木棉树形结构和叶性状对生境要素的响应研究
2024-06-27毛开泽高漫娟吴睿张月萍程希平
毛开泽 高漫娟 吴睿 张月萍 程希平


DOI: 10.11931/guihaia.gxzw202211012
毛开泽, 高漫娟, 吴睿, 等, 2024.
木棉树形结构和叶性状对生境要素的响应研究 [J].
广西植物, 44(5): 863-872.
MAO KZ, GAO MJ, WU R, et al., 2024.
Tree shape structure and leaf trait responses to habitat elements of Bombax ceiba [J].
Guihaia, 44(5): 863-872.
摘 要: 植物形态结构是植物本身与外部环境共同作用的结果。为了分析木棉树形结构与叶性状的相关关系,探索地形、土壤和气象要素对木棉生长的影响,以及揭示木棉的生长策略和适应机制,该文以云南省西双版纳国家级自然保护区内230棵木棉为研究对象,测定6个树形结构指标,8个叶性状指标以及所在区域地形、气象、土壤养分等生境要素,使用结构方程模型和变差分解分析各生境要素对木棉树形结构和叶性状的影响和解释度。结果表明:(1)木棉树形结构各观测指标之间、叶性状各观测指标之间存在相关性(P<0.05)。(2)生境要素各观测指标对木棉生长具有较强影响,其中年均降雨量、坡度、平均温度对木棉生长指标的解释率较高,是影响木棉生长的主要生境指标。(3)从标准化路径系数看,3个生境要素对木棉树形结构和叶性状影响具有一致性,均表现为地形要素>气象要素>土壤要素。(4)3个生境要素叠加解释了树形结构和叶性状分别为43.5%和12.3%,表现出树形结构对生境环境的响应大于叶性状对环境响应的特点。研究结果初步探明了木棉不同生长指标对环境要素的适应策略,为木棉在异质生境的培育、繁殖以及木棉人工林高效经营等提供了理论依据和实践参考。
关键词: 木棉, 生境要素, 树形结构, 叶性状, 冗余分析, 结构方程模型
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2024)05-0863-10
收稿日期: 2023-04-05 接受日期: 2023-06-29
基金项目: 国家自然科学基金(31860206)。
第一作者: 毛开泽(1997—),硕士研究生,研究方向为植物地理,(E-mail)1965567607@qq.com。
*通信作者: 程希平,博士,教授,研究方向为森林生态学,(E-mail)xipingcheng2012@163.com。
Tree shape structure and leaf trait responses to
habitat elements of Bombax ceiba
MAO Kaize1, GAO Manjuan1, WU Rui1, ZHANG Yueping1, CHENG Xiping1,2*
( 1. School of Geography and Ecotourism, Southwest Forestry University, Kunming 650224, China; 2. Southwest Research
Center for Eco-Civilization, National Forestry and Grassland Administration, Kunming 650224, China )
Abstract: Morphology and structure of plants are the results from the joint action of itself and the external environment. In order to analyze the correlation between tree shape structure and leaf trait of Bombax ceiba, and to explore the influence of topographic, soil, and meteorological elements on its growth shape, and to reveal its growth strategy and adaptation mechanism, we studied 230 B. ceiba trees in Xishuangbanna National Nature Reserve of Yunnan Province and measured six tree shape structure indexes, eight leaf trait indexes, and habitat elements of regional topographic, meteorology, and soil. The effects and explantation of habitat elements on tree shape structure and leaf trait of B. ceiba were analyzed by structural equation model and variation decomposition. The results were as follows: (1) There were correlations among the indexes of tree shape structure and among the indexes of leaf trait (P < 0.05). (2) The observed indexes of habitat elements had a strong influence on the growth indexes of B. ceiba leaves. The mean annual precipitation, slope and average temperature had a higher interpretation rate on the growth indexes of B. ceiba leaves, and were the dominant habitat indexes affecting the growth of B. ceiba. (3) According to the standardized path coefficient, the three habitat elements had the same effects on tree shape structure and leaf trait of B. ceiba, and all of them were in the order of topographic element>meteorological element>soil element. (4) The superposition of three habitat elements explained that tree shape structure and leaf trait were 43.5% and 12.3%, respectively, indicating that the response of tree shape structure to habitat environment was greater than that of leaf trait to environment. The results of this study preliminarily explore the adaptation strategies of different growth indexes to environmental elements of B. ceiba, which provides theoretical basis and practical reference for the cultivation and propagation of B. ceiba in heterogeneous habitats and efficient management of B. ceiba plantation.
Key words: Bombax ceiba, habitat elements, tree shape structure, leaf trait, redundancy analysis, structural equation model
近百年来,全球气候呈现显著的变暖趋势,使得全球平均气温、平均降水量都在发生改变(苑丹阳等,2020)。《IPCC全球升温1.5 ℃特别报告》指出,未来全球气温仍持续升高,干旱程度会加剧(IPCC, 2018),气候的变化使得树木生长受到不同程度的影响(Deslauriers et al., 2007),从而改变了森林生态系统的结构和功能(Bonan, 2008)。因此,在气候变化背景下,明确树木生长指标对环境因子的响应,有助于更好地了解森林生态系统的变化趋势,为植物生长、保护和管理提供理论依据。
目前,对植物功能性状的研究不再局限于对单一或成组性状水平的研究,而是越来越多地关注性状间的内在联系与权衡关系(Wright et al., 2004)。明晰植物功能性状与环境、生态系统的内在联系,有助于揭示植物的生长策略及资源分配模式,探讨其生理生态过程的内在机制,可以为研究生态系统的能量流动和物质循环提供更加可靠的背景数据以及更准确地预测气候变化对植物群落的影响(Wright et al., 2004; 何芸雨等,2019)。树形结构能够描述森林生长状况,反映其生长、竞争、更新等自然过程以及受人类活动的影响,其中,胸径和树高是预测群落发展趋势的两个指标(李晨笛等,2023)。木棉在其形态构建过程中往往会受到外部环境的影响。温度是影响树木树形结构的气候因子之一(Delpierre et al., 2019)。树木的径向生长常常受益于生长季早期温度的升高。一方面,树木径向生长所需的非结构性碳水化合物的产生依赖于叶片光合作用,而光合作用依赖于叶绿素和光合酶的作用,生长季温度的升高可加快光合酶的功能(Shi et al., 2015),从而可增加木质部生长所需的碳水化合物的累积。另一方面,肥沃的土壤能为植物的生长提供更多的营养物质(尤其是可利用性N、P),影响植物的生产力及其他生物学过程,促进树木生长。海拔梯度差异使得温度、湿度、光照等诸多环境因子发生剧烈变化,成为影响树木生长的一个重要指标(蔡礼蓉等,2017)。叶片作为植物的基本结构和功能单位,是植物、土壤、大气能量和物质交换的重要器官,叶片性状对环境变化的敏感性直接或间接影响着植物的生理生态过程。土壤养分是植物叶片性状构建过程中生长物质和能量的来源,海拔的变化会间接影响温度、降水、光照等因子产生再分配,导致气候环境的空间异质性,影响植物的生长发育并对植物功能性状的变化产生间接作用(Cornelissen et al., 2003)。由于环境的微小变化会使植物表现出不同的外部形态差异,因此探究环境要素对同种植物的不同性状影响具有重要意义。尽管已有学者对植物功能性状与环境关系进行了调查,但研究树种主要集中于胡杨(魏圆慧等,2021; 石义强等,2023)和锦鸡儿(罗源林等,2022; 杨建红等,2023)等少数树种。因此,为更准确地探究植物功能性状与环境之间的相关关系,还需要扩大对不同树种的研究。
本研究以云南省西双版纳国家级自然保护区内230棵木棉(Bombax ceiba)为研究对象,测定6个树形结构指标、8个叶性状指标以及所在区域的地形、气象、土壤养分等生境要素。通过相关性分析、冗余分析(redundancy analysis,RDA)、主成分分析(principal component analysis,PCA)、结构方程模型(structural equation model,SEM)分析以及变差分解(variation decomposition)检验研究,拟探讨以下问题:(1)树形结构各观测指标、叶性状各观测指标有无相关性;(2)不同生境观测指标对木棉生长影响差异;(3)树形结构和叶性状对同一生境不同生境要素响应差异。通过以上问题的研究,为实现木棉野外种群的建立和保护提供科学依据。
1 材料与方法
1.1 研究区概况
试验区位于云南省西双版纳国家级自然保护区(100SymbolpB@50′—101SymbolpB@06′ E、21SymbolpB@08′—22SymbolpB@25′ N),海拔在480~1 400 m之间,以山原盆地和山丘沟谷镶嵌交错为主。该区域温湿适宜,拥有充沛的光热水资源,全年降水雨量为1 200~1 600 mm,年平均气温为21.8 ℃,日照时数为2 293.4 h。西双版纳是中国热带雨林集中分布的重要区域,土壤类型主要由砖红壤与红壤组成,土壤呈酸性(马顺容等,2020),全磷、硝态氮含量较为丰富。保护区地处热带生物区系向亚热带生物区系过渡的生物地理群落交错带上(朱华等,2015),森林植被复杂,乔木层物种丰富,同时林下物种多样,形成了我国独特的热带雨林生态系统。主要伴生树种有相思树(Celtis timorensis)、大药树(Antiaris toxicaria)、耳草(Hedyotis auriclaria)、南山花(Prismatomeris connate)等。
1.2 木棉生长指标测定
通过野外实地调查,对西双版纳国家级自然保护区范围内自然生长状况下的230株木棉分别进行形态结构和生境要素的测定。
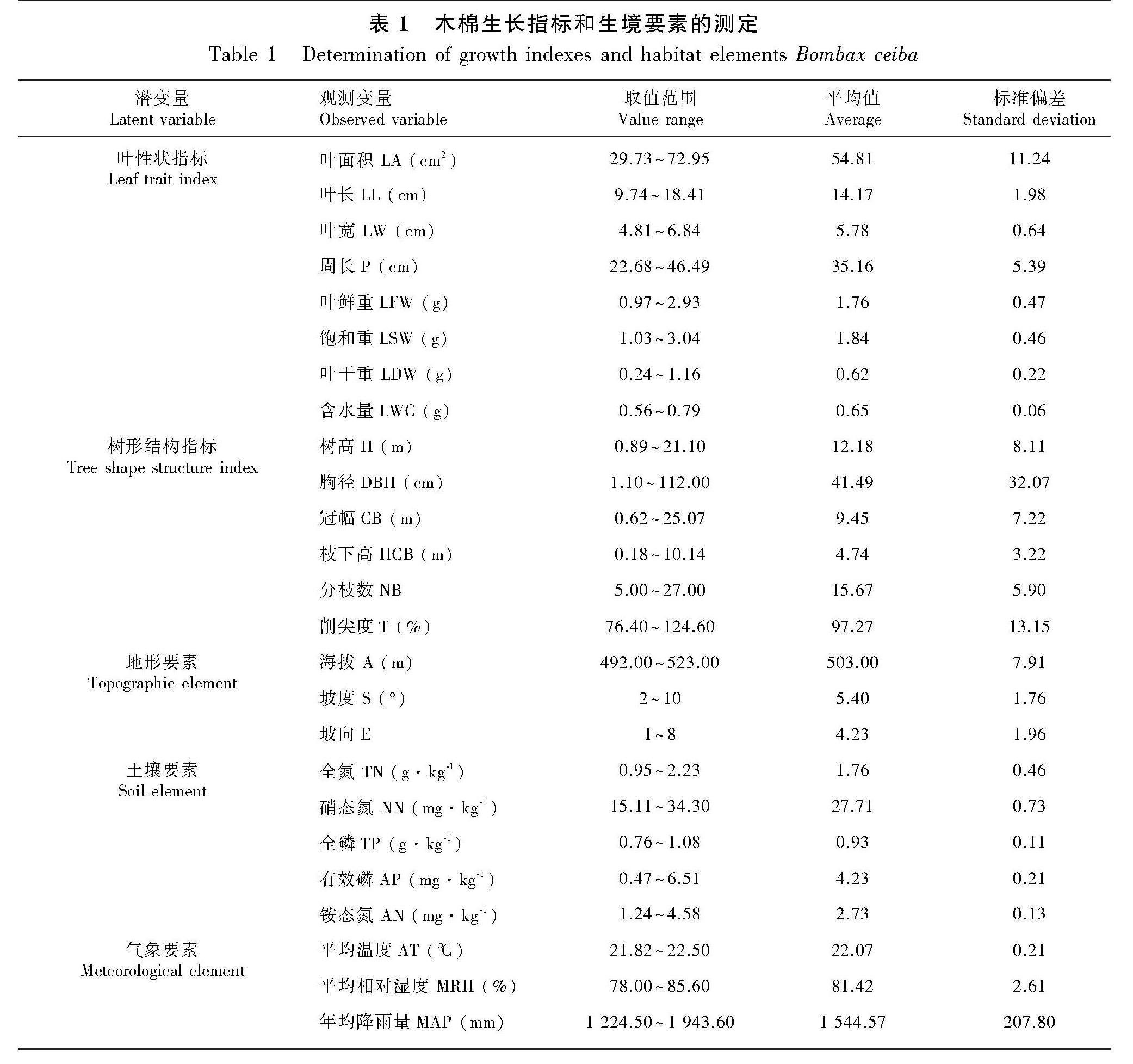
使用丈量尺测量每棵木棉的树高(height,H)、胸径(diameter at breast height,DBH)、冠幅(crown breadth,CB)、枝下高(height to crown base,HCB);从东南西北四个方位数分枝数(number of branches,NB);利用等比例关系求算出削尖度(taperingness,T)。每株木棉从东南西北四个方位的冠层中部外圈摘取充分伸展且健康完整的20枚叶片。利用CID便携式激光叶面积仪(CID CI-202,USA)测定每片叶的叶长(leaf length,LL)、叶宽(leaf width,LW)、叶面积(leaf area,LA)、周长(perimeter,P);通过岛津分析天平(ATY124,Japan)称其叶片的鲜重(leaf fresh weight,LFW)、饱和重(leaf saturated weight,LSW)、干重(leaf dry weight,LDW),其中叶片含水量(leaf water content,LWC)计算公式为LWC(%)=(LFWSymbolm@@LDW)/LDW×100。
1.3 生境要素测定
利用手持式GPS仪实测出每株木棉所在地的经纬度、海拔(altitude,A)、坡向(exposure,E)等数据,再利用地质罗盘测其坡度(slope,S)。对所测坡向进行数量化处理,坡向数据的测量以面朝东方开始,按顺时针方向旋转至东北方向赋值为1~8,后将各项观测指标进行标准化和归一化处理(陈莹等,2019)。根据5点取样法在木棉生长区域采取土样,均匀混合每样方同一层5个点表土层0~20 cm的土样放进布袋带回实验室,将土壤风干、研磨、过筛,带入实验室测定全氮(total nitrogen, TN)、全磷(total phosphorus, TP)、有效磷(available phosphorus, AP)、铵态氮(ammonium nitrogen, AN)、硝态氮(nitrate nitrogen, NN)的化学指标(刘蕾等,2021),5项指标均进行3次重复。气象数据来自中国地面气象站勐腊站(101°35′ E、21°28′ N),获取2009至2018年10年的平均气温(average temperature, AT)、平均相对湿度(mean relative humidity, MRH)、年均降雨量(mean annual precipitation, MAP)(htpp://data.cma.cn/),样地基本特征如表1所示。
1.4 数据处理
采用Excel 2016、SPSS 22.0、Amos 21.0和Canoco 5.0软件对数据进行统计分析。首先,在α=0.01的显著性水平下,采用Pearson法分别对木棉树形结构各指标、叶性状各指标进行相关分析。然后,对树形结构各指标、叶性状各指标、土壤要素、地形要素以及气象要素分别做主成分分析,利用第1主成分轴载荷因子除以相应主成分特征值的平方根作为各观测变量的系数。在Amos 21.0统计软件中使用结构方程模型拟合, 使用卡方值/自由度(Chi-square/df)进行拟合优度比较;CFI值和GFI值大于0.9以上为理想(董灵波等,2021)。最后,采用Canoco 5.0软件进行冗余分析(RDA)和变差分解,得到不同生境要素对木棉形态影响大小以及对木棉形态结构的解释率。
2 结果与分析
2.1 树形结构指标、叶性状指标相关性分析
皮尔逊相关分析表明,树高与胸径、冠幅、枝下高、分枝数呈极显著正相关,胸径与冠幅、枝下高、分枝数呈极显著正相关,冠幅与枝下高、分枝数呈极显著正相关,削尖度与树高、枝下高呈极显著负相关,整体上木棉树形结构各观测指标之间相关性较强。对叶性状各指标进行相关性分析可知,叶面积、叶长、叶宽与叶干重呈极显著负相关,与其余各指标呈极显著正相关;周长与叶干重呈显著负相关,与其他各指标呈极显著正相关;鲜重、饱和重、干重与含水量呈极显著负相关,与其他各指标呈极显著正相关(图1)。
2.2 生境要素、木棉指标权重分析
对3个生境要素指标和木棉形态各指标进行冗余分析(RDA)。前两轴分别解释总方差的60.52%和8.01%,共解释了68.53%,说明RDA的结果可靠。环境因子对木棉生长指标解释大小依次为年均降雨量(46.4%)>坡度(7.9%)>平均温度(5.7%)>海拔(4.3%)>全氮(1.9%)>有效磷(1.5%)>硝态氮(1.4%)>铵态氮(1.3%)>平均相对湿度(1.2%)>坡向(0.5%)(图2)。
应用主成分分析(PCA)可有效表征结构方程模型(SEM)中观测变量和对应潜变量的关系。计算3个生境要素以及两类木棉生长指标进行权重系数,发现树高、胸径、冠幅3个指标在树形结构权重中所占比重较大,其系数分别为0.241、0.287、0.316。叶面积、叶长、叶宽、叶片含水量在叶性状中权重占比较大,其系数分别为0.254、0.255、0.217、0.221。全氮、全磷、有效磷在土壤要素中所占权重较大,权重系数分别为0.252、0.243、0.250。平均相对湿度在气象要素中权重占比最大,系数为0.467,而坡度在地形要素中所占权重最大,系数为0.580(表2)。
2.3 生境要素对木棉树形结构、叶性状路径分析
利用结构方程模型(SEM)检验气象要素、土壤要素及地形要素对木棉树形结构与叶性状的效应, 最优模型与数据之间具有较高匹配度 (GFI=0.998;AGFI=0.968;SRMR<0.005)。木棉树形结构与叶性状呈极显著负相关(P<0.01),相关系数为-0.47。地形要素、气象要素和土壤要素三者相关性较强,呈极显著正相关(P<0.01)。由路径分析可得,地形要素、气象要素、土壤要素对木棉树形结构均为正向影响,其中地形要素和气象要素产生极显著正向影响(P<0.01),路径系数分别为0.47和0.39,土壤要素影响较小,路径系数为0.12。3个生境要素对木棉树形结构和叶性状影响存在差异,地形要素对叶性状具有极显著负向影响,路径系数为-0.32,而气象要素、土壤要素两者对木棉叶性状无显著影响(图3)。
2.4 生境要素对树形结构、叶性状解释分析
气象要素、地形要素和土壤要素是生境要素的重要组成部分,对植物的生长具有重要意义。利用变差分解分析量化地形要素、气象要素和土壤要素单独及交互作用对木棉树形结构与叶性状的贡献(图4)。结果表明,3个生境要素的总叠加作用解释了木棉树形结构的43.5%。其中,地形要素解释最大,为18.6%;其次是气象要素,解释了树形结构的17.5%;土壤要素解释度最小,仅为12.4%。相较于树形结构解释而言,土壤要素、地形要素以及气象要素对叶性状的总解释度低于树形结构,总解释度为12.3%。地形要素、土壤要素和气象要素3个生境要素对木棉树形结构解释度分别为6.0%、20.1%和4.7%。
3 讨论与结论
3.1 木棉树形结构对生境要素的响应
树形结构与环境生境要素具有较强的相关关系。本研究中,3个生境要素对树形结构具有较大影响,其中地形要素对木棉树形结构解释度最高,气象要素次之,而土壤要素解释最小。可见,在热带雨林中地形对木棉的生长影响最大。雷静品等(2012)探讨了不同海拔梯度上气候变化对树木径向生长的影响,结果显示随着海拔的变化限制青海云杉径向生长的气候因子从降水转变为温度,不同季节的限制因子也发生变化,与本文研究结果存在差异,原因可能是地形的变化引起木棉生长区水分和湿度产生变化,当海拔较低时木棉可以从地面汲取较多水分以及土壤养分,但随着海拔的升高,气温下降,大气压降低,光强增加,直接影响植物的代谢和调节过程,限制了树木的生长(Pan et al., 2009),故表现出地形解释较大的特点。同时,本研究结果与Hu和 Fan(2016)研究结果相似,即树干半径增量与相对湿度和降雨量呈正相关,而与日最高气温呈负相关。研究发现,木棉树高、胸径、冠幅3个生长指标具有显著正相关(P<0.01),应对环境时表现出一致性,这可能是木棉内部基因、外部环境两者共同作用所造成。王婷等 (2010)在伏牛山地区对华山松生长研究发现, 5、6月份气温升高,蒸发增强,容易造成土壤水分亏缺,进而影响华山松的生长,这一研究结果与本文研究结果存在差异,原因可能在于春夏之交的高温对华山松生长有显著的限制作用,而本研究区位于热带地区,不仅温度较高,而且水分充足,水分能够通过影响细胞的压缩进而影响细胞分裂和生长(Muller et al., 2011),较高的温度有利于增强木棉的光合作用,有利于生成更多的碳水化合物用于木棉树的生长,从而更好地进行树形结构的构建。
3.2 木棉叶性状对生境要素的响应
树木的叶性状不仅受到树木本身遗传因素的影响,同时还可能受到各种环境因子的调控(Rathgeber et al., 2016)。研究发现,土壤要素对叶性状解释度最大,地形要素其次,而气象要素最小。同时,路径分析表明地形要素与气象要素对木棉叶性状具有负向影响且地形要素影响大于气象要素。而木棉叶面积、周长、叶片含水量、叶宽、叶长与硝态氮、全氮、有效磷呈正相关,反映出木棉生长过程中将更多土壤养分用于叶片性状构建,本研究结果与黄磊等(2021)对杉木生长与土壤养分的关系研究一致,表明当土壤养分越高(如本研究的有效磷)的提高,木棉叶性状倾向于有更大的周长、叶面积和更高的比叶面积。因此,在养分更充足的情况下,西双版纳的木棉倾向于快速投资型生长策略,本研究结果与很多研究结果相似(罗达等,2021; 刘思文等,2021; 黄郑雯等,2022)。气象要素对叶性状影响会因环境差异而变化。在本研究中,叶面积、周长、叶片含水量、叶长、叶宽与年均降水量、平均相对湿度呈正相关,与平均气温呈负相关。这表明空气湿度与土壤湿度对木棉叶性状具有促进作用,而高温对叶性状起到抑制作用,表现为温度升高会使叶片气孔关闭,减少光合作用,进而影响木棉生物量的积累,该位置处于热带季风气候影响区域,4月中旬正值当地旱季(枯水期后期),光照充足,温度较高对叶片影响较显著,体现为高温对叶性状的抑制,但树形结构相较于叶性状而言,对于环境变化敏感度较小,夏季温度升高对自身养分传输与形态构建影响不大,使得在标准化路径系数中表现为正向影响,研究结果与前人研究结果一致,即干旱缺水环境对植物生长具有抑制作用(施钦等,2019)。
3.3 木棉树形结构与叶性状的相关性及其对环境条件的适应策略
植物形态结构能够客观表达植物对外部环境的适应(孟婷婷等,2007)。大尺度上,气候要素对植物功能性状的分布起决定性作用;在中等尺度上,土地利用和干扰起主要作用;在小尺度或局地范围内,地形要素和土壤要素决定性状的表达(Venn et al., 2011)。本研究表明,气象要素、地形要素以及土壤要素三者具有极显著相关关系(P<0.01),但对木棉形态结构表现出不同影响,其中地形要素对树形结构和叶性状影响最大,其次为气象要素,土壤要素影响最小。这表明木棉生长指标受种源地的温度、水分以及土壤养分共同作用,而不是由温度或水分单独决定(Barboni et al., 2004)。气象要素决定着木棉生长所需的温度和水分,海拔、坡度和坡向等地形要素则通过影响水、热、光的分配进而改变区域温、湿状况,而土壤受到温度、降水、生物活动以及地质变化等影响,能够营造适合植物生长的物质层,该层蕴含植物生长所需的养分、能量以及合适的土壤酸碱性,最终影响其生长(Huang et al., 2018; 刘宪钊等,2021)。本研究发现,树形结构与叶性状具有极显著负相关关系,而3个生境要素对树形结构解释度(43.5%)高于叶性状(12.3%),表明木棉在不同生长阶段会依据自身需求进行资源分配。本研究结果与前人研究一致,当木棉处于幼苗发育阶段时,为使其自身的快速生长获得营养,通过增大叶面积、比叶面积进而增强捕获光合能力,同时吸收大量水分,满足其旺盛生长活动,以此提高幼苗阶段生存力(杨巧等,2022)。当木棉进入生长稳定期或停滞期时,通过减小叶面积、比叶面积来降低植物的蒸腾作用,减少叶片消耗过多的营养以及水分;叶片含水率的减少有利于木棉发育稳定期间代谢的减少,保障积累更多资源,有效保障木棉树形结构扩展,对于木棉构建优势种群提供便利条件(Wright et al., 2002; 丁曼等,2014; 刘媛霜等,2020)。当然,木棉生长指标不仅包括树形结构与叶性状,能集中反映对光照吸收和水分汇集的年轮性状以及枝条与主干的夹角等在生长过程中具有重要参与作用,未来应该进一步增加更多生长指标对环境要素响应的研究,使其更具合理性。
本研究结果表明,木棉树形结构各指标、叶性状各指标广泛且半数以上可达到极显著相关水平,体现出同一生境下木棉树形结构、叶性状自身调整具有相似性的特征。3个环境要素对木棉树形结构与叶性状影响存在差异,表现出对树形结构影响更大的特点。当然,木棉在生长过程中为应对生境环境的变化,树形结构与叶性状之间采取负相关关系,这是树形结构与叶性状形成不同组合来应对环境的变化结果。
参考文献:
BARBONI D, HARRISON SP, BARTLEIN PJ, et al., 2004. Relationships between plant traits and climate in the mediterranean region: a pollen data analysis [J]. J Veg Sci, 15(5): 635-646.
BONAN GB, 2008. Forests and climate change: forcings, feedbacks, and the climate benefits of forests [J]. Science, 320: 1444-1449.
CAI LR, KUANG X, FANG S, et al., 2017. Factors influencing tree radial growth of three common species in broad-leaved Korean pine mixed forests in Changbai Mountains,China. [J]. Chin J Appl Ecol, 28(5): 1407-1413. [蔡礼蓉, 匡旭, 房帅, 等, 2017. 长白山阔叶红松林3个常见树种径向生长的影响因素 [J]. 应用生态学报, 28(5): 1407-1413.]
CHEN Y, DONG LB, LIU ZG, 2019. Optimal species composition for the main forest types of secondary forest in Maoershan Mountain, northeastern China [J]. J Beijing For Univ, 41(5): 118-126. [陈莹, 董灵波, 刘兆刚, 2019. 帽儿山天然次生林主要林分类型最优树种组成 [J]. 北京林业大学学报, 41(5): 118-126.]
CORNELISSEN JHC, LAVOREL S, GARNIER E, et al., 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 51 (4): 335-380.
DELPIERRE N, LIREUX S, HARTIG F, et al., 2019. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers [J]. Glob Chang Biol, 25(3): 1089-1105.
DESLAURIERS A, ANFODILLO T, ROSSI, et al., 2007. Using simple causal modeling to understand how water and temperature affect daily stem radial variation in trees [J]. Tree Physiol, 27(8): 1125-1136.
DING M, WEN ZM, ZHENG Y, 2014. Scale change and dependence of plant functional traits in hilly areas of the loess region [J]. Acta Ecol Sin, 34(9): 2308-2315. [丁曼, 温仲明, 郑颖, 2014. 黄土丘陵区植物功能性状的尺度变化与依赖 [J]. 生态学报, 34(9): 2308-2315.]
DONG LB, TIAN DY, CHEN Y, et al., 2021. Clarifying the factors affecting Larix gmelinii forest regeneration based on structural equation model [J]. Chin J Appl Ecol, 32(8): 2763-2772. [董灵波, 田栋元, 陈莹, 等, 2021. 基于结构方程模型的兴安落叶松天然林更新影响因素 [J]. 应用生态学报, 32(8): 2763-2772.]
HE YY, GUO SL, WANG Z, 2019. Research progress of trade-off relationships of plant functional traits [J]. Plant Sci J, 43(12): 1021-1035. [何芸雨, 郭水良, 王喆, 2019. 植物功能性状权衡关系的研究进展 [J]. 植物生态学报, 43(12): 1021-1035.]
HU LF, FAN ZX, 2016. Stem radial growth in response to microclimate in an asian tropical dry karst forest [J]. Acta Ecol Sin, 36(5): 401-409.
HUANG JG, GUO X, ROSSI S, et al., 2018. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season [J]. Tree Physiol, 38(8): 1225-1236.
HUANG L, WANG G, YANG B, et al., 2021. Relationship between Chinese fir timber, topography and soil nutrient [J]. J Fujian Agric For Univ (Nat Sci Ed), 50(5): 619-623. [黄磊, 王港, 杨冰, 等, 2021. 杉木大径材成材与地形、土壤养分的关系 [J]. 福建农林大学学报(自然科学版), 50(5): 619-623.]
HUANG ZW, YANG L, WANG YJ, et al., 2022. The characteristics of tree shape structure and the influencing factors of Bombax ceiba L. in different habitats [J]. Chin J Ecol, 41(8): 1552-1559. [黄郑雯, 杨霖, 王玉洁, 等, 2022. 不同生境下木棉树形结构特征及其影响因子 [J]. 生态学杂志, 41(8): 1552-1559.]
IPCC, 2018. Global warming of 1.5 ℃ IPCC special report [R]. Korea: Inchon. [IPCC, 2018. 2018年IPCC全球升温1.5 ℃特别报告 [R]. 韩国: 仁川. ]
KOSAKA K, LAMONT BB, LEE T, et al., 2004. The worldwide leaf economics spectrum [J]. Nature, 428: 821-827.
LEI JP, FENG XH, SHI Z, et al., 2012. Stability of relationship between climate and Picea crassifolia radial growth in different elevations [J]. Acta Bot Boreal-Occident Sin, 32(12): 2518-2529. [雷静品, 封晓辉, 施征, 等, 2012. 海拔梯度上青海云杉径向生长与气候关系稳定性研究 [J]. 西北植物学报, 32(12): 2518-2529.]
LI CD, YANG XB, LI DH, et al., 2023. Changes of community structure and diversity of natural forests in the central area of Hainan Island [J]. Chin J Ecol, 42(3): 513-523. [李晨笛, 杨小波, 李东海, 等, 2023. 海南中部山区天然林群落结构与多样性变化 [J]. 生态学杂志, 42(3): 513-523.]
LIU L, XU M, ZHANG GY, et al., 2021. Seasonal variations of arbuscular mycorrhizal fungal community in greenhouse soil under different rotation systems [J]. Chin J Appl Ecol, 32(11): 4095-4106. [刘蕾, 徐梦, 张国印, 等, 2021. 不同轮作模式下设施土壤丛枝菌根真菌群落结构的季相变化 [J]. 应用生态学报, 32(11): 4095-4106.]
LIU SW, AI YB, LIU YH, 2021. Variations in leaf functional traits along the altitude gradient of Pinus tabuliformis and its environmental explanations in Beijing Songshan Mountain [J]. J Beijing For Univ, 43(4): 47-55. [刘思文, 艾也博, 刘艳红, 2021. 北京松山油松叶功能性状沿海拔梯度的变化及其环境解释 [J]. 北京林业大学学报, 43(4): 47-55.]
LIU XZ, HAN WB, GAO RD, et al., 2021. Potential impacts of environmental types on geographical distribution of Larix principis-rupprechtiu [J]. Atca Ecol Sin, 41(5): 1885-1893. [刘宪钊, 韩文斌, 高瑞东, 等, 2021. 不同环境类型对华北落叶松分布的潜在影响 [J]. 生态学报, 41(5): 1885-1893.]
LIU YS, WAN JN, ZHOU YD, et al., 2020. Research progress on Adansonia [J]. Plant Sci J, 38(4): 558-564. [刘媛霜, 万俊男, 周亚东, 等, 2020. 猴面包树属植物的研究进展 [J]. 植物科学学报, 38(4): 558-564.]
LUO D, SHI YJ, SONG FH, et al., 2021. Variation and correlation of leaf functional traits and photosynthetic characteristics of 38 hazelnut germplasm resources [J]. Chin J Ecol, 40(1): 11-22. [罗达, 史彦江, 宋锋惠, 等, 2021. 38个榛种质资源叶功能性状与光合特征变异及其相关性 [J]. 生态学杂志, 40(1): 11-22.]
LUO YL, MA WH, ZHANG XY, et al., 2022. Variation of functional traits of alternative distribution of Caragana species along environmental gradients in Nei Mongol, China [J]. Chin J Plant Ecol, 46(11): 1364-1375. [罗源林, 马文红, 张芯毓, 等, 2022. 内蒙古锦鸡儿属植物地理替代分布种的功能性状沿环境梯度的变化 [J]. 植物生态学报, 46(11): 1364-1375.]
MA SR, LIN YJ, LU TP, et al., 2020. Effects of exogenous phosphorus addition on soil ecological stoichiometry in Xishuangbanna tropical rainforest [J]. Chin J Ecol, 39(10): 3194-3202. [马顺容, 林永静, 卢同平, 等, 2020. 外源磷添加对西双版纳热带雨林土壤生态化学计量特征的影响 [J]. 生态学杂志, 39(10): 3194-3202.]
MENG TT, NI J, WANG GH, 2007.Plant functional traits, environments and ecosystem functioning [J]. Chin J Plant Ecol, 31(1): 150-165. [孟婷婷, 倪健, 王国宏, 2007. 植物功能性状与环境和生态系统功能 [J]. 植物生态学报, 31(1): 150-165.]
MULLER B, PANTIN F, GENARD M, et al., 2011. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs [J]. J Exp Bot, 62(6): 1715-1729.
PAN HL, LI MH, CAI XH, et al., 2009. Responses of growth and ecophsiology of plants to altitude [J]. Ecol Environ Sci, 18(2): 722-730.
RATHGEBER CB, CUNY HE, FONTI P, 2016. Biological basis of tree-ring formation: A crash course [J]. Front Plant Sci, 7: 734.
SHI CG, SILVA LCR, ZHANG HX, et al., 2015. Climate warming alters nitrogen dynamics and total non-structural carbohydrate accumulations of perennial herbs of distinctive functional groups during the plant senescence in autumn in an alpine meadow of the Tibetan Plateau, China [J]. Agric For Meteorol, 200: 21-29.
SHI Q, BAO XW, HUA JF, et al., 2019. Effects of drought stress and recovery on photosynthesis and physiological characteristics of Hibiscus hamabo [J]. Chin J Appl Ecol, 30(8): 2600-2606. [施钦, 包学文, 华建峰, 等, 2019. 干旱胁迫及复水对海滨木槿光合作用和生理特性的影响 [J]. 应用生态学报, 30(8): 2600-2606.]
SHI YQ, ABULIZI R, HALIK U, et al., 2023. Differences in leaf functional traits of Populus euphratica at different growth stages and its relationship with tree shape factors [J]. J For Environ, 43(1): 1-7. [石义强, 热孜也木·阿布力孜, 玉米提·哈力克, 等, 2023. 胡杨叶功能性状差异及其与树形因子的关系 [J]. 森林与环境学报, 43(1): 1-7.]
VENN SE, GREEN K, PICKERING CM, et al., 2011. Using plant functional traits to explain community composition across a strong environmental filter in australian alpine snow patches [J]. Plant Ecol, 212(9): 1491-1499.
WANG T, SHEN LF, YE YZ, et al., 2010. Response analysis between climate chang and tree-ring widths of Pinus armandi in Funiu Mountain [J]. Henan Sci, 28(12): 1549-1551. [王婷, 沈连峰, 叶永忠, 等, 2010. 伏牛山华山松径向生长对气候变化的响应 [J]. 河南科学, 28(12): 1549-1551.]
WEI YY, LIANG WZ, HAN L, et al., 2021. Leaf functional traits of Populus euphratica and its response to groundwater depths in Tarim extremely arid area [J]. Acta Ecol Sin, 41(13): 5368-5376. [魏圆慧, 梁文召, 韩路, 等, 2021. 胡杨叶功能性状特征及其对地下水埋深的响应 [J]. 生态学报, 41(13): 5368-5376.]
WRIGHT IJ, REICH PB, WESTOBY M, et al., 2004. The world wide leaf economics spectrum [J]. Nature, 428(6985): 821-827.
WRIGHT IJ, WESTOBY M, REICH PB, 2002. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span [J]. J Ecol, 90(3): 534-543.
YANG JH, ZHAO CY, LI N, et al., 2023. Temporal variation of leaf functional traits of dominant shrub species in the middle section of Qilian Mountains [J].Acta Ecol Sin, 43(2): 709-718. [杨建红, 赵传燕, 李娜, 等, 2023. 祁连山中部优势灌木物种叶功能性状参数变化特征 [J]. 生态学报, 43(2): 709-718.]
YANG Q, ZHU RJ, YANG CY, et al., 2022. Variation in leaf functional traits of Bombax ceiba Linnaeus communities based on tree structure [J]. Acta Ecol Sin, 42(7): 2834-2842. [杨巧, 朱润军, 杨畅宇, 等, 2022. 基于树形结构的木棉叶功能性状差异性研究 [J]. 生态学报, 42(7): 2834-2842.]
YUAN DY, ZHAO HY, LI ZS, et al., 2020. Radial growth of Pinus koraiensis and Picea koraiensis response to climate change in Yichun City, Heilongjiang Province [J]. Acta Ecol Sin, 40(4): 1150-1160. [苑丹阳, 赵慧颖, 李宗善, 等, 2020. 伊春地区红松和红皮云杉径向生长对气候变化的响应 [J]. 生态学报, 40(4): 1150-1160.]
ZHU H, WANG H, LI BG, et al., 2015. Studies on the forest vegetation of Xishuangbanna [J]. Plant Sci J, 33(5): 641-726. [朱华, 王洪, 李保贵, 等, 2015. 西双版纳森林植被研究 [J]. 植物科学学报, 33(5): 641-726.]
(责任编辑 周翠鸣)
