海拔对塔城地区野果林生态位与种间关联影响
2024-07-10陆鑫,巴乐金,乌兰·吾尚,刘雪冰,夏依达·艾力,巴音达拉
陆鑫,巴乐金,乌兰·吾尚,刘雪冰,夏依达·艾力,巴音达拉

摘要:新疆塔城地区野果林作为新疆野果林的重要组成部分,在维持当地物种资源多样性中起到至关重要的作用,为探索该地区野果林生境因子对其生态位与种间关联的影响,在本地区的额敏县、裕民县和托里县,设立30 m×30 m的样方80个,记录海拔、坡度、坡向、土壤含水量、pH和草本盖度6种生境因子参数,通过Mantel检验分析生境因子对物种分布的影响,并将影响最大的生境因子分成4级,采用Levins生态位宽度指数(Oik)、Spearman秩关联系数(rik)、Shannon-wiener指数和Simpson指数等进行分析。结果表明,1)在6种生境因子中,海拔因子对树种分布影响最大(Mantel指数为0.261 5),且呈现极显著性(P=0.000 1);其次是土壤含水量、草本盖度(Mantel指数分别为0.109 2、0.098 7)。2)塔城地区野果林主要树种为新疆野苹果(Malus sieversii)、红果山楂(Crataegus pinnatifida)和大叶绣线菊(Spiraea salicifolia),主要分布在海拔1 100~1 300 m。3)新疆野苹果的频度和生态位宽度在各个等级均最高,而其余树种在不同等级下却表现出较大的差异,随着海拔的增加,树种的频度与生态位宽度出现了先增长后减少的趋势。4)在4个海拔(第1等级为700~900 m,第2等级为900~1 100 m,第3等级为1 100~1 300 m,第4等级为1 300~1 500 m)下,Oik>0分别占比树种间生态位重叠的83.33%、65.38%、60.44%和76.19%;生态位重叠指数随着海拔的增加呈现出先减少后增加的趋势。5)4个海拔下,rik≥0分别占比相对应海拔树种间正关联性66.67%、58.97%、35.16%和38.10%;树种在第1和第2等级下表现为以正关联性为主,而第3、第4等级则以负关联性为主,同时,正关联性、显著正关联性、显著负关联性随着海拔的增加呈现出先减少后增加的趋势,而无明显关联性却表现出先增加后减少的趋势。研究显示,海拔是塔城地区野果林物种分布最主要的影响因子,随着海拔增加,树种频度与生态位宽度出现了先增长后减少的趋势,生态位重叠指数、种间关联性呈现出先减少后增加的趋势,在海拔为1 100~1 300 m时,树种有着更好的分布与种间关系。
关键词:野果林;海拔;群落格局;样地调查;生境因子;种间关联性
中图分类号:718.54文献标识码:A文章编号:1006-8023(2024)03-0076-12
The Impact of Elevation on the Ecological Niche and Interspecific Association of Wild Fruit Forests in Tacheng Area
LU Xin1, BALEJIN2, WULAN·Wushang1 , LIU Xuebing1, XIAYIDA·Aili1, BAYANDALA1*
(1.College of Forestry and Landscape Architecture, Xinjiang Agricultural University, Urumqi 830052, China; 2.Middle School of Unity Farm, Ninth Agricultural Division, Xinjiang Production and Construction Corps, Emin 834601, China)
Abstract:As an important component of the wild fruit forest in Tacheng, Xinjiang, the wild fruit forest plays a crucial role in maintaining local species diversity. To explore the impact of habitat factors on the ecological niche and interspecific relationships of wild fruit forests in this region, 80 plots of 30×30 meters were established in Emin County, Yumin County, and Toli Count, recording six habitat factor parameters: altitude, slope, aspect, soil moisture content, pH, and herbaceous coverage. The impact of habitat factors on species distribution was analyzed through Mantel test, and the most influential habitat factors were divided into 4 levels. Levins niche width index, Spearman rank correlation number (rik), Shannon-wiener index, and Simpson index were used for analysis. The results showed that, 1) among the six habitat factors, altitude had the greatest impact on the distribution of tree species (Mantel index was 0.261 5), and showed extremely significant (P=0.000 1); next were soil moisture content and herbaceous coverage (Mantel indices of 0.109 2 and 0.098 7, respectively). 2) The main tree species in the wild fruit forest of Tacheng area were Malus sieversii, Crataegus pinnatifida, and Spiraea salicifolia, which were mainly distributed at an altitude of 1 100-1 300 meters. 3) The frequency and niche width of Malus sieversii were the highest at all levels, while other tree species showed significant differences at different levels. With the increase of altitude, the frequency and niche width of tree species showed a trend of first increasing and then decreasing. 4) At four altitudes(the first level is 700-900 meters, the second level is 900-1100 meters, the third level is 1100-1300 meters, and the fourth level is 1300-1500 meters), Oik>0 accounted for 83.33%, 65.38%, 60.44% , and 76.19% of the ecological niche overlap among tree species, respectively; the ecological niche overlap index showed a trend of first decreasing and then increasing with the increase of altitude. 5) At four altitudes, rik≥0 accounted for 66.67%, 58.97%, 35.16%, and 38.10% of the positive correlations among tree species at corresponding altitudes; tree species exhibited a predominantly positive correlation in the first and second levels, while negative correlation was predominant in the third and fourth levels. At the same time, positive correlation, significant positive correlation, and significant negative correlation showed a trend of first decreasing and then increasing with increasing altitude, while no significant correlation showed a trend of first increasing and then decreasing. The results indicated that, altitude was the main influencing factor on the distribution of wild fruit forest species in the Tacheng area, as the altitude increased, the frequency of tree species and niche width showed a trend of first increasing and then decreasing, while the niche overlap index and inter species correlation showed a trend of first decreasing and then increasing, at an altitude of 1 100 to 1 300 meters, tree species had better distribution and interspecific relationships.
Keywords:Wild fruit forest; elevation; community pattern; sample site survey; habitat factor; species association
0引言
森林在维护生物多样性和碳汇方面发挥着核心作用,有助于缓解气候变化[1-3],但特定的环境因素可能对森林的组成和发育起到重要影响,并可能沿一定的梯度表现出显著的差异性[4-6]。大量的研究表明,生态系统功能随纬度的增加而下降,与生物多样性的大尺度格局是一致的[7-8]。相比之下,生态系统功能与海拔梯度之间的关系研究较少,海拔梯度的变化会影响温度、湿度和土壤等多种环境因子,而这些环境因子在海拔梯度的变化要比纬度梯度上快1 000倍[9],这种快速且相对明显的变化对研究环境因子对物种的关系、多样性影响具有重要意义。
植物群落内不同的物种之间具有相互促进和竞争等多种关系,这些关系主要体现在物种的生态位和种间关联上[10]。种间关联是由于群落生境的差异导致物种分布差异而引起的,因此研究种间关联可以揭示物种之间的相互作用,判断物种是否存在竞争关系,预测群落演替,有助于正确认识植物群落的特征。近年来,种间关联在研究中得到广泛应用,对于指导濒危物种的保护和保育起到了积极作用[11-17]。因此,揭示群落内物种之间的关系,是了解物种组合规律及其多样性维持机制的有效手段,对于了解种群在群落中的地位、种群间的相互关系、植被资源的保护和可持续利用等具有重要意义[18-20]。
新疆野果林作为新疆特殊的生物多样性区域之一,具有重要的保护价值,是珍贵的野生果树起源地和遗传基因库,分布于伊犁河谷和塔城地区[21]。然而,近年来全球气候变暖、旅游业快速发展等多种因素导致野果林面积急速减少。新疆野果林的主要组成物种新疆野苹果(Malus sieversii)已面临大面积死亡和更新困难问题,已被列为中国优先保护物种名录和国家重点保护植物[22]。目前,对新疆野果林的研究主要集中在资源调查、遗传多样性、病虫害防治和栽培起源等方面,但对于新疆野果林的种群分布报道较少,且关于塔城地区管辖内野果林种间关系及关联性方面的研究尚未报道[23-27]。因此,本研究以塔城地区野果林为对象,通过建立样地和样方进行调查,获得群落基本数据,对不同生境因子等级下树种频度、生态位宽度、种间关联性和多样性等进行分析,旨在为维持新疆野果林群落的稳定性、分析放牧对野果林的影响、保护和管理珍稀野生果树资源提供科学依据。
1研究区概况
塔城地区野果林作为新疆野果林重要组成部分,主要分布在额敏县、裕民县和托里县,除以上区域外没有分布。其中额敏野果林是塔城地区面积最大的野果林,主要坐落于额敏县霍吉尔特蒙古民族乡以东13 km处,地处吾尔喀夏尔山南麓西部,地理坐标为83°59′ E,46°22′ N,海拔为900~1 500 m;裕民县野果林分布区位于82°30′ E~83°30′ E,45°51′ N~47°04′ N,海拔为800~1 300 m;托里县野果林位于托里县以北20 km,准噶尔盆地西侧,塔额盆地边缘,分布在老风口林场境内,地理坐标为83°33′ E,46°08′ N,年降水量为246.9 mm。
2研究方法
2.1样地调查
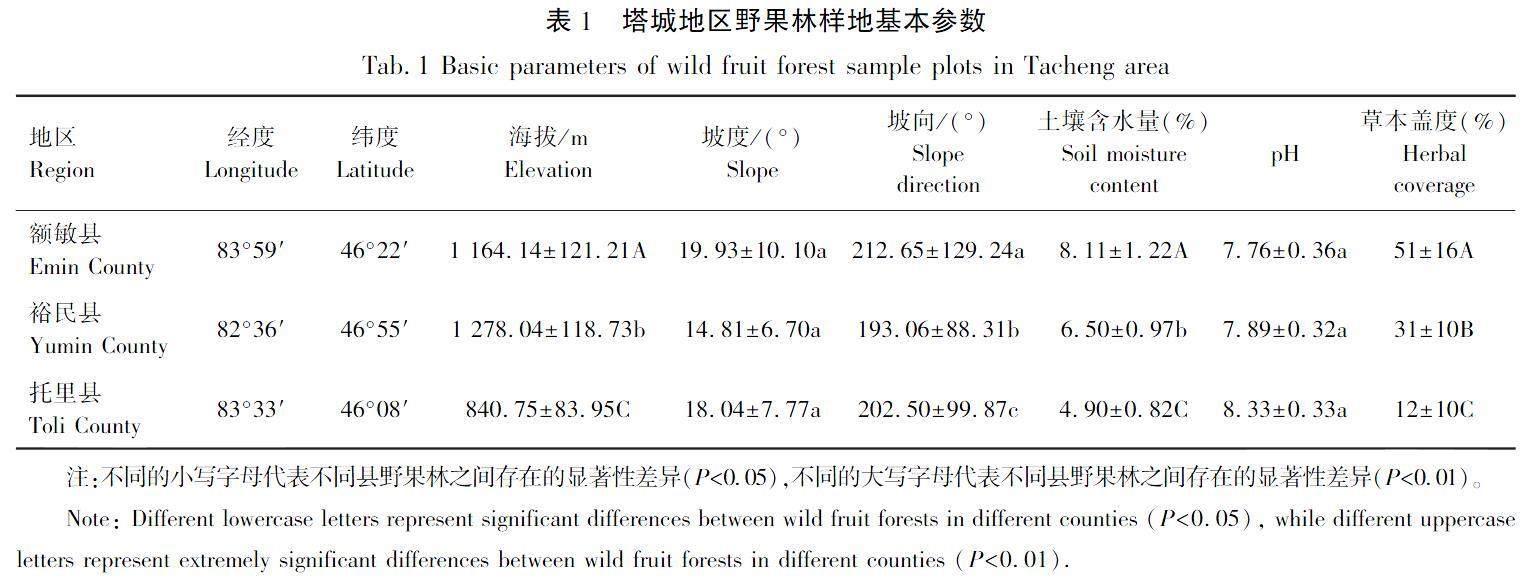
2022年6—7月对塔城地区额敏县、裕民县和托里县的野果林进行样地调查,涵盖山脊、山谷、山坡和河流等常见地形地貌,其总样地为80个,其中额敏县40个,裕民县16个,托里县24个,样方大小为30 m×30 m,总计72 000 m2,并在样方中心架设全站仪,记录样方的海拔、坡度、坡向、每个乔木、灌木的坐标位置、基径、高度和冠幅等参数。草本盖度采用5点法,即在样方4个角点和中心点对草本盖度进行测量并计算平均值;坡向采取正北方向为0的方式,沿东、南、西方向依次增加,范围为0°~360°;土壤含水量及pH使用TZS-3D土壤含水量/pH/光照度测定仪(浙江托普云农科技股份有限公司,中国)进行测定,并将每个生境因子下野果林物种数量,分成4个等级进行生态位分析。样地应选择群落内物种的组成、群落的结构以及生境要相对均匀的地方,样地之间尽量保证300 m以上的间距[28]。样地基本参数,见表1。
2.2研究方法
2.2.1Mantel检验
使用Mantel检验,控制环境距离矩阵度量地理距离与物种组成的关联性[29],公式如下。
B(j,k)=∑ni=1Xij-Xik∑ni=1Xij+Xik。(1)
式中:B(j,k)为第j个和第k个样方间的Bray-Curtis距离系数;n为物种数;Xij、Xik分别为第j个和第k个样方的第i种的多度数据。
2.2.2生态位宽度
采用Levins生态位宽度指数,确定一个群落中所利用的各种不同资源的总和。
Bi=1∑rj=1(Pij)2。(2)
式中:Bi为物种 i 的生态位宽度;Pij为物种 i在给定地点j 中的占比;r 为样地数量。
2.2.3生态位重叠
采用Levins生态位重叠指数,表达在群落中物种间的生态位重叠的程度。
Oik=∑rj=1Pij-Pkj∑rj=1(Pij)2。(3)
式中:Oik 表示物种 i与物种 k 的生态位重叠值;Pij 和 Pkj 分别表示物种 i 和 k 在资源位 j 上的重要值占所有资源位上的重要值总和的比例。
2.2.4种间关联性
Spearman秩关联系数可以反映出物种关系的指标[30],其公式如下
rik=1-6∑Nj=1(xij-xkj)2N3-N。(4)
式中:rik为物种i与物种k的种间关联性;N代表总样本数量;xij和xkj分别为物种i 和物种 k在样方中j 中的重要值。
2.3数据处理
本研究使用R语言(R 4.3.0版本)完成数据分析与作图。运用R语言中Species Association Analysis (SPAA)程序包完成的生态位宽度、生态位重叠和种间关联性分析,制图中运用了ggplot2、corrplot程序包[31]。
3结果与分析
3.1Mantel统计量
通过表2可知,塔城地区野果林综合生境因子对群落物种变化呈极显著性(P=0.001 3),Mantel指数为0.139 8,同时随着综合生境因子百分数的增加,表明总体生境因子对塔城地区野果林物种变化有着极为显著的影响。在对各个生境因子进行分析时发现,在6种生境因子中,海拔对树种变化影响最大,Mantel指数为0.261 5,且呈现极显著性(P=0.000 1);坡向因子对物种变化影响的Mantel指数为-0.009 5,但不显著(P=0.351 9);海拔、坡度、土壤pH、土壤含水量、草本盖度对物种变化影响的Mantel指数均为正值,但坡度(P=0.412 1)与土壤pH(P=0.196 3)却表现出不显著;结合Mantel指数与显著性水平可以得出,在塔城地区野果林中对群落物种分布影响最重要的依次是海拔、土壤含水量和草本盖度。为更好地展示不同海拔对塔城地区野果林的影响,将塔城地区野果林海拔范围分成4个等级,第1等级为700~900 m,第2等级为900~1 100 m,第3等级为1 100~1 300 m,第4等级为1 300~1 500 m。
由图1可知,塔城地区野果林主要树种为新疆野苹果(Malus sieversii)、红果山楂(Crataegus sanguinea)、大叶绣线菊(Spiraea salicifolia),同时,新疆野苹果、疏花蔷薇(Rosa multiflora)和小叶忍冬(Lonicera microphylla)等树种的丰度总体上随着海拔的增加呈现出增加的趋势;而大叶绣线菊、疣枝桦和野扁桃等树种的丰度总体上随着海拔的增加呈现出减少的趋势;黑果小檗(Berberis atrocarpa)、密叶杨(Populus talassica)、欧洲稠李(Padus racemosa)、野扁桃(Prunus tenella)、白柳(Salix alba)、欧亚圆柏(Juniperus sabina)和黑果栒子(Cotoneaster melanocarpus)等树种的丰度总体上与海拔的增加呈现出极小的变化或不相关;同时,绝大多数树种的主要分布在海拔1 100~1 300 m(第3等级)。
3.2频度与生态位宽度
由表3可知,在塔城地区野果林4个海拔等级下,新疆野苹果、红果山楂和大叶绣线菊均有分布。新疆野苹果的频度和生态位宽度在各个等级都最高,优势度与利用资源的能力和适应性也很高,而其余树种在不同等级下却表现出较大的差异性,甚至没有出现。新疆野苹果、红果山楂和疏花蔷薇等树种的频度与生态位宽度较大,而白柳、锦鸡儿、疣枝桦和欧亚圆柏等树种的频度与生态位宽度较小。随着海拔的增加,大多数树种的频度与生态位宽度出现了先增长后减少的趋势,在海拔的第2和第3等级下,大多数树种可以拥有较好的生存。同时,树种的相对频度的大小与生态位宽度的大小总体呈现出同步的水平。
3.3塔城地区野果林生态位重叠
生态位重叠反映的是生物种群某种特定资源状态的共同利用程度,或者是物种之间生态因子联系上的相似性。通过对塔城地区野果林在4个海拔等级的生态位重叠进行分析可以看出(表4—表7),野果林在4个等级下生态位重叠对数分别是6、78、91、21对,具有生态位重叠(Oik>0)分别占比83.33%、65.38%、60.44%和76.19%,同时,Oik>0.5分别占比33.33%、19.23%、16.48%、28.57%,塔城地区野果林树种之间的生态位重叠随着海拔的增加,表现出先减少后增加的趋势。同时,如新疆野苹果与红果山楂在4个等级下的生态位重叠指数为0.89、0.48、0.57、0.70,也表现出随着海拔的增加,生态位重叠指数也出现了先减少后增加的趋势;但如新疆野苹果与大叶绣线菊为 0.11、0.39、0.83、1.27,新疆野苹果与欧洲稠李为0.00、0.73、0.51、0.07,表现出随着海拔的增加,生态位指数出现了先增加后减少的趋势;而如红果山楂与野扁桃为0.08、0.00、0.00、0.00,红果山楂与大叶绣线菊为 0.00、0.20、0.09、0.14,表现出随海拔增加,生态位重叠指数变化较小或无明显规律。
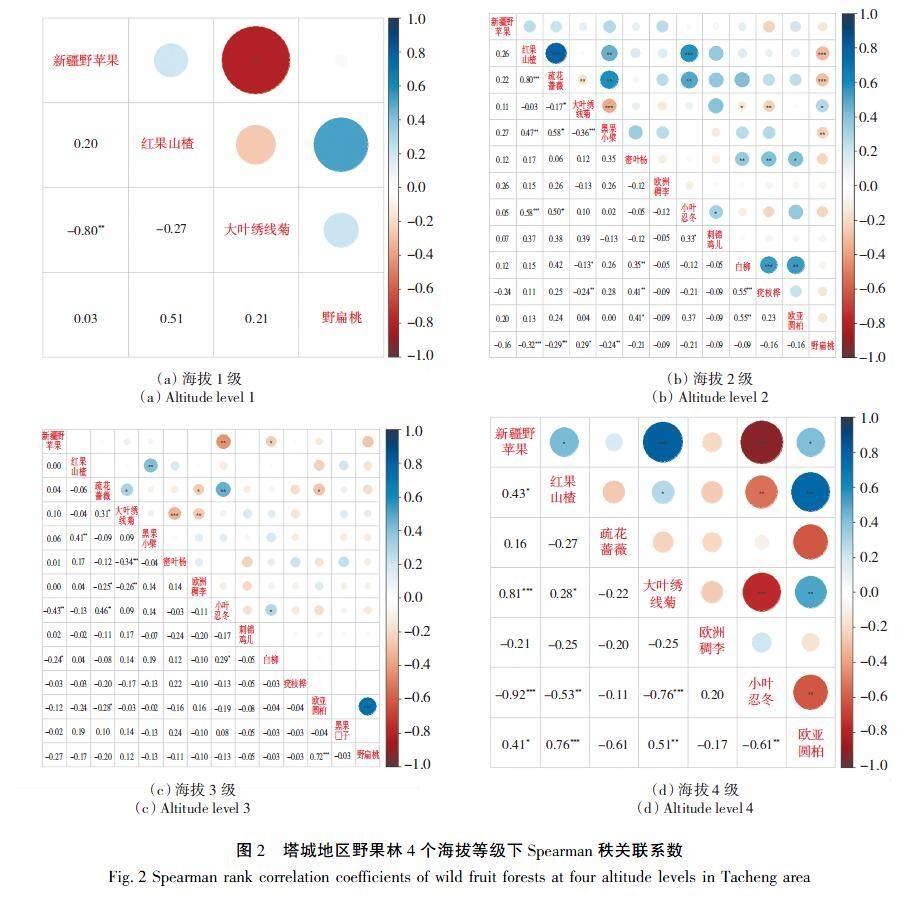
3.4塔城地区野果林种间关联性
在种间关联性中正关联表达了物种间相互吸引和生态位重叠性,负关联表达了物种间相互排斥和生态位分离。对塔城地区野果林4个海拔的种间关联性采用Spearman秩关联系数进行计算,其结果,如图2所示。
由图2可以看出,野果林在4个等级下种间关联性对数分别是6、78、91、21对,呈正关联(rik≥0)分别有4、46、32、9组,分别占比66.67%、58.97%、35.16%、42.86%;显著正关联(rik>0.5)分别有1、5、1、3对,占比16.67%、6.41%、1.10%、14.29%;无明显关联(-0.10≤rik≤0.10)有1、18、45、0对,分别占比16.67%、23.08%、49.45%、0%;显著负关联(rik<-0.50)有1、0、0、5对,分别占比16.66%、0%、0%、23.81%;树种在第1和第2等级下表现为以正关联性为主,而第3和第4等级则以负关联性为主,同时,正关联性、显著正关联性、显著负关联性随着海拔的增加呈现出先减少后增加的趋势,而无明显关联性却表现出先增加后减少的趋势。如新疆野苹果与红果山楂在4个等级下的种间关联性指数为 0.20、0.26、0、0.43,也表现出随着海拔的增加,关联性总体呈现出先减少后增加的趋势;但如新疆野苹果与大叶绣线菊为 -0.80、0.11、0.10、0.81,表现出随着海拔的增加,关联性呈现出增加趋势,而如新疆野苹果与小叶忍冬为0.00、0.05、-0.43、-0.92,表现出随着海拔的增加,关联性呈现出减少的趋势。
4讨论
4.1生境因子
各种生境因子会对群落造成不同的影响,进而引起群落分布和种间关系的变化[32]。本研究表明,在不同等级海拔中树种分布有着显著差异(P=0.000 1),可能由于海拔梯度引起的气候和环境变化,导致植物群落在不同海拔高度表现出差异,如温度、湿度和光照等因素在海拔变化中发挥重要作用,进而塑造了不同海拔下的植物分布[33-34]。新疆野苹果是该区域主要树种,具有较强的抗寒和抗旱等生物学特征。这些生物学特征导致适应海拔为1 100 m~1 300 m(图1),而过高的海拔会因为降水量过高,不利于新疆野苹果等特殊植物的生长[35],同时,海拔过低时会受到气温过高、病虫害和农业等各种因素的干扰[36]。
尽管坡向对物种变化的影响不显著,其Mantel指数为-0.009 5,意味着坡向对野果林树种分布的作用相对较小,然而在实际调查中,坡向的不同对野果林的分布存在一定的影响,野果林主要分布在阴坡、河谷等较为阴暗的环境中,未来需要更详细的研究来解释其在这个特定环境中的作用。土壤pH、土壤含水量、草本盖度对物种变化的正向Mantel指数表明,这些因子与植物群落的变化呈正相关,反映了植物对土壤和水分条件的适应性,以及草本植物在维持生态系统平衡中的重要作用。不同生境因子之间可能存在复杂的交互作用,海拔、坡向和土壤特性等因素在实际环境中往往是相互联系的,共同塑造了植物群落的结构和物种组成[37],塔城地区野果林存在显著的气候和地形差异,导致了生境因子对植物群落的显著影响,气温、湿度、降水等气候因素与地形特征相结合,形成了对生态系统的综合影响[38]。
4.2生态位重叠
由于生态位重叠反映的是生物种群对资源的竞争程度,因此生态位重叠度高,意味着种群之间的竞争激烈或是相似度高[39-40]。研究表明,随着海拔的增加,生态位重叠指数出现了先减少后增加的趋势,代表着树种种间竞争也呈现出先较少后增加的趋势;同时,生态位宽度较大树种往往生态位重叠也较高,生态位宽度较小的树种之间生态位重叠也普遍较小,甚至不存在生态位重叠,生态位重叠与生态位宽度总体呈现出正关联性[41]。树种在资源利用上存在一定的竞争,而生态位宽度的增加可能是为了适应更多的环境条件,降低竞争强度,以保证自身的生存。由于温暖湿润气候条件更加有利于大部分植被生长,资源丰富导致各类树种在此层面上呈现高度重叠,随着海拔的增加,耐寒的树种通过降低自身需求范围来躲避直接竞争[42],因此海拔第2和3等级的生态位重叠指数下降。在海拔第4等级,部分树种的生态位重叠指数再次上升,在较高的海拔条件下,能够生存的树种数量有限,因此生存下来的树种需要利用相似的资源来维持生存,从而导致生态位重叠指数上升。
4.3种间关联性
种间的联结性通常被认为受到环境驱动和资源竞争的影响,种间关联性中正相关表达了物种间相互吸引和生态位重叠性[43],负相关表达了物种间相互排斥和生态位分离[44]。塔城地区野命林种间关联性表现出随着海拔的增加,正关联性、显著正关联性、显著负关联性先减少后增加的趋势,而无明显关联性却表现出先增加后减少的趋势,并且在第1和2等级下,树种间的关联性主要表现为正关联,而在第3和4等级下,树种间的关联性则主要表现为负关联。其原因可能是,不同海拔会影响树种的分布和资源利用,例如,较低海拔地区可能提供更丰富的资源,使得多种树种能够共存,从而产生正关联;而在高海拔地区,由于环境条件更为严酷,资源更为稀缺,树种间可能需要通过分化资源利用来避免竞争,从而产生负关联[45-47];一些生命周期较长、繁殖策略保守、分散能力弱的树种可能更倾向于在低海拔地区形成正关联。同时,种间关联性与生态位重叠有着密切的关系,生态位重叠指数越高,种间关联性越容易表现为负关联,而生态位重叠指数很低或没有时,种间关联性往往表现为不相关,这点与当前诸多研究有着相似的结果[43-44]。
近二十多年来,外来病虫害入侵、农业面积不断扩增和旅游等各种客观原因,导致塔城地区野果林特有树种面临着大面积的死亡。在今后的保护工作中应当重视特有物种的就地保护、切断旅游等外来物种传播途径,选择生态位宽度较大、适生范围较广、生命力较强的新疆野苹果等先锋树种,提高生态系统的稳定性和多样性。进一步深入研究野果林树种共存机制,解决实生苗更新困难问题,建立协调的群落,考虑植物与微生物之间的共生关系,如菌根真菌和植物的互惠共生,以促进根系健康和养分吸收[48-50]。
5结论
塔城地区野果林地理位置较特殊,山区地形复杂,降水量较低,在这样特殊的环境分布的植物具有一定抗旱、抗寒性。由于海拔的不同植物资源分布也有了一定的规律,本研究得出了以下结论。
1)海拔是塔城地区野果林物种分布最主要的影响因子。
2)随着海拔增加,树种频度与生态位宽度出现了先增长后减少的趋势,生态位重叠指数、种间关联性呈现出先减少后增加的趋势。
3)在海拔为1 100~1 300 m时,树种有着更好的分布与较好的种间共存关系。
【参考文献】
[1]ANDEREGG W R L, TRUGMAN A T, BADGLEY G, et al. Climate-driven risks to the climate mitigation potential of forests[J]. Science, 2020, 368(6497): eaaz7005.
[2]GIBSON L, LEE T M, KOH L P, et al. Primary forests are irreplaceable for sustaining tropical biodiversity[J]. Nature, 2011, 478(7369): 378-381.
[3]LUYSSAERT S, SCHULZE E D, BRNER A, et al. Old-growth forests as global carbon sinks[J]. Nature, 2008, 455(7210): 213-215.
[4]BURROWS M T, SCHOEMAN D S, BUCKLEY L B, et al. The pace of shifting climate in marine and terrestrial ecosystems[J]. Science, 2011, 334(6056): 652-655.
[5]LOARIE S R, DUFFY P B, HAMILTON H, et al. The velocity of climate change[J]. Nature, 2009, 462(7276): 1052-1055.
[6]NISHIZAWA K, SHINOHARA N, CADOTTE M W, et al. The latitudinal gradient in plant community assembly processes: a meta-analysis[J]. Ecology Letters, 2022, 25(7): 1711-1724.
[7]LIETH H, WHITTAKE R H. Primary productivity of the biosphere (Vol.14)[M]. New York: Springer Science & Business Media, 1975.
[8]TIEGS S D, COSTELLO D M, ISKEN M W, et al. Global patterns and drivers of ecosystem functioning in rivers and riparian zones[J]. Science Advances, 2019, 5(1): eaav0486.
[9]厉桂香,马克明.土壤微生物多样性海拔格局研究进展[J].生态学报,2018,38(5):1521-1529.
LI G X, MA K M. Progress in the study of elevational patterns of soil microbial diversity[J]. Acta Ecologica Sinica, 2018, 38(5): 1521-1529.
[10]BROOKER R W, MAESTRE F T, CALLAWAY R M, et al. Facilitation in plant communities: the past, the present, and the future[J]. Journal of Ecology, 2008, 96(1): 18-34.
[11]LESSER J S, JAMES W R, STALLINGS C D, et al. Trophic niche size and overlap decreases with increasing ecosystem productivity[J]. Oikos, 2020, 129(9): 1303-1313.
[12]HU Y, WANG H C, JIA H P, et al. Ecological niche and interspecific association of plant communities in alpine desertification grasslands: a case study of Qinghai Lake basin[J]. Plants, 2022, 11(20): 2724.
[13]周赛霞,彭焱松,丁剑敏,等.珍稀植物狭果秤锤树群落木本植物种间联结性及群落稳定性研究[J].广西植物,2017,37(4):442-448.
ZHOU S X, PENG Y S, DING J M, et al. Analysis on community stability and inter-specific correlations among dominant woody populations of the endangered plant Sinojackia rehderiana communities[J]. Guihaia, 2017, 37(4): 442-448.
[14]叶权平,张文辉,于世川,等.桥山林区麻栎群落主要乔木种群的种间联结性[J].生态学报,2018,38(9):3165-3174.
YE Q P, ZHANG W H, YU S C, et al. Interspecific association of the main tree populations of the Quercus acutissima community in the Qiaoshan forest area[J]. Acta Ecologica Sinica, 2018, 38(9): 3165-3174.
[15]DALE M R T, JOHN E A, BLUNDON D J. Contact sampling for the detection of interspecific association: a comparison in two vegetation types[J]. The Journal of Ecology, 1991, 79(3): 781.
[16] ADLER P B, SMULL D, BEARD K H, et al. Competition and coexistence in plant communities: intraspecific competition is stronger than interspecific competition[J]. Ecology Letters, 2018, 21(9): 1319-1329.
[17]李帅锋,刘万德,苏建荣,等.滇西北云南红豆杉群落物种生态位与种间联结[J].植物科学学报,2012,30(6):568-576.
LI S F, LIU W D, SU J R, et al. Niche and interspecific association of species of Taxus yunnanensis communities in northwest Yunnan Province[J]. Plant Science Journal, 2012, 30(6): 568-576.
[18]郑俊鸣,李敏,张盟,等.滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析[J].热带亚热带植物学报,2021,29(5):465-473.
ZHENG J M, LI M, ZHANG M, et al. Interspecific association and niche of mixed forest communities of Casuarina equisetifolia and Dendrocalamus minor var. amoenus in the windbreak of sandy coast[J]. Journal of Tropical and Subtropical Botany, 2021, 29(5): 465-473.
[19]康冰,刘世荣,温远光,等.广西大青山南亚热带次生林演替过程的种群动态[J].植物生态学报,2006,30(6):931-940.
KANG B, LIU S R, WEN Y G, et al. Population dynamics during succession of secondary natural forest in Daqingshan, Guangxi, China[J]. Journal of Plant Ecology, 2006, 30(6): 931-940.
[20]韩大勇,杨永兴,杨允菲,等.松嫩平原破碎化羊草草甸退化演替系列植物多样性的空间格局[J].应用生态学报,2012,23(3):666-672.
HAN D Y, YANG Y X, YANG Y F, et al. Spatial patterns of plant species diversity in a degraded successional series of fragmented Leymus chinensis meadow in Songnen Plain of Northeast China[J]. Chinese Journal of Applied Ecology, 2012, 23(3): 666-672.
[21]邓莉萍,白雪娇,李露露,等.辽东山区次生林优势木本植物种间联结与相关分析[J].生态学杂志,2015,34(6):1473-1479.
DENG L P, BAI X J, LI L L, et al. Interspecific association and correlation among dominant woody plants of secondary forest in montane region of eastern Liaoning Province, China[J]. Chinese Journal of Ecology, 2015, 34(6): 1473-1479.
[22]林培钧,崔乃然.天山野果林资源:伊犁野果林综合研究[M].北京:中国林业出版社,2000.
LIN P J, CUI N R. Wild fruit forests resources in Tianshan Mountains[M]. Beijing: China Forestry Publishing House, 2000.
[23]褚佳瑶,冯琳骄,侯毅兴,等.新疆野苹果种群受损现状[J].经济林研究,2022,40(1):265-273.
CHU J Y, FENG L J, HOU Y X, et al. Analysis on population damage of Malus sieversii[J]. Non-wood Forest Research, 2022, 40(1): 265-273.
[24]李飞飞,崔大方,廖文波,等.中国新疆野苹果(Malus sieversii(Ldb.)Roem.)种群地理分布格局及其遗传关系研究[J].干旱区地理,2011,34(6):926-932.
LI F F, CUI D F, LIAO W B, et al. Geographic distribution pattern and genetic relationship of Malus sieversii (Ldb.) Roem. in China[J]. Arid Land Geography, 2011, 34(6): 926-932.
[25]白玲,阎国荣,许正.伊犁野果林植物多样性及其保护[J].干旱区研究,1998,15(3):10-13.
BAI L, YAN G R, XU Z. Study on Phyto-diversity and conservation of the wild fruit forest of Tianshan Mts in Xinjiang[J]. Arid Zone Research, 1998, 15(3): 10-13.
[26]冯婷婷,周志钦.栽培苹果起源研究进展[J].果树学报,2007,24(2):199-203.
FENG T T, ZHOU Z Q. Advances in research of the origin of cultivated apple[J]. Journal of Fruit Science, 2007, 24(2): 199-203.
[27]羊海军,崔大方,许正,等.中国天山野果林种子植物组成及资源状况分析[J].植物资源与环境学报,2003,12(2):39-45.
YANG H J, CUI D F, XU Z, et al. Analysis on the components and resource situation of seed plants in the wild fruit forest in Tianshan Mountain in China[J]. Journal of Plant Resources and Environment, 2003, 12(2): 39-45.
[28]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
FANG J Y, WANG X P, SHEN Z H, et al. Methods and protocols for plant community inventory[J]. Biodiversity Science, 2009, 17(6): 533-548.
[29]于娜.基于Mantel检验构建基因集差异分析方法[D].济南:山东大学,2021.
YU N. A gene set differential analysis method based on Mantel test[D]. Jinan: Shandong University, 2021.
[30]李秋华,马一明.桐梓河浮游植物优势种生态位与种间联结性分析[J].贵州师范大学学报(自然科学版),2022,40(2):11-18.
LI Q H, MA Y M. Analysis on niche and interspecific association of dominant species of phytoplankton in Tongzi River[J]. Journal of Guizhou Normal University (Natural Sciences), 2022, 40(2): 11-18.
[31]张金龙,马克平.种间联结和生态位重叠的计算:spaa程序包.第十届全国生物多样性保护与持续利用研讨会论文集[C].北京:气象出版社,2014,5(3):165-174.
ZHANG J L, MA K P. Calculation of interspecific linkage and niche overlap: Spaa package. Proceedings of the 10th National Symposium on Biodiversity Conservation and Sustainable Utilization[C]. Beijing: Meteorological Publishing House, 2014, 5(3): 165-174.
[32]金山,武帅楷.太行山南段油松林火烧迹地优势草本生态位及种间关系[J].北京林业大学学报,2021,43(4):35-46.
JIN S, WU S K. Niche and interspecific association of dominant species in herb layer of burned Pinus tabuliformis forest in the southern Taihang Mountain of Northern China[J]. Journal of Beijing Forestry University, 2021, 43(4): 35-46.
[33]唐志红,尉秋实,刘虎俊,等.祁连山东段高寒植被群落特征及其与地形气候因子关系研究[J].生态学报,2020,40(1):223-232.
TANG Z H, YU Q S, LIU H J, et al. Characteristics of alpine vegetation community and its relationship to topographic climate factors in the eastern Qilian Mountain[J]. Acta Ecologica Sinica, 2020, 40(1): 223-232.
[34]唐志尧,方精云,张玲.秦岭太白山木本植物物种多样性的梯度格局及环境解释[J].生物多样性,2004,12(1):115-122.
TANG Z Y, FANG J Y, ZHANG L. Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains[J]. Chinese Biodiversity, 2004, 12(1): 115-122.
[35]田中平,庄丽,李建贵,等.伊犁河谷北坡野果林木本植物种间关系及环境解释[J].生物多样性,2011,19(3):335-342,396-397.
TIAN Z P, ZHUANG L, LI J G, et al. Interspecific and environmental relationships of woody plant species in wild fruit-tree forests on the north slope of Ili Valley[J]. Biodiversity Science, 2011, 19(3): 335-342, 396-397.
[36]刘爱华.新疆野果林苹果小吉丁与优势天敌生物学、生态学研究[D].乌鲁木齐:新疆农业大学,2010.
LIU A H. Biology and ecology of Agrilus mali Matsumura and its dominant natural enemies in wild fruit forests in Xinjiang[D]. Urumqi: Xinjiang Agricultural University, 2010.
[37]BIRHANU L, BEKELE T, TESFAW B, et al. Relationships between topographic factors, soil and plant communities in a dry Afromontane forest patches of Northwestern Ethiopia[J]. PLoS One, 2021, 16(3): e0247966.
[38]高福元,赵成章,卓马兰草.高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性[J].生态学报,2014,34(16):4698-4706.
GAO F Y, ZHAO C Z, ZHUO M L C. Spatial distribution and spatial association of Stellera chamaejasme population in the different altitude in in degraded alpine grassland[J]. Acta Ecologica Sinica, 2014, 34(3): 605-612.
[39]舒琪,徐瑞晶,胡璇,等.海南岛甘什岭青梅群落主要乔木树种生态位与种间联结[J].生态学杂志,2021,40(9):2689-2697.
SHU Q, XU R J, HU X, et al. Niche and interspecific association of the main tree species in Vatica mangachapoi communities in Ganshiling, Hainan Island, China[J]. Chinese Journal of Ecology, 2021, 40(9): 2689-2697.
[40]霍红,冯起,苏永红,等.额济纳绿洲植物群落种间关系和生态位研究[J].中国沙漠,2013,33(4):1027-1033.
HUO H, FENG Q, SU Y H, et al. Interspecies relationship and niche analysis on phytocoenosium in the Ejina Oasis[J]. Journal of Desert Research, 2013, 33(4): 1027-1033.
[41]沈雪梨,吴远密,朱艺璇,等.濒危植物百花山葡萄群落物种生态位特征与种间联结研究[J].植物科学学报,2020,38(2):195-204.
SHEN X L, WU Y M, ZHU Y X, et al. Community niche characteristics and interspecific associations of critically endangered species, Vitis baihuashanensis M. S. Kang et D. Z. Lu[J]. Plant Science Journal, 2020, 38(2): 195-204.
[42]MARTORELL C, FRECKLETON R P. Testing the roles of competition, facilitation and stochasticity on community structure in a species-rich assemblage[J]. Journal of Ecology, 2014, 102(1): 74-85.
[43]刘润红,陈乐,涂洪润,等.桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J].生态学报,2020,40(6):2057-2071.
LIU R H, CHEN L, TU H R, et al. Niche and interspecific association of main species in shrub layer of Cyclobalanopsis glauca community in Karst hills of Guilin, southwest China[J]. Acta Ecologica Sinica, 2020, 40(6): 2057-2071.
[44]刘润红,常斌,荣春艳,等.漓江河岸带枫杨群落主要木本植物种群生态位[J].应用生态学报,2018,29(12):3917-3926.
LIU R H, CHANG B, RONG C Y, et al. Niche of main woody plant populations of Pterocarya stenoptera community in riparian zone of Lijiang River, China[J]. Chinese Journal of Applied Ecology, 2018, 29(12): 3917-3926.
[45]李丘霖,宗秀虹,邓洪平,等.赤水桫椤群落乔木层优势物种生态位与种间联结性研究[J].西北植物学报,2017,37(7):1422-1428.
LI Q L, ZONG X H, DENG H P, et al. Niche and interspecific association of dominant species in tree layer of Chishui Alsophilaspinulosa Community[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(7): 1422-1428.
[46]郑俊鸣,李敏,张盟,等.滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析[J].热带亚热带植物学报,2021,29(5):465-473.
ZHENG J M, LI M, ZHANG M, et al. Interspecific association and niche of mixed forest communities of Casuarina equisetifolia and Dendrocalamus minor var. amoenus in the windbreak of sandy coast[J]. Journal of Tropical and Subtropical Botany, 2021, 29(5): 465-473.
[47]GU L, GONG Z W, LI W Z. Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on Loess Plateau, P.R. China[J]. Scientific Reports, 2017, 7(1): 6604.
[48]WU S N, WEN L, DONG S K, et al. The plant interspecific association in the revegetated alpine grasslands determines the productivity stability of plant community across restoration time on Qinghai-Tibetan Plateau[J]. Frontiers in Plant Science, 2022, 13: 850854.
[49]刘秉儒,张秀珍,胡天华,等.贺兰山不同海拔典型植被带土壤微生物多样性[J].生态学报,2013,33(22):7211-7220.
LIU B R, ZHANG X Z, HU T H, et al. Soil microbial diversity under typical vegetation zones along an elevation gradient in Helan Mountains[J]. Acta Ecologica Sinica, 2013, 33(22): 7211-7220.
[50]巴音达拉.病原菌及菌根菌维持森林植物多样性机制的研究进展[J].中国野生植物资源,2023,42(10):96-100.
BAYANDALA. Advances in mechanisms of pathogenic and mycorrhizal fungi maintenance on forest plant diversity[J]. Chinese Wild Plant Resources, 2023, 42(10): 96-100.
