不同氮肥与糖蜜配施对烟田土壤酶活性、温度和微生物群落的影响
2024-07-03李佳李云捷陈振国喻雪婧云月利朱蓉李亚东
李佳 李云捷 陈振国 喻雪婧 云月利 朱蓉 李亚东

doi:10.15889/j.issn.1002-1302.2024.10.036
摘要:为探究不同种类氮肥与糖蜜配施对烟草根际土壤环境的影响,以湖北省利川市海拔1 200 m处的植烟土壤为研究对象,以烟草专用肥为对照,探究了不同施肥组合对土壤微生物群落结构的变化以及对土壤酶活性和土壤温度的影响。结果表明:(1)土壤微生物测序结果表明,多肽能够提高硝化螺旋菌门的相对丰度,糖蜜的添加显著提高了微生物种群的数量和多样性;(2)氨基酸母液粉能显著提高脲酶活性(P<0.05),其中氨基酸母液粉与 150 kg/hm2 糖蜜(M2)处理的脲酶活性最高,与对照组相比增加了68.5%;糖蜜的施加能显著提高蔗糖酶活性(P<0.05),多肽与450 kg/hm2糖蜜(T4)处理的蔗糖酶活性最高,与对照组相比增加了63.1%;(3)糖蜜的施加能快速提高苗期土壤温度,烟苗移栽7 d后多肽与450 kg/hm2糖蜜(T4)处理土壤温度最高,与烟草专用肥处理组相比提高了 2.07 ℃;(4)相关性分析结果表明,土壤温度与微生物数量之间存在极显著的正相关关系(P<0.01),土壤中脲酶活性与土壤温度显著相关(P<0.05),蔗糖酶活性与土壤温度极显著相关(P<0.01)。研究表明,多肽与450 kg/hm2糖蜜配施能够通过改良烟田土壤微生物群落结构,提高硝化螺旋菌门的物种丰度和化能异养型细菌功能的占比,增强土壤脲酶与蔗糖酶的活性,在烟草种植苗期提高烟田土壤温度,在解决高海拔地区低温导致的烟草苗期生长缓慢方面具有实际生产意义。
关键词:土壤;微生物;温度;酶活性;氮源;糖蜜
中图分类号:S572.606 文献标志码:A
文章编号:1002-1302(2024)10-0260-08
收稿日期:2023-11-07
基金项目:湖北省重点研发计划(编号:2023BCB077)。
作者简介:李 佳(1999—),女,河南周口人,硕士研究生,研究方向为烟草栽培与土壤改良。E-mail:1154772796@qq.com。
通信作者:李亚东,博士,教授,博士研究生导师,研究方向为微生物工程、环境生态学、蛋白废弃物资源化利用。E-mail:lyd55555@sina.com。
烟草是我国重要的经济作物,是卷烟工业的基础,烟叶的产量和质量直接影响着烟草的种植收益[1]。近年来,由于烟田长期连作、化肥的不科学使用导致烟田土壤板结酸化,严重影响作物的生长发育与品质,具体表现为土壤有机质含量下降、pH值降低、土壤微生态失去平衡等[2]。其原因主要是高山地区烟草苗期温度过低,烟农通常使用覆膜的方法保证烟苗生长的温度,地膜的残留部分在土壤中逐年堆积、难以分解,导致土壤板结、通透性变差、土壤微生物活性降低,影响烟株对养分的吸收,造成烟叶产量品质下降,烟田土壤肥力降低[3]。因此,如何增强高海拔地区土壤温度、降低连作障碍、提高烟叶质量成了烟草生产中一个亟待解决的问题。
对于不同海拔的植烟土壤而言,由于土壤的理化性质、养分供应状况、酶活性与微生物群落动态不尽相同,对烟株生长发育进程造成的影响也不同[4]。烟草是一种喜温作物,最适温度为25~28 ℃,而烟苗种植时期在5月上旬,气温通常达不到烟苗生长所需的标准[5]。土壤温度对烟草前期生长发育和烟叶品质具有重要影响,高海拔烟区土壤温度过低会导致同化物质的转化和积累受到抑制,肥料利用速率下降,烟草生长缓慢,妨碍烟叶正常成熟,而生产出劣质烟叶[6-8]。在农业生产中,通常使用覆膜、深耕等方式来提高土壤温度。目前关于烤烟地膜覆盖栽培与深耕栽培等方面已有大量研究,初步探明了提高温度能够促进植株对肥料的利用效率、加快烟株生长,提高烟叶的产量和品质[9-11]。土壤温度作为影响土壤微生物群落结构的重要因素,与土壤的理化性质以及养分利用效率密切相关[12-13]。现有的研究主要集中在通过改变栽培方式来增强土壤的保温保墒能力,但通过改良施肥组合来提高土壤温度并系统探究温度与酶活性和土壤微生物群落结构三者之间相互关系的研究还相对较少。
Morley等研究发现,土壤中的有机质能为大多数异养微生物提供碳源,并为土壤呼吸作用提供基质,添加碳源能够增强土壤呼吸强度并显著增加土壤微生物量,通过微生物生长代谢活动产生的生物热能有效提高烟田土壤温度[14-16]。为此,本研究以高海拔地区植烟土壤为切入点,以烟草种植的专用肥(硝态氮)、多肽(蛋白氮)、氨基酸母液粉(氨基酸氮)作为土壤微生物的氮源,配施微生物生长的碳源——糖蜜(糖厂下脚料,含糖量50%),通过研究不同氮源与不同用量的糖蜜结合配施的方式,探讨微生物群落结构与土壤酶活性和土壤温度三者的相互作用关系,旨在探究一种新型肥料对烟田土壤环境的改良作用,为保证高海拔地区烟苗发育温度和促进烟苗前期生长提供理论依据和科学指导。
1 材料与方法
1.1 土壤背景概况
本试验于2023年5月对位于湖北省利川市柏杨坝镇海拔1 200 m处的大田植烟土壤进行分析,植烟品种为云烟87,供试土壤类型为黄棕壤,土壤基本理化性质见表1。
1.2 肥料的理化性质
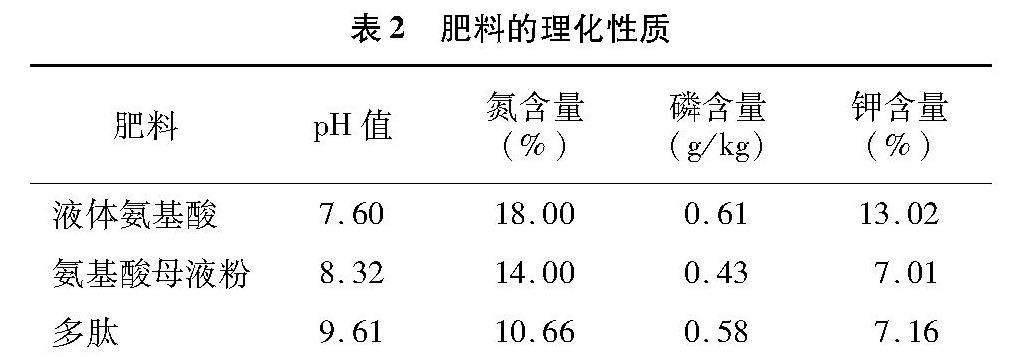
本研究所用的氨基酸母液粉与多肽肥是笔者所在实验室自主研发的新型有机肥料,是将动物羽毛等蛋白废弃物,在高温条件下水解转化成全水溶性的多肽和氨基酸等小分子物质;所用的液体氨基酸肥是将蛋白废弃物用强酸水解为小分子物质,使用氢氧化钾中和;烟草专用肥购自史丹利农业集团股份有限公司;糖蜜购自山东长惠化工有限公司,为制糖工业下脚料,主要成分为蔗糖。肥料的理化性质详见表2。
1.3 试验设计
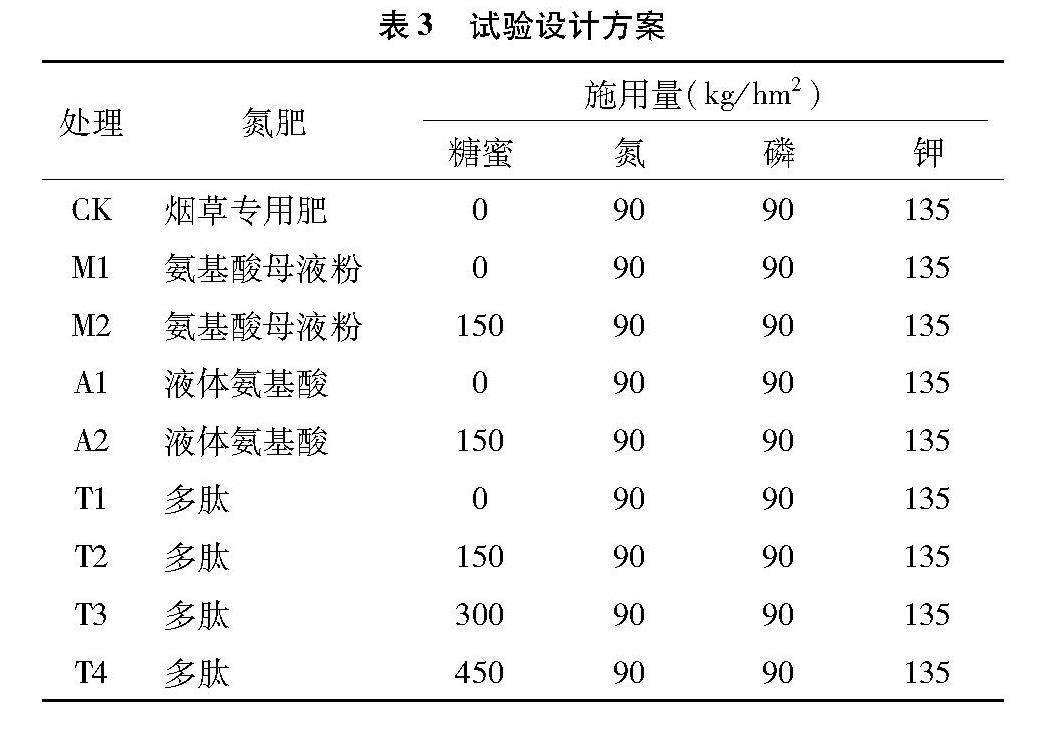
本试验共设4种氮肥处理:烟草专用肥(硝态氮)、氨基酸母液粉(氨基酸氮)、液体氨基酸肥(液体氨基酸氮)、多肽(蛋白氮),以烟草专用肥作对照,按照施氮肥90 kg/hm2,氮磷钾比例为1 ∶1 ∶1.5配施磷肥与钾肥,磷肥为过磷酸钙,钾肥为硫酸钾。同时对多肽组合设4种不同用量的糖蜜作底肥处理,具体试验设计组合见表3。每个处理设置30个生物学重复。
1.4 土壤酶活性测定
过氧化氢酶活性的测定采用高锰酸钾滴定法,蔗糖酶活性的测定采用3,5-二硝基水杨酸比色法,脲酶活性的测定采用苯酚钠-次氯酸钠比色法[17]。
1.5 土壤温度测定
使用 HC-D40 型号土壤熵情检测器,测量移栽后 28 d 内的烟苗根部土壤温度。为减少太阳光照对土壤温度的影响,选择每日20:00进行测量,严格控制每日测量时间,时间误差不超过10 min[18]。
1.6 土壤样品采集
采样时间为移栽后28 d,采用五点取样法从烟株根部取混合样作为1个处理的代表样品,每个处理设置3次重复,将采集的土样保存于-80 ℃冰箱中用于微生物测序。
1.7 土壤微生物测序
本研究基于Illumina NovaSeq测序平台对文库进行双末端测序,使用CTAB法从样品中提取总DNA,利用2%琼脂糖凝胶电泳检测样品DNA的浓度和纯度,使用带Barcode的特异引物,New England Biolabs公司的Phusion High-Fidelity PCR Master Mix with GC Buffer和高效高保真酶进行PCR。使用TruSeq DNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建,构建好的文库经过Qubit和Q-PCR定量。文库合格后,使用NovaSeq 6000进行上机测序。对测序得到的有效数据进行物种注释以及丰度分析,揭示样本的物种构成,探究不同施肥处理间微生物种群结构差异。
1.8 数据分析
使用Qiime软件(version 1.9.1)计算微生物多样性指数;使用R软件(version 2.15.3)绘制稀释曲线、丰度等级曲线和物种累积曲线;使用R软件进行α多样性指数组间差异分析;使用Origin 2022、SPSS 22.0等软件进行数据处理和多因素方差分析;使用Spearman分析土壤微生物多样性指数与土壤温度和土壤酶活性之间的相关性。
2 结果与分析
2.1 不同施肥处理后土壤酶活性差异比较
由图1可以看出,不同施肥处理后,土壤酶活性有显著差异。由图1-a可以看出,不同处理之间,氨基酸母液粉与150 kg/hm2糖蜜处理的脲酶活性最高,与对照组相比显著增加486 μg/(g·d)。对比不同种类氮肥对脲酶活性的影响,与烟草专用肥相比,施加氨基酸母液粉的组合脲酶活性显著增加了253 μg/(g·d),施加多肽组合脲酶活性显著增加了176 μg/(g·d),施加液体氨基酸对脲酶活性没有显著影响。比较不同用量糖蜜对脲酶活性的影响,施加150、300、450 kg/hm2糖蜜土壤脲酶活性均有显著增加,其中施加150 kg/hm2糖蜜脲酶活性最高,增加糖蜜用量后,脲酶活性略有下降,但降幅不显著。
由图1-b可以看出,不同处理之间,多肽与450 kg/hm2糖蜜处理的蔗糖酶活性最高,与对照组相比显著增加9.97 mg/(g·d)。对比不同种类氮肥对蔗糖酶活性的影响,与烟草专用肥相比,施加氨基酸母液粉的组合蔗糖酶活性显著增加了 3.46 mg/(g·d),施加多肽的组合蔗糖酶活性增加了2.33 mg/(g·d),施加液体氨基酸对蔗糖酶活性没有显著影响。比较不同用量糖蜜对蔗糖酶活性的影响可以看出,随糖蜜施加量增加,土壤蔗糖酶活性呈增加趋势,其中施加450 kg/hm2糖蜜蔗糖酶活性最高。
由图1-c可以看出,不同处理之间,多肽与300 kg/hm2糖蜜处理的过氧化氢酶活性最高,其次为氨基酸母液粉不施加糖蜜处理。对比不同种类氮肥对过氧化氢酶活性的影响,氨基酸母液粉处理显著高于其他氮肥处理。比较不同用量糖蜜对过氧化氢酶活性酶活性的影响发现,随施加糖蜜量增加,过氧化氢酶活性呈增加趋势,施加300 kg/hm2糖蜜时过氧化氢酶活性显著最高,继续增加糖蜜用量后,酶活性降低。
2.2 不同施肥处理后土壤温度差异比较
分析不同施肥处理后土壤温度的变化规律,结果如图2所示,移栽后28 d内不同种类氮肥处理及不同用量糖蜜处理后的土壤温度均存在一定差异,但不同日期的温度差异有所不同。由图 2-a 可以看出,不同种类氮肥处理对土壤温度具有一定影响,移栽后7 d各处理间温度差异达到最大。在烟苗移栽1周后使用氨基酸母液粉施肥处理的土壤温度最高,显著高于烟草专用肥处理1.47 ℃;在烟苗移栽2周后,使用多肽施肥处理的土壤温度最高,显著高于烟草专用肥处理0.93 ℃;随着移栽时间的增加,各处理间土壤温度差异逐渐降低,当烟苗移栽28 d后,不同氮肥处理后的土壤温度无显著差异。由图2-b可以看出,不同用量的糖蜜对土壤温度的
影响更为明显,在烟苗移栽前期,土壤温度与施加糖蜜的总量呈显著正相关,烟苗移栽1周后,施加450 kg/hm2糖蜜处理土壤温度比不施加糖蜜高1.14 ℃,比烟草专用肥处理提高2.07 ℃;烟苗移栽2周后,施加450 kg/hm2糖蜜处理土壤温度显著高于不施加糖蜜处理 0.84 ℃;烟苗移栽3周后,各处理间土壤温度差异逐渐降低,当烟苗移栽28 d后,不同用量糖蜜处理后的土壤温度无显著差异。这表明在烟苗移栽前期,氨基酸母液粉与多肽能有效提高土壤温度,施加糖蜜对短期内提高烟田土壤温度具有明显效果。
2.3 烟田土壤微生物多样性
2.3.1 土壤微生物OTU分析
为研究不同施肥处理后烟田土壤的微生物物种组成,对所有处理样本的有效序列以97%的一致性进行稀释性曲线分析,结果如图3-a所示,随着样品数量的增加,各个样本曲线均趋于平稳,这说明根际微生物群的细菌种类达到饱和阶段;稀释性曲线可以表明本研究测序数据量的合理性,可以看出,T4 处理样本物种数量最多,这说明多肽与450 kg/hm2糖蜜配施提高了土壤微生物种群的数量。丰度等级曲线可用来解释物种丰度和物种均匀度,物种的丰度由曲线的宽度来反映,物种的丰度越高,曲线在横轴上的范围越大;曲线的形状(平缓程度)反映了样品中物种的均度,曲线越平缓,则表明物种分布越均匀。如图3-b所示,T4处理物种丰度最高且微生物种群分布更均匀。
2.3.2 土壤微生物物种组成
不同处理中优势细菌相对丰度如图4所示。从门水平(图4-a)上看,不同处理间共同的优势菌门主要有变形菌门(Proteobacteria)、拟杆菌门(Bacteroidota)、硝化螺旋菌门(Nitrospirae)、厚壁菌门(Firmicutes),占全部序列的69.63%~82.87%,对比M1和M2、A1和A2、T1和T2处理可以看出,添加糖蜜有效增加了土壤微生物的物种丰富度。其中,变形菌门作为细菌中最大的门,广泛存在于自然界中的土壤、水体和动物体内等环境中,其具有一系列重要的生物功能,例如在氮循环、有机物分解、土壤修复等生态过程中起到重要作用[19],变形菌门作为相对丰度最高(47.25%~65.82%)的优势菌门,在纯液体氨基酸处理中相对丰度高于其他组合。从属水平(图4-b)上看, 对相对丰度前 20 名的细菌组成进行分析,
可以看出,多肽处理的组合中硝化螺旋菌属(Nitrospira)的相对丰度明显高于对照组。硝化螺旋菌属于亚硝酸盐氧化细菌,在陆地环境中普遍存在,并且在农业土壤的生物氮循环和硝化作用中发挥重要作用,硝化螺旋菌属在多肽处理中丰度较高,表明多肽的施加使烟田土壤中氮循环发生了改变。
2.3.3 土壤微生物α多样性分析
对不同样本在97%一致性阈值下的α多样性分析指数进行统计,由表4可知,各处理土壤样品覆盖度均达到0.99,说明测序结果的真实情况良好;Chao1指数是菌种丰富度指数,添加糖蜜处理的Chao1指数高于CK,其中T4处理细菌 Chao1 指数高于T1、T2和T3处理,这说明增加糖蜜用量能够提升细菌丰富度;M2处理Shannon指数高于M1处理,A2处理 Shannon指数高于A1处理,T4处理Shannon指数高于T1、T2和T3处理, 这说明施加糖蜜有助于提高群落多样性以及物种分布均匀程度。α多样性分析指数表明,施加糖蜜有助于提高烟草苗期烟田土壤微生物丰富度与多样性。
2.3.4 烟田土壤细菌群落功能
本研究使用FAPROTAX数据库,能较好地预测环境样本中原核生物的生物化学循环过程[20],揭示烟田土壤在不同施肥处理后细菌群落的生态功能,进一步探究不同氮肥与糖蜜的结合对烟田土壤细菌群落生化过程的改变。基于FAPROTAX数据库功能注释,本研究选取丰度排名前20的功能绘制聚类热图,如图5所示,与对照组相比,施加糖蜜有助于提高化能异养型和好氧化能异养型细菌功能的占比,这些功能可以消化分解土壤中的有机物释放能量[21]。其中T4处理明显提高了与氮循环相关功能的丰度,如亚硝酸盐反硝化、氮呼吸、硝酸盐反硝化、硝酸盐呼吸、硝酸盐还原等功能,这些微生物的生命代谢活动给土壤提供大量热量。
2.4 土壤温度与微生物多样性指数和土壤酶活性的相关性分析
对土壤温度与土壤微生物多样性指数和土壤酶活性进行相关性分析,由表5中结果可知,土壤温度与细菌 Chao1 指数存在显著正相关关系(P<0.05),与ACE指数存在极显著的正相关关系(P<0.01),相关系数达0.829;土壤温度与脲酶活性存在显著正相关关系,相关系数达0.785(P<0.05),与蔗糖酶活性存在极显著正相关关系(P<0.01),相关系数达0.938。脲酶活性与细菌ACE指数存在显著的正相关关系(P<0.05);蔗糖酶活性与细菌Chao1指数和ACE指数存在极显著的正相关关系(P<0.01),相关系数分别达0.826、0.828;而过氧化氢酶活性与土壤微生物多样性指数相关性不显著。这说明多肽和氨基酸母液粉与糖蜜配施,能够有效通过提高土壤微生物种群多样性与酶活性来实现土壤温度的提高。
3 讨论
本试验以1 200 m高海拔地区植烟土壤为研究对象,利用 Illumina 高通量测序检验不同施肥处理后烟田土壤的微生物群落结构,结合不同处理土壤酶活性与土壤温度的差异做相关性分析,探究糖蜜与不同种类氮肥结合配施对提高高海拔地区烟田土壤温度的作用。结果表明,使用多肽代替烟草专用肥,并适量添加糖蜜配施能够提高土壤微生物种群的数量和多样性,并提高土壤酶活性与温度。土壤微生物是衡量土壤有效养分的重要指标,是植物吸收养分的重要驱动者,在养分的转化和供应方面起重要作用[22]。相关研究表明,糖蜜的添加能够增强微生物对碳源的利用能力和代谢能力,本试验使用糖蜜作为土壤的外加碳源,能有效改良烟田土壤的微生物群落结构,其原理是不同的微生物对不同的碳源利用效率不同,这与杨泳等的研究结果[23]一致。糖蜜和多肽的施入为土壤微生物的生命活动提供充足的碳源和氮源,土壤细菌的数量明显增多[24]。对比不同处理后微生物群落结构差异,多肽与糖蜜配施明显提高了氮循环功能微生物的相对丰度,根际土壤细菌的优势属为硝化螺旋菌,其属于亚硝酸盐氧化细菌,在农业土壤的生物氮循环和硝化作用中发挥重要作用[25]。
土壤酶与微生物一起参与土壤的生物化学过程和物质循环,与土壤的代谢过程密切相关,是土壤养分转化的重要标志[26-27]。本研究结果表明,微生物种群数量与脲酶活性呈现显著正相关关系,与蔗糖酶活性呈现极显著正相关关系,微生物多样性与蔗糖酶活性呈现显著正相关关系;而过氧化氢酶与土壤微生物多样性指数无明显相关。土壤脲酶在土壤有机物氮键的水解作用中起重要作用,其活性增强能够有效促进土壤中的有机氮转化为有效的无机氮被植株吸收利用,通常用来表征土壤氮素的供应状况,因此脲酶活性的增强对于提高烟田土壤肥力、生态系统生产力以及促进土壤生态系统中的物质循环极为关键[28];而土壤蔗糖酶活性与土壤中营养元素含量、微生物数量及土壤呼吸强度密切相关,可以作为评价土壤肥力的重要指标[29]。本研究结果表明,土壤脲酶活性与多肽以及糖蜜的添加具有显著相关性。究其原因是土壤脲酶活性随土壤养分含量的增加而变大,土壤全氮和有机质含量通过直接和间接效应成为土壤脲酶活性的主要影响因素[30-31]。这与董齐琪等的研究结果[32]相一致。
本研究通过改良施肥组合来达到提高烟田土壤温度促进植物生长的目的,分析土壤微生物多样性指数与土壤温度的关系发现,土壤温度与微生物种群和数量显著相关。研究结果表明多肽与糖蜜的配施能改变土壤微生物种群结构,大幅提高微生物数量以及土壤中脲酶和蔗糖酶活性,这说明施肥组合的改良能够影响土壤性质[33]。此外,土壤微生物在生长活动过程中产生的代谢物质能够促进土壤有机质的分解,这使得土壤的理化性质发生改变,土壤酶活性增加,又对微生物的呼吸代谢活动产生影响[34-36],这些影响可能是提高烟田土壤温度、改善土壤品质的重要原因。
4 结论
(1)比较不同施肥组合下土壤脲酶活性差异,不同种类氮肥对脲酶活性的影响显著,氨基酸母液粉与150 kg/hm2糖蜜处理的脲酶活性最高,与对照组相比增加了68.5%;比较不同施肥组合下土壤蔗糖酶活性差异,糖蜜用量对蔗糖酶活性影响显著,随糖蜜施加量增加,土壤蔗糖酶活性呈增加趋势,其中多肽与450 kg/hm2糖蜜处理蔗糖酶活性最高,与对照组相比增加了 63.1%;比较不同施肥组合下土壤过氧化氢酶活性差异,多肽与300 kg/hm2糖蜜处理的过氧化氢酶活性最高,与对照组相比增加了30.0%。
(2)分析不同施肥处理后土壤温度的变化规律,结果表明,施加不同用量的糖蜜对土壤温度的影响较大,在烟苗移栽前期,土壤温度与施加糖蜜的用量呈正相关,在烟苗移栽7 d后,各处理间土壤温度差异达到最大,多肽与450 kg/hm2 糖蜜处理土壤温度最高,与烟草专用肥处理组相比提高了2.07 ℃。
(3)不同施肥组合下烟田土壤微生物群落结构存在差异,糖蜜的添加显著提高了微生物种群的数量和多样性,其中多肽与450 kg/hm2糖蜜配施处理的微生物数量与多样性指数最高。使用 FAPROTAX 数据库对土壤微生物测序结果进行预测,结果表明,与烟草专用肥对比,施加氨基酸母液粉与多肽可以提高与氮循环相关的功能丰度,施加糖蜜有助于提高化能异养型和好氧化能异养型细菌功能的相对丰度。
(4)对不同土壤温度与土壤微生物多样性指数和酶活性进行相关性分析,结果表明,土壤温度与微生物数量存在极显著的正相关关系,微生物种群数量增加对土壤温度具有显著影响;微生物种群数量对脲酶活性也具有显著影响;微生物数量和多样性均对蔗糖酶活性具有显著影响;而过氧化氢酶与土壤微生物多样性指数无明显相关性。
参考文献:
[1]Yang L Y,Yang S L,Li J Y,et al. Effects of different growth temperatures on growth,development,and plastid pigments metabolism of tobacco (Nicotiana tabacum L.) plants[J]. Botanical Studies,2018,59(1):5.
[2]查宇璇,冉 茂,周鑫斌. 烟田土壤酸化原因及调控技术研究进展[J]. 土壤,2022,54(2):211-218.
[3]高维常,蔡 凯,曾陨涛,等. 农用地膜残留对土壤氮素运移及烤烟根系生长的影响[J]. 土壤学报,2020,57(6):1556-1563.
[4]杨 承,莫 江,蹇国友,等. 不同海拔烟区覆膜天数对土壤温度及烟株生长的影响[J]. 安徽农业科学,2013,41(3):1001-1005.
[5]沈少君,郭学清,郑玉木,等. 低温胁迫对烤烟生长和产质量的影响[J]. 中国烟草科学,2010,31(6):35-37.
[6]樊希彬,李 丹,左晓晴,等. 气候条件对烟草生长的影响分析[J]. 黑龙江农业科学,2016(4):27-30.
[7]田 峰,陈前锋,单海涛,等. 移栽期对烤烟农艺性状及经济性状与上部烟叶可用性的影响[J]. 作物研究,2010,24(3):184-186,191.
[8]邱雪柏,高维常,陈 伟,等. 井窖式移栽烟田温湿度对烟株前期生长发育的影响[J]. 烟草科技,2015,48(7):20-24,35.
[9]冯 军,石 超,Cholidah L,等. 不同覆盖类型下减量施肥对油菜产量及水肥利用效率影响[J]. 农业工程学报,2019,35(15):85-93.
[10]刘智炫,周清明,穰中文,等. 深耕对植烟土壤温湿度及烤烟根系发育和经济性状的影响[J]. 烟草科技,2019,52(12):23-30.
[11]王 毅,刘志刚,刘永新,等. 氧化-生物双降解膜对土壤水热变化及烟株生长的影响[J]. 中国烟草科学,2023,44(1):24-31.
[12]Heinze J,Gensch S,Weber E,et al. Soil temperature modifies effects of soil biota on plant growth[J]. Journal of Plant Ecology,2017,10(5):808-821.
[13]Ma L,Guo C,Lü X,et al. Soil moisture and land use are major determinants of soil microbial community composition and biomass at a regional scale in northeastern China[J]. Biogeosciences,2015,12(8):2585-2596.
[14]Morley N,Baggs E M. Carbon and oxygen controls on N2O and N2 production during nitrate reduction[J]. Soil Biology and Biochemistry,2010,42(10):1864-1871.
[15]强学彩,袁红莉,高旺盛. 秸秆还田量对土壤CO2释放和土壤微生物量的影响[J]. 应用生态学报,2004,15(3):469-472.
[16]崔 鹏,艾超凡,廖汉鹏,等. 超高温堆肥微生物群落强化产热功能特征分析[J]. 土壤学报,2022,59(6):1660-1669.
[17]关松荫. 土壤酶及其研究法[M]. 北京:农业出版社,1986:115-173.
[18]张 衡. 播期和土壤温度对芝麻种子出苗的影响[J]. 种子科技,2023,41(3):1-4.
[19]王 丹. 海洋细菌SAR86全球分布及其多样性研究[D]. 舟山:浙江海洋大学,2020:1-2.
[20]Louca S,Parfrey L W,Doebeli M. Decoupling function and taxonomy in the global ocean microbiome[J]. Science,2016,353(6305):1272-1277.
[21]伍建业,吴永贵,兰美燕,等. 复合人工湿地对陆基水产养殖废水中氮磷的净化及其微生物群落特征[J]. 环境工程学报,2023,17(2):517-531.
[22]王传杰,肖 婧,蔡岸冬,等. 不同气候与施肥条件下农田土壤微生物生物量特征与容量分析[J]. 中国农业科学,2017,50(6):1067-1075.
[23]杨 泳,王新池,尹统统,等. 低剂量糖蜜添加对吉富罗非鱼养殖水质和微生物代谢多样性的影响[J]. 淡水渔业,2022,52(6):36-43.
[24]Gryta A,Frc M,Oszust K. Genetic and metabolic diversity of soil microbiome in response to exogenous organic matter amendments[J]. Agronomy,2020,10(4):546.
[25]侯海军,秦红灵,陈春兰,等. 土壤氮循环微生物过程的分子生态学研究进展[J]. 农业现代化研究,2014,35(5):588-594.
[26]徐祥玉,孟贵星,袁家富,等. 翻压绿肥对植烟土壤活性有机质和土壤酶的影响[J]. 中国烟草科学,2011,32(增刊1):103-107.
[27]施 娴,刘艳红,王田涛,等. 有机肥与烟草专用肥配施对植烟土壤微生物和土壤酶活性的动态变化[J]. 土壤通报,2017,48(5):1126-1131.
[28]王凤娟,秦绍龙,张鸿雁,等. 不同森林类型土壤酶活性和微生物数量特征研究[J]. 陕西农业科学,2021,67(10):75-80,111.
[29]万 晓,田丹青,潘刚敏,等. 不同肥料对有机栽培薄荷生长和土壤酶及微生物量的影响[J]. 浙江农业科学,2023,64(11):2642-2645.
[30]陈双林,郭子武,杨清平. 毛竹林土壤酶活性变化的海拔效应[J]. 生态学杂志,2010,29(3):529-533.
[31]刘广深,徐冬梅,许中坚,等. 用通径分析法研究土壤水解酶活性与土壤性质的关系[J]. 土壤学报,2003,40(5):756-762.
[32]董齐琪,王海燕,杜 雪,等. 东北低山区典型林分类型土壤脲酶活性特征[J]. 应用与环境生物学报,2023,29(3):690-695.
[33]许文欢,张雅坤,王国兵,等. 不同施肥方式对苏北杨树人工林土壤微生物碳源代谢的影响[J]. 生态学杂志,2015,34(7):1791-1797.
[34]章家恩,刘文高,胡 刚. 不同土地利用方式下土壤微生物数量与土壤肥力的关系[J]. 土壤与环境,2002,11(2):140-143.
[35]黄昆鹏,董昆乐,李芳芳,等. 烟草抗病嫁接对根际土壤微生物多样性的影响[J]. 江苏农业科学,2023,51(20):239-247.
[36]闫秋艳,董 飞,段增强,等. 土壤温度对不同施肥方式下辣椒生长及土壤理化性质的影响[J]. 华北农学报,2018,33(2):195-201.
