武夷山地方茶树种质生化特性和茶多糖清除超氧阴离子自由基活性分析
2024-07-01石玉涛谢惠珍郑淑琳羽观华王飞权李力张渤李远华罗盛财
石玉涛 谢惠珍 郑淑琳 羽观华 王飞权 李力 张渤 李远华 罗盛财

摘要:为探明武夷山地方茶树种质主要生化成分特性和茶多糖清除超氧阴离子自由基活性差异,测定了31份武夷山地方茶树种质主要生化成分含量,采用水提醇沉淀法提取茶多糖,并采用黄嘌呤氧化酶法比较了茶多糖SOD活力。结果表明,武夷山地方茶树种质主要生化成分存在较强的变异和遗传多样性,具有较大的遗传改良潜力。平均变异系数为21.84%,平均遗传多样性指数为2.08,平均改良潜力为50.92%;茶多酚、游离氨基酸、黄酮类化合物、可溶性糖含量和酚氨比均呈正态分布;茶多酚含量与黄酮类化合物含量呈极显著正相关,与可溶性糖含量呈极显著负相关,游离氨基酸含量与可溶性糖含量呈极显著负相关;提取的前2个主成分包含了5个生化指标81.08%的信息,游离氨基酸和茶多酚含量是5个生化性状的特征指标,主成分综合得分排名前5位的茶树种质为‘白牡丹‘红海棠‘向天梅‘半天妖和‘香石角,可作为武夷山茶区优质乌龙茶品种选育和推广栽培的良好材料;基于生化成分的聚类分析将31份茶树种质分为3类,第Ⅰ类群13份种质茶多酚和黄酮类化合物含量高,可溶性糖含量低,第Ⅱ类群8份种质游离氨基酸含量较低,可溶性糖含量较高,第Ⅲ类群10份种质酚氨比较高,可溶性糖含量中等;基于茶多糖SOD活力的聚类分析,可将31份茶树种质分为3类,第Ⅰ类群8份种质茶多糖SOD活力低,第Ⅱ类群19份种质茶多糖SOD活力居中,第Ⅲ类群4份种质茶多糖SOD活力高;茶多糖SOD活力存在丰富的变异,变异系数达40.62%,遗传多样性指数为2.07;‘白鸡冠‘大红袍‘大红梅和‘灵芽4份种质茶多糖SOD活力高,清除超氧阴离子自由基能力强。研究结果可为武夷山地方茶树种质的改良和开发利用提供科学依据。
关键词:武夷山;茶树种质;生化成分;茶多糖;超氧阴离子自由基;主成分分析;聚类分析
doi:10.13304/j.nykjdb.2023.0622
中图分类号:S571.1 文献标志码:A 文章编号:10080864(2024)05006512
种业是农业的“芯片”,种质资源又是种业的“芯片”。种质资源研究和创新的深度和广度,直接影响种质资源的利用效率和种业的可持续发展。近年来,农业种质资源的保护和开发利用已成为世界各国农业科技创新驱动战略的重要组成部分[1]。茶树[Camellia sinensis (L.) O. Kuntze]种质资源是茶树品种选育和创新的基础,在实现茶产业优质、高产、高效中发挥着重要作用[2]。茶产业是福建省武夷山市的特色产业、支柱产业和富民产业。武夷山地处我国茶树区划的江南茶区,是茶树迁移、演化的过渡带。武夷山独特的地理和气候条件孕育出丰富的地方特色茶树种质资源,素有“茶树品种王国”之称[3]。地方种质蕴含着丰富的变异类型和稀有性状,开展地方茶树种质的发掘利用对促进武夷山茶产业的转型升级和创新发展具有重要意义。茶叶中的生化成分不仅是茶树育种的物质基础,同时也决定着茶树品种的适制性和茶叶感官品质的形成[4]。目前,已有研究者针对武夷山地方茶树种质的农艺性状[5]、叶片解剖结构[6]、矿质元素[7]等方面进行了分析,筛选出一些优异资源,但对其生化特性的分析尚不全面。茶叶是一种有益健康的饮料,也是提取茶多酚、茶氨酸和茶多糖等生物活性物质的来源。茶多糖是一类与蛋白质结合的杂多糖,具有提高免疫力、抗氧化、降血糖等多种生物活性[8]。茶树是茶叶中天然活性成分的“生物工厂”,可以合成具有工业和制药价值的生物分子[9]。筛选茶多糖含量高或生物活性强的茶树种质,对于今后选育功能型茶树品种具有重要意义。不同品种茶树的茶多糖在组成、理化性质和抗氧化活性上存在差异[1011]。研究发现,武夷山不同地方茶树种质的茶多糖提取率及其清除1,1-二苯基-2-苦基肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基和羟基自由基的活性存在丰富的多样性[12],但不同地方茶树种质茶多糖清除超氧阴离子自由基(superoxideanion free radical,O-2·)的活性差异尚不明确。因此,本研究选取31份武夷山地方茶树种质,对其主要生化成分含量进行测定,并提取不同茶树种质的茶多糖,测定其对超氧阴离子自由基的清除活性,通过多元统计分析方法探明武夷山地方茶树种质生化特性和对超氧阴离子自由基(O-2·)清除活性的差异,筛选清除超氧阴离子自由基能力强的茶树种质材料,以期为武夷山地方茶树种质的改良和开发利用提供科学依据。
1 材料与方法
1.1 供试材料
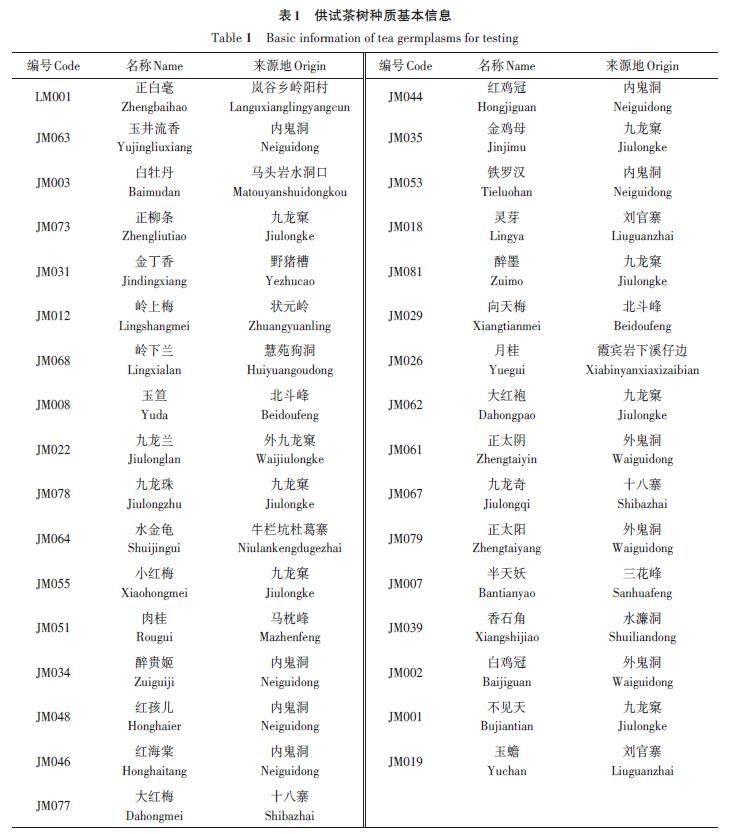
31份参试武夷山地方茶树种质(表1)样品采集自武夷山龟岩种植园茶树资源圃,位于福建省武夷山风景区梅子桥(27°36′28″ N,117°57′57″ E),海拔202 m,圃内立地条件和肥水管理一致。每份种质采集春季第1轮新梢1芽3叶,蒸汽杀青固样,烘干、粉碎后过40目筛,密封后置于-20 ℃冰箱中保存备用,每份种质重复3次。
1.2 仪器与试剂
FZ102型植物粉碎机购自北京市永光明医疗仪器有限公司;Alpha2-4型冷冻干燥机购自德国Christ公司;JJ-1型精密增力电动搅拌器购自江苏中大仪器科技有限公司;UV-3200PC型分光光度计购自上海美普达仪器有限公司;硫酸亚铁、酒石酸钾钠、谷氨酸、水合茚三酮、氯化亚锡、三氯化铝、蒽酮均为分析纯,购自昀冠(上海)生物科技有限公司;超氧化物歧化酶(superoxide dismutase,SOD)测试盒购自南京建成生物工程研究所。
1.3 试验方法
1.3.1 生化成分含量测定
采用酒石酸铁比色法测定茶多酚(tea polyphenols,TP)含量,采用茚三酮比色法测定游离氨基酸(amino acids,AA)总量,采用三氯化铝比色法测定黄酮类化合物(flavones,FLA)总量[13];采用硫酸-蒽酮比色法[14]测定可溶性糖(soluble sugar,SS)含量。
1.3.2 茶多糖提取
采用水提醇沉淀法[12]提取茶多糖。取粉碎后的茶样,加入55 ℃纯水(固液比为1 g∶20 mL),置于55 ℃水浴锅中磁力搅拌辅助浸提2.5 h,过滤除去茶渣后,滤液以4 000 r·min-1离心15 min,离心所得上清液中加入3倍体积的95%乙醇,沉淀茶多糖,然后将混合液在4 000 r·min-1离心15 min,取沉淀,冷冻干燥后即得茶多糖。
1.3.3 茶多糖清除超氧阴离子自由基活性测定
将提取的茶多糖配制成1 mg·mL-1 的溶液,采用黄嘌呤氧化酶法测定超氧化物歧化酶(SOD)活性,按照SOD测试盒(南京建成)说明书进行测定。以类SOD活性(nU·mL-1)高低表示茶多糖样品对超氧阴离子自由基(O-2·)清除率的相对大小[11]。
1.4 数据处理
采用Excel 2021软件进行数据整理和统计分析;利用SPSS 26.0软件进行正态分布检验、方差分析和主成分分析,组间差异采用单因素方差分析(One-Way ANOVA),组间两两比较采用Duncan检验法,正态分布检验采用Shapiro-Wilk法[15],主成分分析提取特征值>1的因子作为主成分,并利用主成分对不同茶树种质进行综合评价,综合评价函数计算方法为主成分得分与相应权重求和[16];采用TBtools v1.123软件进行层次聚类分析(hierarchical cluster analysis, HCA)并绘制聚类热图,聚类方法为Complete-linkage 法,距离为欧氏距离[17];采用Origin Pro 2023软件绘图;遗传多样性指数(H′)采用Shannon-Wiener 指数法[18]计算;各性状的改良潜力[19]计算公式如下。
改良潜力=(Xmax-Xmean)/Xmean×100% (1)
式中,Xmax为测定指标最高含量,Xmean为测定指标平均含量。
2 结果与分析
2.1 武夷山地方茶树种质主要生化成分差异和分布状态
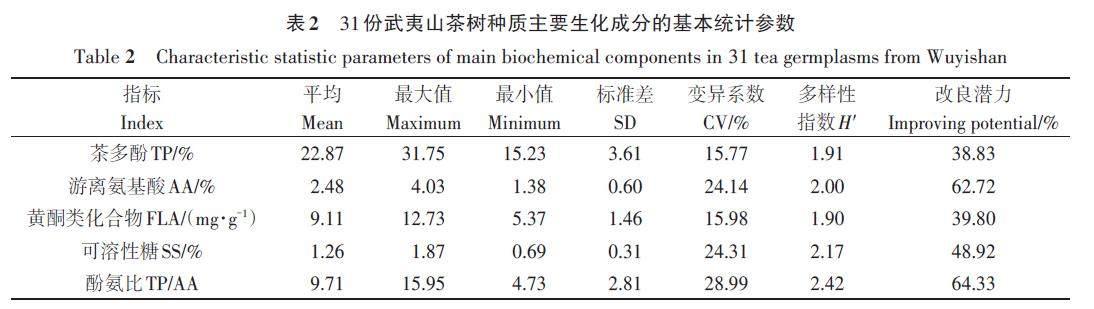
对31份茶树种质的主要生化成分含量进行测定,结果(表2)表明,5个生化指标含量差异明显。茶多酚含量为15.23%~31.75%,含量最高的种质是JM019(‘ 玉蟾),最低的种质是JM026(‘月桂);游离氨基酸含量为1.38%~4.03%,含量最高的种质是JM068(‘岭下兰),最低的种质是JM029(‘向天梅);黄酮类化合物含量为5.37~12.73 mg·g-1,含量最高的种质是JM046(‘ 红海棠),最低的种质是JM079(‘正太阳);可溶性糖含量为0.69%~1.87%,含量最高的种质是LM001(‘正白毫),最低的种质是JM002(‘白鸡冠);酚氨比为4.73~15.95,比值最高的种质是JM003(‘白牡丹),最低的种质是JM026(‘月桂)。5个指标的变异系数为15.77%~28.99%,平均变异系数为21.84%,可见茶树种质在5个生化指标上存在丰富的变异类型,其中变异系数最大的指标是酚氨比,最小的是茶多酚含量。5个指标的平均遗传多样性指数为2.08,表明31份武夷山地方茶树种质在生化成分含量上具有较为丰富的遗传多样性;且各生化成分指标均具有不同程度的改良潜力,其中酚氨比的改良潜力最大,为64.33%,茶多酚含量改良潜力较低,为38.83%。
对5个生化指标的分布进行检验,结果(图1)显示,茶多酚、游离氨基酸、黄酮类化合物和可溶性糖的含量及酚氨比值均呈现正态分布,表明在31份武夷山地方茶树种质中大部分种质的4种化学成分含量和酚氨比处于平均水平。
2.2 不同茶树种质生化成分间的相关性分析
5个生化指标的相关性分析结果如图2所示。茶多酚含量与黄酮类化合物含量呈极显著正相关,与酚氨比呈显著正相关,与可溶性糖含量呈极显著负相关;游离氨基酸含量与可溶性糖含量和酚氨比呈极显著负相关。
2.3 武夷山地方茶树种质生化成分的主成分分析和综合评价
对31份茶树种质5个生化指标进行主成分分析,提取到2个特征值>1的主成分,累计方差贡献率达81.80%,包含了5个生化指标的大部分信息(图3)。第1主成分代表了主要生化成分42.40%的信息,主要影响因子是游离氨基酸含量(影响为负),其次为可溶性糖含量;第2主成分代表了主要生化成分39.40%的信息,主要影响因子是茶多酚含量,其次为黄酮类化合物含量。
31 份茶树种质的主成分综合得分如图4 所示。第1主成分得分较低的种质有JM068(‘岭下兰)、JM044(‘ 红鸡冠)、JM002(‘ 白鸡冠)、JM062(‘大红袍)、JM051(‘肉桂),表明这些种质在游离氨基酸含量方面的品质较其他种质表现更优;得分较高的种质有JM029(‘ 向天梅)、JM079(‘正太阳)、LM001(‘正白毫)、JM035(‘金鸡母)、JM046(‘红海棠),表明这些种质在游离氨基酸含量方面的品质较其他种质表现较差,应在育种中加以改良。第2主成分得分较高的种质有JM003(‘白牡丹)、JM046(‘红海棠)、JM019(‘玉蟾)、JM002(‘白鸡冠)、JM051(‘肉桂),表明这些种质在茶多酚和黄酮类化合物含量方面的品质较其他种质表现较优。综合得分排名前5位的种质为JM003(‘白牡丹)、JM046(‘红海棠)、JM029(‘向天梅)、JM007(‘半天妖)、JM039(‘香石角),这些茶树种质可作为武夷山茶区优质乌龙茶品种选育和推广栽培的良好材料。
2.4 基于生化成分含量的武夷山地方茶树种质的聚类分析
基于5个生化指标对31份茶树种质进行聚类分析,结果(图5)表明,按照生化成分含量高低,可将31 份种质划分为3 个类群。第Ⅰ类包含JM067、JM044、JM002、JM051、JM019、JM064、JM008、JM022、JM046、JM003、JM055、JM012、JM007 共13 份种质;第Ⅱ类包含JM029、JM035、JM061、JM031、JM039、JM048、LM001、JM079 共8份材料;第Ⅲ 类包含JM018、JM077、JM034、JM081、JM078、JM063、JM062、JM053、JM068、JM026共10份种质。
对3个类群5个生化指标进行方差分析,结果(图6)显示,第Ⅰ类群的茶多酚和黄酮类化合物含量显著高于其余2个类群,但可溶性糖含量较低;第Ⅱ类群的游离氨基酸含量较低,但可溶性糖含量较高;第Ⅲ类群的酚氨比较高,可溶性糖含量处于中等水平。
2.5 武夷山地方茶树种质茶多糖SOD 活力分析
对茶多糖SOD活力进行测定,结果显示,31份茶树种质茶多糖的SOD活力为1.93~18.83 nU·mL-1,其中,SOD活力最高的种质是JM002(‘白鸡冠),最低的种质是JM029(‘向天梅)。SOD活力变异系数达40.62%,遗传多样性指数为2.07,改良潜力为93.49%,显示武夷山地方茶树种质茶多糖在清除O-2·能力方面存在丰富的变异和遗传多样性,具有较大的改良潜力。
以31份武夷山地方茶树种质茶多糖SOD活力绘制热图,并进行层次聚类分析(图7),可将31份种质分为3个类群。第Ⅰ类群包含JM029、JM022、JM061、JM063、JM031、JM068、JM003、JM078共8份种质,此类群种质中茶多糖SOD活力低,清除O-2·的能力较低;JM048、JM044 和JM007等19份种质为第Ⅱ类,此类群种质中茶多糖SOD 活力中等,清除O-2·能力居中;第Ⅲ类包含JM002、JM062、JM077和JM018共4份种质,此类群种质中的茶多糖SOD活力最高,清除O-2·能力强。
3 讨论
茶树地方种质是长期自然选择演化而来的,它们保持了较高的遗传多样性。地方种质资源的遗传多样性是丰富种质资源遗传多样性的基础,也是选育新品种的重要途径和手段[20]。然而多年来,随着茶产业向集约化方向发展,武夷山茶区对地方种质的关注度明显降低,且缺乏系统深入的研究,导致其利用效率较低[21]。地方种质在产量等相关性状上与育成品种有一定的差距,但在品质和功能育种上有很强的优势[22]。近年来,随着茶树育种的目标向优质化转变[23],系统评价和利用地方种质是拓展武夷山茶区茶树资源遗传基础的重要途径。本研究通过对武夷山31份地方茶树种质的5个生化指标进行分析,它们的遗传多样性指数为1.90~2.42,表明5个生化指标的遗传基础较广,可从中筛选高生化成分含量的优异种质材料,为茶树品质和功能育种提供基础,进一步提高武夷山地方茶树种质的利用效率和价值。相关性分析表明,茶多酚含量与黄酮类化合物含量呈极显著正相关,与可溶性糖含量呈极显著负相关;游离氨基酸含量与可溶性糖含量呈极显著负相关,表明武夷山地方茶树种质新梢内的茶多酚与黄酮类化合物之间存在着某种程度的协同累积效应,而茶多酚、游离氨基酸与可溶性糖则表现出不同的累积特征。因此,在以武夷山的地方茶树种质为材料开展育种工作时,可根据各主要生化成分之间的相关性选择亲本材料,或以简单易测的性状为指标,实现同时改良多个性状,提高育种效率[7]。主成分分析提取出2个主成分,代表了5个生化指标81.80%的信息,其中游离氨基酸和茶多酚含量可作为评价武夷山地方茶树种质的特征指标。主成分综合得分排名前5位的种质为‘白牡丹‘红海棠‘向天梅‘半天妖‘香石角,它们的生化成分综合品质高,可作为茶树优质育种的亲本材料,具有较高的开发利用潜力。
酚氨比是衡量茶树种质新梢适制性的重要指标[24],酚氨比较低(<8)的茶树种质适制绿茶,酚氨比较高(>15)的茶树种质适制红茶,而酚氨比适中(8~15)的茶树种质较适制乌龙茶[25]。31份武夷山地方茶树种质的酚氨比为4.73~15.95,变异系数为28.99%,即酚氨比变异幅度较大,其中酚氨比小于8的种质有9份,适合加工绿茶;酚氨比高于15的种质有2份,适制红茶;酚氨比为8~15的种质有20份,占供试种质的64.52%,适制乌龙茶。武夷山是闽北乌龙茶武夷岩茶的主产区,这些地方茶树种质资源为特色和多样化武夷岩茶产品的生产提供丰富的种质基础。
茶树种质资源中,有部分种质将其加工制作茶叶后品质不佳,但仍可以作为茶叶活性成分提取的原料,提高其经济价值[26]。茶多糖是一类天然的抗氧化和降血糖活性成分,具有高茶多糖含量或生物活性的茶树品种可以作为茶多糖的提取来源,成为“茶多糖工厂”[27]。超氧阴离子是一种活性氧,它是细胞中氧分子受单一电子还原的产物,在细胞的氧化还原代谢中发挥重要作用[28]。超氧阴离子在细胞内能够通过SOD转化为H2O2,当生物体内超氧阴离子含量超过细胞所能调节的范围时,可能会引起细胞坏死、DNA损伤、脂质过氧化等[29]。SOD活力(清除O-2·活性)已被广泛应用于植物多糖抗氧化活性的评价中[30-32]。本研究结果表明,31份茶树种质茶多糖SOD活力变异系数达40.62%,遗传多样性指数为2.07,显示武夷山地方茶树种质茶多糖在清除O-2·能力方面存在丰富的变异。课题组前期研究发现,武夷山地方茶树种质中的茶多糖在清除DPPH自由基和羟基自由基活性方面具有丰富的遗传多样性[12],说明武夷山地方茶树种质为筛选含高清除自由基活性茶多糖种质材料奠定了良好的基础,其茶多糖在体外清除自由基活性方面具有较大的改良潜力。本研究从31份武夷山地方茶树种质中筛选出‘白鸡冠‘大红袍‘大红梅‘灵芽共4份茶多糖清除O-2·能力强的种质,可作为培育高茶多糖活性茶树品种的优选种质,在茶树功能育种和茶叶深加工领域加以利用。
参考文献
[1] 黎裕,李英慧,杨庆文,等.基于基因组学的作物种质资源研究:现状与展望[J].中国农业科学,2015,48(17):3333-3353.
LI Y, LI Y H, YANG Q W, et al .. Genomics-based cropgermplasm research: advances and perspectives [J]. Sci. Agric.Sin., 2015, 48(17):3333-3353.
[2] 金基强,张晨禹,马建强,等.茶树种质资源研究“十三五”进展及“十四五”发展方向[J].中国茶叶,2021,43(9):42-49,76.
JIN J Q, ZHANG C Y, MA J Q, et al .. Research progress on teagermplasms during the “13th five-year” plan period anddevelopment direction in the“ 14th five-year” plan period [J].China Tea J., 2021, 43(9):42-49,76.
[3] 李力,罗盛财,王飞权,等. 基于GBS-SNP 的武夷茶树(Camellia sinensis, Synonym: Thea bohea L.)遗传分析及标记开发[J].茶叶科学,2023,43(3):310-324.
LI L, LUO S C, WANG F Q, et al .. Genetic analysis and markerdevelopment for Wuyi tea (Camellia sinensis, synonym: Theabohea L.) based on GBS-SNP [J]. J. Tea Sci., 2023, 43(3):310-324.
[4] 董方,李小飞,沈思言,等.江西茶树资源的遗传多样性分析及优异种质筛选[J]. 江西农业大学学报,2022,44(6):1466-1477.
DONG F, LI X F, SHEN S Y, et al .. Genetic diversity analysisand screening of excellent germplasm of tea plant resources inJiangxi [J]. Acta Agric. Univ. Jiangxiensis, 2022, 44(6):1466-1477.
[5] 王飞权,冯花,罗盛财,等.武夷名丛茶树种质资源农艺性状多样性分析[J].中国农业科技导报,2019,21(6):43-54.
WANG F Q, FENG H, LUO S C, et al .. Diversity analysis ofagronomic traits of Wuyi Mingcong tea plant germplasmresources [J]. J. Agric. Sci. Technol., 2019, 21(6):43-54.
[6] 王飞权,李纪艳,冯花,等.武夷名丛茶树种质资源叶片解剖结构分析[J].热带作物学报,2019,40(12):2375-2389.
WANG F Q, LI J Y, FENG H, et al .. Analysis of leafanatomical structure of Wuyi Mingcong tea germplasmresources [J]. Chin. J. Trop. Crops, 2019, 40(12):2375-2389.
[7] 石玉涛,郑淑琳,王飞权,等.武夷名丛茶树种质资源矿质元素含量特征分析[J].中国农业科技导报,2020,22(7):37-50.
SHI Y T, ZHENG S L, WANG F Q, et al .. Characteristicsanalysis of mineral element contents in Wuyi Mingcong teaplant germplasm resources [J]. J. Agric. Sci. Technol., 2020,22(7):37-50.
[8] DU L L, FU Q Y, XIANG L P, et al .. Tea polysaccharides andtheir bioactivities [J]. Molecules, 2016, 25(2):144-149.
[9] YAO J, WENG Y, DICKEY A, et al .. Plants as factories forhuman pharmaceuticals: applications and challenges [J]. Int. J.Mol. Sci., 2015, 16(12):28549-28565.
[10] 陈薛,左欣欣,徐安安,等.不同茶树品种鲜叶多糖的理化性质和抗氧化活性比较研究[J].茶叶科学,2022,42(6):806-818.
CHEN X, ZUO X X, XU A A, et al .. Comparative study on thephysicochemical characteristics and antioxidant activities ofpolysaccharides in different tea cultivars [J]. J. Tea Sci., 2022,42(6):806-818.
[11] 陈玉琼,余志,张芸,等.茶树品种、部位和嫩度对茶多糖含量和活性的影响[J]. 华中农业大学学报, 2005, 24(4):406-409.
CHEN Y Q, YU Z, ZHANG Y, et al .. Effect of tea cultivars andtenderness on tea polysaccharide [J]. J. Huazhong Agric. Univ.,2005, 24(4):406-409.
[12] 石玉涛,郑淑琳,吴伟伟,等.武夷名丛茶树种质资源茶多糖抗氧化活性分析[J].福建农业学报,2020,35(7):801-810.
SHI Y T, ZHENG S L, WU W W, et al .. Antioxidant activity ofpolysaccharides in Wuyi Mingcong tea germplasms [J]. FujianJ. Agric. Sci., 2020, 35(7):801-810.
[13] 李远华.茶学综合实验[M]. 北京:中国轻工业出版社, 2018:193-194.
[14] 张正竹.茶叶生物化学实验教程(第二版)[M].北京:中国农业出版社, 2021:44-45.
[15] 魏广伟,阳慧怡,王敏,等.芝麻种质资源表型性状遗传多样性分析及综合评价[J].江苏农业科学,2022,50(18):122-130.
[16] 代涛,万嘉欣,黎洁华,等.基于主成分与聚类分析综合评价杧果种质资源果实糖酸品质[J].果树学报,2022,39(12):2253-2263.
DAI T, WAN J X, LI J H, et al .. Comprehensive evaluation offruit sugar and acid quality of mango germplasm based onprincipal component and cluster analysis [J]. J. Fruit Sci.,2022, 39(12):2253-2263.
[17] CHEN C J, CHEN H, ZHANG Y, et al .. TBtools: an integrativetoolkit developed for interactive analyses of big biological data [J].Mol. Plant, 2020, 13(8): 1194-1202.
[18] 杨春,郭灿,乔大河,等.三都野生茶树表型性状和生化组分多样性分析[J].江苏农业科学,2023,51(8):111-119.
[19] 杨丽英,陈进,葛再伟.云南花生种质丰产性和品质性状的改良潜力[J].花生学报,2002,31(1):33-36.
YANG L Y, CHEN J, GE Z W, et al .. Improving potential ofproductivity and quality in groundnut in Yunnan province [J].J. Peanut Sci., 2002, 31(1):33-36.
[20] 严碧蓉,黄飞毅,蓝华中.地方特色茶树种质资源江华苦茶研究与产业化开发[J].湖南农业,2022(8):30-31.
[21] 罗盛财,陈德华,黄贤格,等.武夷名丛单丛茶树种质资源收集、整理鉴定与保护利用研究[J]. 中国茶叶,2017,39(12):18-20.
[22] 闫满朝,肖长顺,陈志龙,等.西乡县地方茶树种质资源的生物学性状观察及生化成分分析[J]. 西北农业学报,2020,29(8):1224-1231.
YAN M C, XIAO C S, CHEN Z L, et al .. Analysis of biologicalcharacteristics and biochemical components of tea germplasmresources in Xixiang county [J]. Acta Agric. Bor-Occid. Sin.,2020, 29(8):1224-1231.
[23] XIA E H, TONG W, WU Q, et al .. Tea plant genomics:achievements, challenges and perspectives [J/OL]. Hortic.Res., 2020, 7:7 [2023-07-08]. https://doi.org/10.1038/s41438-019-0225-4.
[24] 疏再发,刘瑜,邵静娜,等.浙南早生茶树种质资源主要品质成分分析及优异资源鉴选[J]. 浙江农业学报,2022,34(11):2438-2450.
SHU Z F, LIU Y, SHAO J N, et al .. Analysis of main qualitycomponents and selection of excellent resources of earlysproutingtea germplasm resources in southern Zhejiang, China [J].Acta Agric. Zhejiangensis, 2022, 34(11):2438-2450.
[25] 江新凤,李琛,曹挥华,等‘. 黄金菊和‘宁州2号茶树生化成分季节变化特征分析[J]. 食品安全质量检测学报,2023,14(10):261-269.
JIANG X F, LI C, CAO H H, et al .. Seasonal variationcharacteristics of biochemical components of Camellia sinensisvar.‘ Huangjinju and Camellia sinensis var.‘ Ningzhou 2 [J].J. Food Saf. Qual., 2023, 14(10):261-269.
[26] SARKER U, OBA S. Polyphenol and flavonoid profiles andradical scavenging activity in leafy vegetable Amaranthusgangeticus [J/OL]. BMC Plant Biol., 2020, 20(1):2700-0 [2023-07-08]. https://doi.org/10.1186/s12870-020-02700-0.
[27] FAN Y, ZHOU X, HUANG G. Preparation, structure, andproperties of tea polysaccharide [J]. Chem. Biol. Drug Des.,2022, 99(1):75-82.
[28] JIE Z, LIU J, SHU M, et al .. Detection strategies for superoxideanion: a review [J/OL]. Talanta, 2022, 236: 122892 [2023-07-08]. https://doi.org/10.1016/j.talanta.2021.122892.
[29] VO Q V, HOA N T, THONG N M, et al .. The hydroperoxyl andsuperoxide anion radical scavenging activity of anthocyanidinsin physiological environments: theoretical insights into mechanismsand kinetics [J/OL]. Phytochemistry, 2021, 192: 112968 [2023-09-08]. https://doi.org/10.1016/j.phytochem.2021.112968.
[30] LIU Y, SUN Y, HANG G. Preparation and antioxidantactivities of important traditional plant polysaccharides [J]. Int.J. Biol. Macromol., 2018, 111:780-786.
[31] CHEN L, HUANG G. Antioxidant activities of phosphorylatedpumpkin polysaccharide [J]. Int. J. Biol. Macromol., 2019, 125:256-261.
[32] CHEN F, HUANG G, YANG Z, et al .. Antioxidant activity ofMomordica charantia polysaccharide and its derivatives [J].Int. J. Biol. Macromol., 2019, 138:673-680.
(责任编辑:张冬玲)
基金项目:福建省自然科学基金项目(2023J011048);福建省大学生创新训练计划项目(S202210397038, S202210397057);中央引导地方科技发展专项(2021L3058);武夷学院科技创新发展基金项目(2019J04)。
