 吴祝华)
基金项目:上海市科委重点研发项目(22dz1202200,20dz1203700);上海市绿化和市容管理局(林业局)科研专项(G200203)。
第一作者:陈香波(cxb7210@163.com),高级工程师。
*通信作者:张冬梅(zdm@shsyky.com),教授级高级工程师。
引文格式:陈香波,张冬梅,傅仁杰,等.白玉兰二次开花诱导与生理调控研究[J]. 南京林业大学学报(自然科学版),2024,48(5):97-104.
CHEN X" B,ZHANG D M, FU R J, et al.Induction and physiological regulatory mechanism of secondary flowering in Yulannia denudata[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(5):97-104.