幼龄栓皮栎叶面积预测模型建立及应用
2024-10-09李慧张婉常益豪杨霞肖祥伟朱景乐
摘要:【目的】建立幼龄栓皮栎叶面积预测模型,实现快速无损测量,并对43个栓皮栎家系的叶片形状变异情况和差异特征进行分析,为栓皮栎遗传多样性研究及良种选育提供理论基础。【方法】从全国不同地区筛选出43个栓皮栎家系作为研究对象,通过对叶长、叶宽、叶长宽比、叶长宽积与实测叶面积的相关性分析及曲线拟合分析,筛选出叶面积预测模型最佳参数;通过几何模型和曲线拟合分析,筛选最优的幼龄栓皮栎叶面积预测模型,并进行验证;运用统计描述、单因素方差分析、相关性分析、聚类分析的方法进行不同家系间叶片的差异分析。【结果】①叶长宽积(X1×X2)与幼龄栓皮栎叶面积呈显著相关,可根据这一组合指标进行幼龄栓皮栎叶面积预测模型的建立。②幼龄栓皮栎叶面积预测模型Y=0.595X1×X2+257.640最为精确,R2=0.946,标准误差低至32.830 cm2,可以用来预测幼龄栓皮栎叶片的面积。③43个栓皮栎家系间各项叶片指标均有不同程度的变异,不同家系间和家系内叶片差异均较大。④叶片性状与原产地地理信息相关性分析结果表明,除叶干质量与年降水量外,与其他指标均无显著相关性;43个家系不能独立分类,规律性不强。【结论】以叶长宽积为参数,建立了较为精准的幼龄栓皮栎叶面积预测模型Y=0.595X1×X2+257.640,为获得栓皮栎叶面积提供了一种高效、无损的方法,为栓皮栎良种培育和家系选择提供理论基础。
关键词:栓皮栎家系;叶面积预测模型;叶片形状;变异特征
中图分类号:S792"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)05-0246-09
Construction and application of the leaf area prediction model for young Quercus variabilis
LI Hui1,2, ZHANG Wan1,2, CHANG Yihao2, YANG Xia3, XIAO Xiangwei4, ZHU Jingle1
(1. Key Laboratory of Non-timber Forest Germplasm Enhancement amp; Utilization of National Forestry and Grassland Administration," Research Institute of Non-timber Forestry, Chinese Academy of Forestry," Zhengzhou" 450003, China; 2. College of Forestry, Henan Agriculture University, Zhengzhou 450002, China; 3. Nanwan State Forest Farm, Xinyang 464100, China; 4. Tianmushan State Forest Farm, Pingqiao District, Xinyang 464100, China)
Abstract: 【Objective】The leaf area prediction model of young Quercus variabilis was established to apply a rapid and nondestructive measurement. The leaf shape variation and difference characteristics of 43 different Q. variabilis families were analyzed to provide a theoretical basis for studying the genetic diversity and breeding of Q. variabilis. 【Method】A total of 43 Q. variabilis families were selected from different regions of China as the research objects. The optimal parameters of the leaf area prediction model were selected through correlation analysis and curve fitting analysis of leaf length, width, length-width ratio, length-width product, and measured leaf area.Through geometric model and curve fitting analysis, the optimal prediction model of the leaf area of young Q. variabilis was screened, and the results were verified. The differences in leaves among different families were analyzed by statistical description, one-way analysis of variance, correlation analysis, and cluster analysis. 【Result】(1) Leaf length-width product (X1×X2) was significantly correlated with the" leaf area of young Q. variabilis, and the prediction model of leaf area of young Q. variabilis could be established according to this combination index. (2) The leaf area prediction model of young Q. variabilis" Y=0.595X1 ×X2+257.640" was the most accurate, R2=0.946, and the standard error was as low as 32.830 cm2, which could be used to predict the leaf area of young Q. variabilis.(3) The leaf indexes of the 43 Q. variabilis families had different degrees of variation. The differences in leaves among different families and within families were large. (4) The results of the correlation analysis between leaf traits and geographical information of origin showed no significant correlation between other indexes except leaf dry weight and annual precipitation. 43 families could not be classified independently, and the regularity was not strong. 【Conclusion】A more accurate prediction model of the leaf area of young Q. variabilis Y=0.595X1×X2+257.640 was established by using leaf length and width product as parameters, which provided an efficient and nondestructive method for obtaining the leaf area of Q. variabilis and provided a theoretical basis for breeding and family selection of Q. variabilis.
Keywords:Quercus variabilis family; leaf area prediction model; leaf shape; variation characteristics
栓皮栎(Quercus variabilis)是壳斗科栎属乔木,是我国分布较为广泛的落叶阔叶树种之一,具有重要的观赏价值和经济价值。栓皮栎林分是森林景观的重要部分,在自然环境中发挥着维持生物多样性和环境保护作用。由于栓皮栎分布区域较广,分布区内环境复杂,所以不同地区的栓皮栎种群经过长期的自然选择和种群内的遗传分化,其中叶片的形状变异最为突出,形成了遗传结构不同的地理种群。
叶片是高等植物光合作用的主要器官,在植物生理功能中起着重要作用[1-2]。叶片大小对植物光能利用、水分蒸腾、生长和衰老、干物质积累等有着显著的影响,而叶面积作为植物功能性状的重要指标,已广泛应用于评价植物生态适应性、碳同化能力等多方面[3-6]。叶片表型性状直接影响植物体的一些生理活动及植物利用资源的能力,体现了植物为获得最大碳同化量所采取的生存适应策略[7-9]。叶片表型变异亦是植物遗传变异和环境互作的共同反映,由于叶片形态与植物营养[10]、生理[11]以及生态因子[12-13]等密切相关,因此具有较高的研究价值。
当前测量叶片面积的方法有:方格法、称量法、图像法、打孔称质量法、仪器法以及拟合法等,但这些方法都存在测定速度慢,效率低的情况。而通过构建叶面积预测模型能够在保证试验精度的前提下简化工作步骤,降低工作强度,显著提高科研效率,在植株无损状态下快速得到植物叶面积。这种测定方法的高效准确性在很多植物的叶面积测定中被验证,如西番莲属(Passiflora)[14]、杉木(Cunninghamia lanceolata)[15]、海南风吹楠(Horsfieldia hainanensis)[16]等。
参数的选择对叶面积预测模型的精度和使用便捷性有很大影响[17]。当前建立叶面积预测模型选用的预测指标主要有叶长、叶宽及组合指标等[18-19],但越来越多的研究结果表明,以叶片长宽积为自变量构建的叶面积线性模型能得到较高的拟合效果[20-22]。
本研究拟根据叶片形态和参数拟合的方法构建幼龄栓皮栎叶面积预测模型,并检验筛选出精度最高的模型,根据最优模型计算43个栓皮栎家系叶片面积,通过方差分析比较不同家系的差异显著性,最后通过聚类分析研究不同家系间叶片形态上的关系。研究有望建立幼龄栓皮栎叶面积预测模型,掌握栓皮栎家系间叶片遗传变异规律,为栓皮栎遗传多样性研究提供依据。
1 材料与方法
1.1 试验地概况
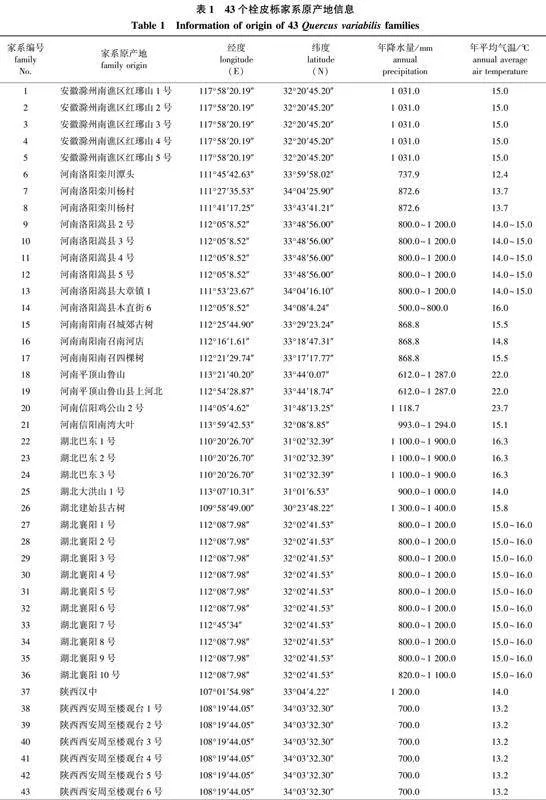
试验所用的43个栓皮栎家系叶片材料(表1)来自中国林业科学研究院经济林研究所信阳基地试验林,该试验林位于河南省信阳市平桥区(113°54′26″E,32°35′14″N)。该地区年平均气温15.1 ℃,1月平均气温1.9 ℃,7月平均气温28 ℃,年降水量900.0~1 200.0 mm,降水多集中在5—9月;年均无霜期221.4 d,年平均蒸发量1 662.2 mm,空气相对湿度74%,年平均日照2 026.7 h。从全国各地搜集到43个栓皮栎半同胞家系的种子,各家系原产地信息见表1。于2019年进行播种、育苗和定植,株行距2.0 m×1.0 m,2021年6月对各家系栓皮栎叶片进行采集,此时各栓皮栎家系树龄约2 a,平均株高0.8 m,平均地径1.1 cm。
1.2 试验方法
1.2.1 叶片面积测定及栓皮栎叶片预测模型的确定
从试验林中随机选择30个单株,每个单株选择5片叶片作为建立栓皮栎叶面积预测模型的材料,用叶面积仪分别精确测量叶片的叶面积;用游标卡尺测量150片叶片的叶长、叶宽,分别记作X1和 X2,并计算叶长宽积(X1×X2)和长宽比(X1/X2)两个组合指标,预测叶片面积记作Y。对叶长(X1)、叶宽(X2)先进行叶片相关指标与实际叶面积相关分析,以筛选叶片面积模型预测因子,再进行叶面积预测模型构建及检验[23-24]。具体步骤如下:
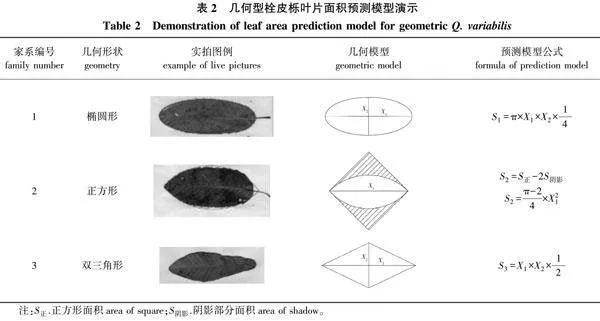
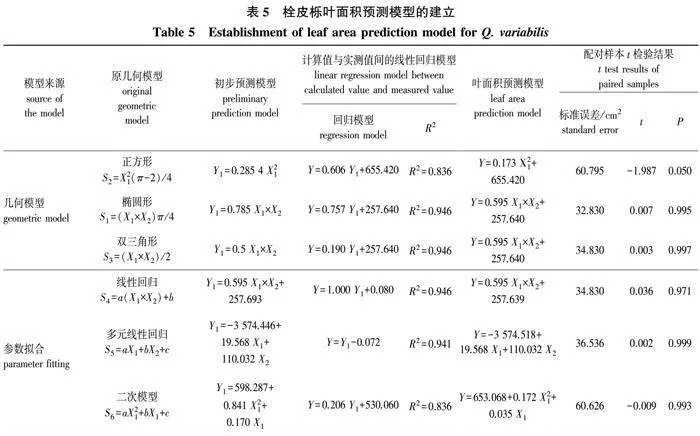
1)首先根据栓皮栎的叶片形状,选择椭圆形、正方形中两个半圆组合图形(后简称正方形)、双三角形3个几何模型(表2),用几何计算的方法,根据所需指标初步预测叶片面积,分别记作S1、S2、S3。同时再进行曲线拟合及多元线性回归的方法,在SPSS 26.0中分别以叶长、叶宽及叶长宽积为自变量,筛选其模型预测叶片面积的方法。
2)建立叶片实测面积与计算面积的线性回归模型,并根据模型计算经矫正后的叶片面积。
3)通过多配对样本t检验,检验矫正过的叶片面积与实测面积间的差异显著性,确定最终叶片面积预测模型。
1.2.2 43个家系叶片形状变异分析
采集试验林中43个栓皮栎家系的叶片,在每个家系选择15株(其中36号家系选择仅有的2个单株),每株在外部枝条中上部共采集5片叶子,带回实验室在80 ℃烘箱内烘干48 h,测定叶长、叶宽及叶干质量,并利用最精确的栓皮栎叶面积预测模型得到43个家系叶片的叶面积。计算每单株5片叶片的叶长、叶宽、叶干质量、叶面积的平均值作为该单株数值,通过家系15个单株的平均值、标准差的计算得到变异系数(CV),以衡量叶片性状家系内的变异水平[23],并通过单因素方差分析比较不同家系间的差异显著性。
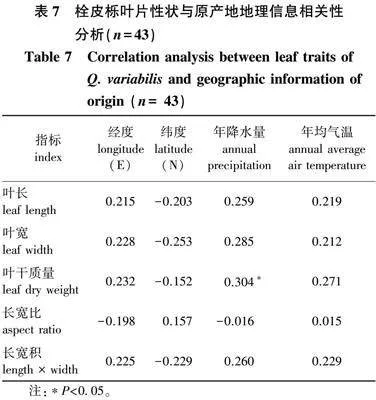
1.2.3 栓皮栎叶片性状与原产地地理信息相关性分析
以43个栓皮栎家系的叶片为研究对象,分别求得叶长、叶宽、叶干质量、叶长宽比、叶长宽积、叶面积的平均值,与各个家系原产地的经度、纬度、年降水量、年平均气温进行相关性分析。
1.2.4 43个栓皮栎家系叶片性状的聚类分析
以43个栓皮栎家系为对象,以叶长、叶宽、叶干质量、叶长宽比、叶长宽积及叶片面积为指标,进行聚类分析。
1.2.5 统计软件及操作
本研究中的相关性分析、曲线拟合及多元线性回归、单因素方差分析、描述统计及聚类分析等统计分析使用SPSS 26.0,线性回归图形绘制使用Excel 2019。
2 结果与分析
2.1 幼龄栓皮栎叶面积预测模型的建立与检验
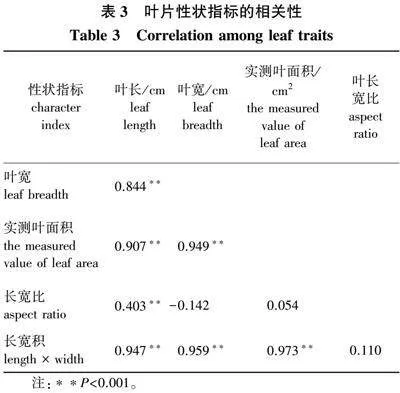
150片栓皮栎叶片的叶长、叶宽、叶长宽比与叶面积实测值间的相关性见表3,叶长、叶宽、叶长宽积与实测叶面积间均存在极显著相关关系,其相关关系分别为0.907、0.949和0.973,而实测叶面积和长宽比无显著相关关系。说明叶面积和叶长、叶宽关系非常紧密,且和叶片的组合指标间可能存在很强关联性。
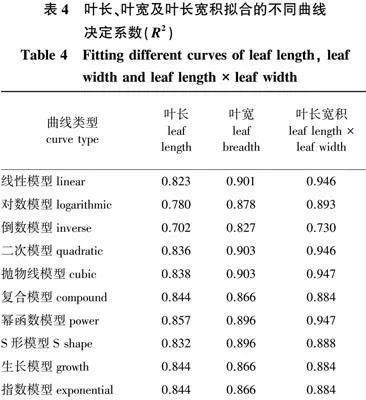
以叶长、叶宽及叶长宽积组合指标分别进行不同曲线拟合 (表4),结果显示:以叶长为自变量得到曲线的决定系数为0.702~0.857,其中幂函数模型拟合效果最好;以叶宽为自变量得到曲线的决定系数为0.827~0.901,其中线性模型拟合效果最好;以叶长宽积为自变量得到的曲线决定系数为0.730~0.947,其中幂函数模型和抛物线模型拟合效果最好。虽然以不同指标建立的回归曲线在拟合程度上有一定的差异,但组合指标应用整体效果优于单个指标。
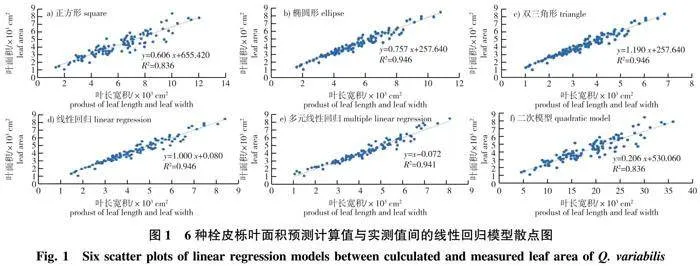
3种几何模型及3种曲线拟合方法得到的预测模型结果及预测准确性(图1、表5),6个模型决定系数为0.836~0.946,线性关系非常明显,说明拟合效果较好。为进一步对比拟合方程的准确性,通过多配对样本t检验,检验矫正过的叶片面积与实测面积间的差异显著性,结果表明,除正方形预测模型外,其余5种模型的叶面积与实测值间均无显著性差异(P>0.05且接近于1),且标准误低至32.830,使用这5种模型进行叶片面积的预测均较为可靠。其中,椭圆形、双三角形、线性回归3种方法得到的叶面积模型几近相同,为Y=0.595X1×X2+257.640。因此,采用测定叶长宽积(X1×X2)从而预测叶片面积是可行并可靠的。
2.2 43个栓皮栎家系叶片形状变异分析
43个家系共632株栓皮栎的叶长变异幅度为5.38~17.92 cm,变异系数为17.34%;叶片宽的变异幅度为1.62~8.74 cm,变异系数为20.12%;叶片干质量的变异幅度为0.06~0.79 g,变异系数为33.94%;叶片面积变异幅度为8.34~61.05 cm2,变异系数为30.86%。
43个家系中叶长最大的为36号家系,平均叶长为13.91 cm;最小的为41号家系,平均叶长为7.23 cm;家系内叶长的变异系数为8.90%~27.98%。
43个家系中叶宽最大的为36号家系,平均叶宽为4.46 cm;最小的为41号家系,平均叶宽为2.34 cm;家系内叶宽的变异系数为8.16%~41.54%。
43个家系中干质量最大的为20号家系,干质量为0.39 g;最小的为41号家系,干质量为0.15 g;家系内干质量的变异系数为12.84%~72.05%。
43个家系中叶面积最大的家系为36号家系,叶面积为39.85 cm2;最小的为41号家系,叶面积为12.84 cm2;家系内叶面积的变异系数为12.74%~57.77%。
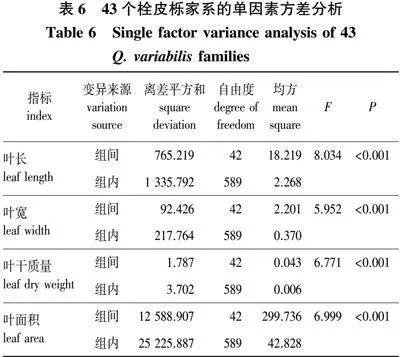
由表6的单因素方差分析可知,叶长、叶宽、叶干质量及叶面积这些指标在43个家系中均存在极显著差异(P<0.001),说明不同家系间的叶片性状差别非常明显。
2.3 43个栓皮栎家系叶片性状与原产地地理信息相关性分析
由叶片性状与原产地地理信息相关分析结果(表7)可知,经度、纬度、年平均气温与叶长、叶宽、叶干质量、长宽积、长宽比、叶面积之间均没有相关关系,年降水量除与叶片干质量之间有显著正向相关关系外,与叶片的其他指标无显著的相关关系。即随着年降水量的增长,叶片干质量也随之增长。
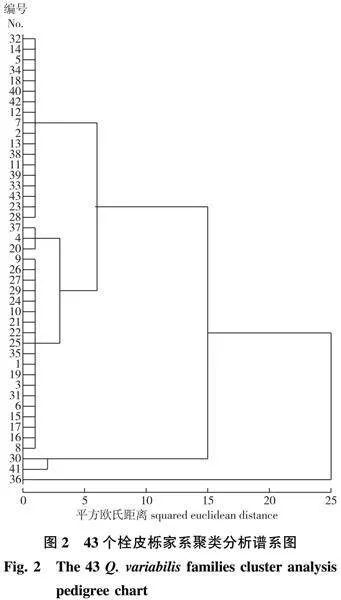
2.4 43个栓皮栎家系聚类分析
43个家系的聚类分析(图2)显示,不同来源的栓皮栎家系聚类没有规律而言,说明可能存在以下原因:①样本数量不够,无法为聚类分析提供足够的数据;②可能是只选择叶片性状来聚类分析是不行的,应该增加更多的指标,如果实特点、物候期等;③叶片性状本身变异就比较大,而且家系内部变异及种源内家系间变异幅度较大,所以无法区分开不同家系。
3 结论与讨论
3.1 结论
1)栓皮栎的叶长、叶宽及长宽积与叶面积间具有显著的关联性,长宽比与叶面积间无显著相关性;用叶长、叶宽及长宽积拟合不同曲线,不同指标、不同预测模型间有一定差异,但组合指标整体优于单个指标。
2)配对样本t检验的结果说明,直接根据正方形预测模型得到的叶面积与实测面积间可能存在显著差异(P=0.05),其余5种模型的预测值与实测值间均无显著性差异(P>0.05且接近于1),且标准误差低至32.830 cm2, 说明使用该5种模型进行叶片面积的预测均较为可靠,具有生物学意义和统计学意义。
3)在所得5种叶面积预测模型中,椭圆形、双三角形、线性回归3种方法所得模型相近,为Y=0.595 X1×X2+257.640。可作为幼龄栓皮栎叶面积模型,以简化测量工作难度,降低工作量。
4)对43个栓皮栎家系叶片指标的差异性分析发现栓皮栎不同家系间差异较大,同时家系内也存在较大差异。叶长、叶宽、叶干质量及叶面积这些指标在43个家系中均存在极显著差异,说明不同家系间的叶片性状差别非常明显。
5)叶片性状与原产地地理信息相关性分析显示,叶干质量与年降水量存在显著关系,而其他指标之间均无显著相关关系。
6)聚类分析时,不同产地的栓皮栎不能聚在一起,没有规律。建议增加测定指标,可结合果实特点、物候期等多个指标再进行不同产地间的对比。
3.2 讨论
1)在植物叶面积预测模型的建立中,叶片指标的选择直接影响着最终结果的准确性[25]。许多研究表明,在常用的叶宽、叶长、叶长宽积等参数中,以长宽积建立预测模型拟合度和准确度较高[26]。虽然如此,不同家系间幼龄栓皮栎叶片性状变异大,且关于幼龄栓皮栎叶面积模型建立时参数的选择方面鲜有报道,仍需对不同指标进行分析验证。结果表明,幼龄栓皮栎实测叶面积与长宽积(X1×X2)这一组合指标相关性显著,所得模型准确、可靠。
2)叶片作为植物进行生理生化活动的重要场所,了解其外部性状指标对探究植物生长状态有重要的作用[27]。通过叶面积预测模型的建立,能够在无损状态下同样实现对栓皮栎生长状态的评估。叶面积预测模型的计算方法除了曲线拟合,还包括几何模型的建立[28-29]。本试验通过参数选择、相关性分析、拟合度比较、t检验等方法,初步筛选出5种较为准确的叶面积模型,其中椭圆形、双三角形、线性回归3种方法得到的模型相近,为Y=0.595 X1×X2+257.640。这充分证明了最终选用的预测模型的准确性和可靠性,同时也进一步说明,相比其他指标,叶长宽积的组合指标与幼龄栓皮栎叶面积的关联更为密切。
3)43个栓皮栎家系间叶片性状差异较大,家系内也存在较大差异。这个结果说明不同栓皮栎家系在积极响应和适应环境,同时丰富的变异情况有利于栓皮栎种群适应环境[30-31]。虽然不同栓皮栎家系间和各家系内叶片性状有较大差异,但试验得到的叶面积预测模型具有普遍性和较高的准确性,叶面积预测模型的建立对于了解幼龄栓皮栎的生物量等信息有重要作用。
4)叶片性状与原产地地理信息相关性分析显示,除叶干质量与年降水量外,其他指标之间均无显著相关关系,说明他们之间并无明显的规律。聚类分析结果也表明无法将不同来源地的家系聚在一起,没有明显规律。考虑到栓皮栎在环境中具有较强的适应性,叶片会对环境因素作出积极响应[30-33],可以增加试验样品数量,丰富测试指标,使聚类结果更加准确。
参考文献(reference):
[1]李娟霞,田青.兰州市6种园林植物叶片形态和光合生理特征[J].西北农林科技大学学报(自然科学版),2022,50(1):72-80.LI J X,TIAN Q.Leaf morphology and photosynthetic physiological characteristics of six garden plants in Lanzhou[J].J Northwest A & F Univ (Nat Sci Ed),2022,50(1):72-80.DOI:10.13207/j.cnki.jnwafu.2022.01.009.
[2]魏龙鑫,章异平,李艺杰,等.栓皮栎叶片和枝条非结构性碳水化合物调配关系研究[J].南京林业大学学报(自然科学版),2021,45(2):96-102.WEI L X,ZHANG Y P,LI Y J,et al.Allocation of non-structural carbohydrates (NSC) contents in leaves and branches of Quercus variabilis during its growth process[J].J Nanjing For Univ (Nat Sci Ed),2021,45(2):96-102.DOI: 10.12302/j.issn.1000-2006.201909029.
[3]JAUSORO V,LLORENTE B E,APSTOLO N M.Structural differences between hyperhydric and normal in vitro shoots of Handroanthus impetiginosus (Mart.)[J].Plant Cell Tiss Organ Cult,2010,101(2):183-191.DOI:10.1007/s11240-010-9675-y.
[4]BAJWA G A,UMAIR M,NAWAB Y,et al.Morphometry of leaf and shoot variables to assess aboveground biomass structure and carbon sequestration by different varieties of white mulberry (Morus alba L.)[J].J For Res,2021,32(6):2291-2300.DOI:10.1007/s11676-020-01268-7.
[5]WONG C Y S,GAMON J A.The photochemical reflectance index provides an optical indicator of spring photosynthetic activation in evergreen conifers[J].New Phytol,2015,206(1):196-208.DOI:10.1111/nph.13251.
[6]薛雪,张金池,孙永涛,等.上海常绿树种固碳释氧和降温增湿效益研究[J].南京林业大学学报(自然科学版),2016,40(3):81-86.XUE X,ZHANG J C,SUN Y T,et al.Study of carbon seqestration amp; oxygen release and cooling amp; humidifying effect of main greening tree species in Shanghai[J].J Nanjing For Univ (Nat Sci Ed),2016,40(3):81-86.DOI:10.3969/j.issn.1000-2006.2016.03.014.
[7]孙文勇,肖小华,王玉芳,等.十堰市引种栽培油橄榄品种的叶片与果实表型性状分析[J].湖北林业科技,2022,51(3):10-13.SUN W Y,XIAO X H,WANG Y F,et al.Analysis on leaf and fruit phenotypic characters of Olea europea varieties introduced and cultivated in Shiyan[J].Hubei For Sci Technol,2022,51(3):10-13. DOI:10.3969/j.issn.1004-3020.2022.03.004.
[8]罗芊芊,周志春,邓宗付,等.南方红豆杉天然居群叶片的表型性状和氮磷化学计量特征的变异规律[J].植物资源与环境学报,2021,30(1):27-35.LUO Q Q,ZHOU Z C,DENG Z F,et al.Variation law of phenotypic traits and nitrogen and phosphorus stoichiometric characteristics of leaf of natural populations of Taxus wallichiana var. mairei[J].J Plant Resour Environ,2021,30(1):27-35.DOI:10.3969/j.issn.1674-7895.2021.01.04.
[9]MCCULLOH K A,SPERRY J S.Patterns in hydraulic architecture and their implications for transport efficiency[J].Tree Physiol,2005,25(3):257-267.DOI:10.1093/treephys/25.3.257.
[10]于秋香,刘警,李扬,等.核桃种质资源叶片表型性状的遗传多样性研究[J].农业科学与技术:英文版,2021,22(1):1-8.YU Q X,LIU J,LI Y,et al.Genetic diversity of leaf phenotypic traits in walnut germplasm resources[J].Agric Sci & Technol,2021,22(1):1-8.DOI:10.16175/j.cnki.1009-4229.2021.01.001.
[11]仲磊,张焕朝,范俊俊,等.夏季淹水胁迫对北美枫香苗木叶色及光合荧光特性的影响[J].南京林业大学学报(自然科学版),2021,45(2):69-76.ZHONG L,ZHANG H C,FAN J J, et al.Effects of flooding stress on leaf color and photosynthetic fluorescence characteristics of Liquidambar styraciflua cutting seedlings in summer[J].J Nanjing For Univ (Nat Sci Ed),2021,45(2):69-76.DOI:10.12302/j.issn.1000-2006.201909045.
[12]GUILHERME PEREIRA C,DES MARAIS D L.The genetic basis of plant functional traits and the evolution of plant-environment interactions[J].Int J Plant Sci,2020,181(1):56-74.DOI:10.1086/706190.
[13]WRIGHT I J,DONG N,MAIRE V,et al.Global climatic drivers of leaf size[J].Science,2017,357(6354):917-921.DOI:10.1126/science.aal4760.
[14]吴凤婵,李安定,蔡国俊,等.6种西番莲属(Passiflora)植物叶面积经验模型构建[J].果树学报,2021,38(9):1600-1610.WU F C, LI A D, CAI G J, et al.Construction of empirical models for leaf area estimation in six Passiflora species[J].J Fruit Sci,2021,38(9):1600-1610.DOI:10.13925/j.cnki.gsxb.20210018.
[15]彭曦,闫文德,王光军,等.杉木叶形态特征与叶面积估算模型[J].生态学报,2018,38(10):3569-3580.PENG X,YAN W D,WANG G J,et al.Leaf morphological characteristics and leaf area estimation model for Cunninghamia lanceolata[J].Acta Ecol Sin,2018,38(10):3569-3580.DOI:10.5846/stxb201704130660.
[16]杜尚嘉,王鑫,吴海霞,等.海南风吹楠成熟叶叶面积估算模型建立[J].热带林业,2021,49(4):17-21.DU S J,WANG X,WU H X,et al.Developing models to estimate the area of mature leaves of Horsfieldia hainanensis[J].Trop For,2021,49(4):17-21.DOI:10.3969/j.issn.1672-0938.2021.04.004.
[17]巫娟,胡姝珍,茅思雨,等.基于叶片形态的毛竹单叶叶面积模型[J].林业科学,2020,56(8):47-54.WU J,HU S Z,MAO S Y,et al.Single leaf area model of Phyllostachys edulis based on leaf morphology[J].Sci Silvae Sin,2020,56(8):47-54.DOI:10.11707/j.1001-7488.20200806.
[18]王进杰,杨军,胡姝珍,等.3种丛生竹叶面积预测模型研究[J].南方林业科学,2020,48(1):7-12.WANG J J,YANG J,HU S Z,et al.Study on leaf area predicting model of three sympodial bamboo species[J].South China For Sci,2020,48(1):7-12.DOI:10.16259/j.cnki.36-1342/s.2020.01.002.
[19]谈丽华,巫娟,胡妹珍,等.基于叶形分类的淡竹叶面积预测模型研究[J].世界竹藤通讯,2020,18(1):6-10.TAN L H,WU J,HU M Z,et al.Leaf area predicting model for Phyllostachys glauca based on leaf morphology classification[J].World Bamboo Rattan,2020,18(1):6-10.DOI:CNKI:SUN:ZTTX.0.2020-01-004.
[20]YU X J,SHI P J,SCHRADER J,et al.Nondestructive estimation of leaf area for 15 species of vines with different leaf shapes[J].Am J Bot,2020,107(11):1481-1490.DOI:10.1002/ajb2.1560.
[21]SCHRADER J,SHI P J,ROYER D L,et al.Leaf size estimation based on leaf length,width and shape[J].Ann Bot,2021,128(4):395-406.DOI:10.1093/aob/mcab078.
[22]CAI H Y,DI X Y,JIN G Z.Allometric models for leaf area and leaf mass predictions across different growing seasons of elm tree (Ulmus japonica)[J].J For Res,2017,28(5):975-982.DOI:10.1007/s11676-017-0377-8.
[23]张鹏,杨颖,奚如春,等.高州油茶种群叶片性状变异分析[J].林业与环境科学,2018,34(5):13-19.ZHANG P,YANG Y,XI R C,et al.Leaf phenotypic variation in populations of Camellia gauchowensis[J].For Environ Sci,2018,34(5):13-19.DOI: 10.3969/j.issn.1006-4427.2018.05.003.
[24]YU K, REDDY G V P, SCHRADER J, et al. A nondestructive method of calculating the wing area of insects[J]. Ecology and Evolution, 2022,12(4):e8792. DOI: 10.1002/ece3.8792.
[25]TEOBALDELLI M, ROUPHAEL Y, FASCELLA G, et al. Developing an accurate and fast non-destructive single leaf area model for Loquat (Eriobotrya japonica Lindl) cultivars[J]. Plants,2019,8(230):1-12. DOI:10.3390/plants8070230.
[26]DING X T, YU L Y, JIANG Y P, et al. Changes in leaf length, width, area, and photosynthesis of fruit cucumber in a greenhouse production system[J]. Hort Science,2020,55(7):1-5. DOI:10.21273/HORTSCI14637-19.
[27]PEREIRA C G, MARAIS D L. Special issue: functional trait evolution: the genetic basis of plant functional traits and the evolution of plant-environment interactions[J]. International Journal of Plant Sciences,2020,181(1). DOI:10.1086/706190.
[28]杨红云,路艳,孙爱珍,等. 水稻叶片几何参数无损测量方法研究[J]. 江西农业大学学报,2020,42(2):407-418. YANG H Y, LU Y, SUN A Z, et al. Non-destructive measurement of rice leaf geometric parameters[J]. Acta Agriculturae Universitatis Jiangxiensis,2020,42(2):407-418. DOI:CNKI:SUN:JXND.0.2020-02-024.
[29]LI Y Q, ZOU D T, SHRESTHA N, et al. Spatiotemporal variation in leaf size and shape in response to climate[J]. Journal of Plant Ecology,2020,13(1):87-96. DOI:10.1093/jpe/rtz053.
[30]SHANG J,JIANG B, ZHENG Y H,et al. Responses of leaf functional traits of Torreya fargesii to the altitudinal variation[J].Agricultural Science amp; Technology,2017,18(11):2179-2184. DOI:10.16175/j.cnki.1009-4229.2017.11.037.
[31]李理渊,李俊,同小娟,等.不同光环境下栓皮栎和刺槐叶片光合光响应模拟[J].应用生态学报,2018,29(7):2295-2306. LI L Y, LI J, TONG X J, et al. Simulation on photosynthetic light-responses of leaves of Quercus variabilis and Robinia pseudoacacia under different light conditions[J].Chinese Journal of Applied Ecology,2018,29(7):2295-2306. DOI:10.13287/j.1001-9332.201807.029.
[32]WANG Z H, ZHENG R, YANG L L,et al. Elevation gradient distribution of indices of tree population in a montane forest: the role of leaf traits and the environment[J].Forest Ecosystems,2022,9(1):124-130. DOI:10.1016/J.FECS.2022.100012.
[33]HANG T, LU N, TAKAGAKI M, et al. Leaf area model based on thermal effectiveness and photosynthetically active radiation in lettuce grown in mini-plant factories under different light cycles[J]. Scientia Horticulturae,2019:252. DOI: 10.1016/j.scienta.2019.03.057.
(责任编辑 吴祝华)
基金项目:河南省重点研发专项(241111113300);中央级公益性科研院所基本科研业务费专项资金项目(CAFYBB2018ZB001-11)。
第一作者: 李慧(lh382490531@163.com)。
*通信作者:朱景乐(zhujingle1982@126.com),副研究员。
引文格式:李慧,张婉,常益豪,等. 幼龄栓皮栎叶面积预测模型建立及应用[J]. 南京林业大学学报(自然科学版),2024,48(5):246-254.
LI H, ZHANG W, CHANG Y H, et al. Construction and application of the leaf area prediction model for young Quercus variabilis[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(5):246-254.