份香蕉种质资源SRAP分子标记亲缘关系分析
2024-06-09周海琪夏玲吕顺曾莉莎王芳黄晓彦陈东仪刘文清梁少丽刘丽琴
周海琪 夏玲 吕顺 曾莉莎 王芳 黄晓彦 陈东仪 刘文清 梁少丽 刘丽琴

关键词:香蕉;SRAP;聚类分析;亲缘关系
中图分类号:S668.1 文献标志码:A
香蕉是目前世界上产量和贸易量最大的水果之一,也是热带、亚热带地区重要的经济作物和粮食作物[1]。香蕉味道香甜,营养丰富,物美价廉,深得人们喜爱,而且香蕉植株优美,也常被人们用来装点环境。我国是香蕉的原产地之一,具有丰富的种质资源,同时也具有悠久的栽培历史。目前香蕉的栽培品种主要有香牙蕉、龙牙蕉、贡蕉、粉蕉和大蕉,这些品种基本由尖叶蕉(Musaacuminata Colla,A 基因组)和长梗蕉(Musabalbisiana Colla,B 基因组)这2 个野生品种发展而来[2],染色体倍性包含二倍体、三倍体(主要)和四倍体。由于来源、地域、育种手段等因素,不同的香蕉品种间的遗传背景存在差异。香蕉一直沿用SIMMONDS[3]的分类方法来识别和分类不同的香蕉品种,但是随着研究的深入和研究技术手段的更新,这种单一的、依靠形态特征的分类方法已经无法满足对香蕉亲缘关系和遗传多样性等研究的需求,甚至对研究造成一定的阻碍,如香蕉品种的分类分组与基因组组成不完全一致[4],影响香蕉种质资源的合理利用。因此,结合分子标记技术鉴定香蕉种质资源、分析其亲缘关系和研究遗传多样性是非常必要的。
国内外关于运用分子标记技术来研究香蕉种质的报道众多,SSR、AFLP、ISSR、RAPD、SRAP等分子标记[5-9]被应用在分析香蕉品种(系)的种群鉴定与分类、遗传多样性等方面。SRAP 技术是一种基于PCR 的分子标记技术[10],具有简便、快速、引物设计简便、多态性和信息量丰富等特点,广泛应用于园艺植物遗传育种[11]、作物改良[12]、果树种质资源鉴定和亲缘关系分析[13-17]等方面。谢子四等[18]利用SRAP标记对29 份香蕉材料进行了基因组多态性分析,扩增出324 条可辨认条带,多态性比率为80.86%;漆艳香等[9]应用SRAP 标记对21 份香蕉种质的遗传多态性进行分析。但这些报道都以少量或地方代表的个别品种为试验材料,不够系统和全面。本研究采用SRAP 分子标记技术,以89 份来至国内外的香蕉种质,包括香牙蕉、龙牙蕉、贡蕉、大蕉、粉蕉和不同类型的野蕉等多种香蕉类型为材料,探讨各类型、品种之间的亲缘关系,为香蕉育种、品种鉴定、新品种保护等提供分子水平上的依据。
1 材料与方法
1.1 材料
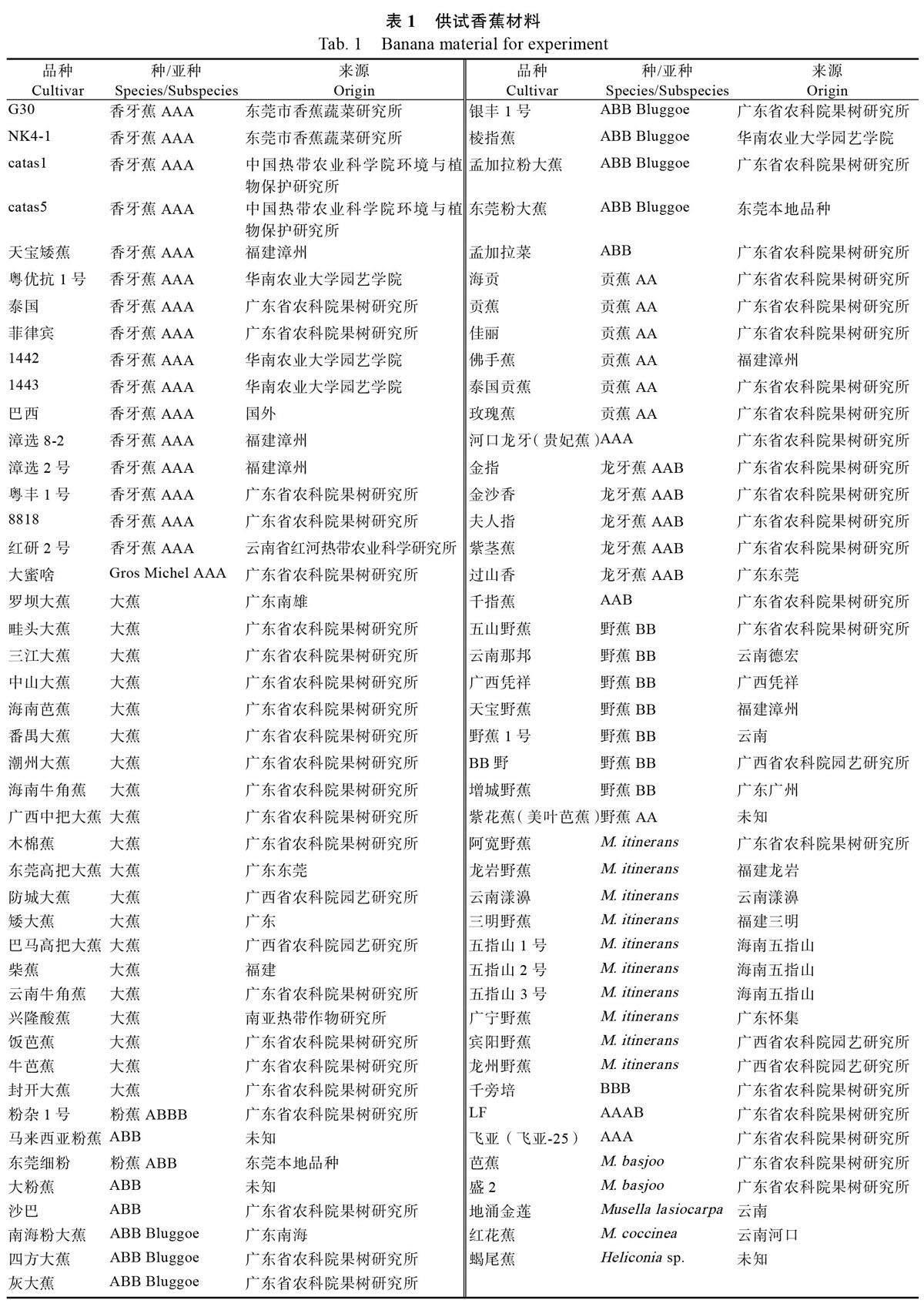
供试89 份材料取自东莞市农业科学研究中心的香蕉种质资源圃和麻涌试验基地,89 份香蕉种质资源样品的基因型和来源见表1,SRAP 引物序列由英潍捷基(上海)贸易有限公司合成,DL2000 DNA marker、dNTPs、Taq DNA 聚合酶,10×Buffer 等均购自TranGene 公司。
1.2 方法
1.2.1 香蕉基因组DNA 的提取及检测 取香蕉幼嫩的新叶,采用改良的CTAB 法[19]提取DNA,经蛋白核酸检测仪检测DNA 浓度与纯度后,用0.8%琼脂糖凝胶电泳检测DNA 质量,最后将DNA 稀释至所需浓度置于?20 ℃冰箱贮存备用。
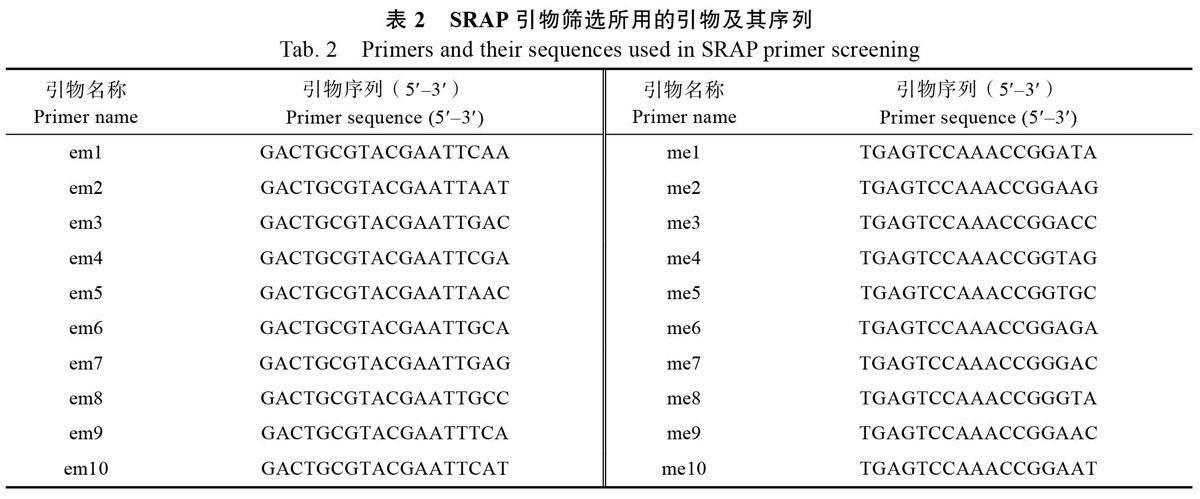
1.2.2 SRAP 引物的筛选及PCR 扩增 运用SRAP 分子标记引物(10 个正向引物和10 个反向引物两两组合成100 对引物组合,表2),以5 份香蕉品种为模板,对100 对引物进行筛选,每个引物重复2~3 次,筛选能扩增出多态性好且条带多的最适合引物组合。PCR 扩增反应在Bio-lePCR 扩增仪上进行,反应体系:20 μL 的PCR 反应体系含有10×buffer (含Mg2+)2 μL,0.2 mmol/LdNTPs(2.5 nmol/L)2 μL,上、下引物(10 μmol/L)各1 μL,DNA(20 ng/L)2 μL 及Taq DNA 聚合酶1 μL。反应程序为94 ℃预变性5 min;94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1 min,5 个循环;94 ℃变性1 min,50 ℃退火1 min,72 ℃延伸1 min,循环35 次;72 ℃延伸10 min。扩增反应结束后,加入4 μL 6×loading buffer,取8~10 μL 扩增产物在1.8%琼脂糖凝胶中电泳,电极缓冲液为0.5×TBE,在凝胶成像仪中观察并照相。
1.3 数据处理
经过2~3 次重复,统计稳定可靠的多态性位点,扩增条带在同一位置不同样品中,有条带记为“1”,无条带记为“0”,建立(0,1)矩阵,用Ntsys 软件采用UPGMA(unweghed pair methodairthtnetic average)进行聚类分析,构建树状图。
2 结果与分析
2.1 89 份香蕉种质资源基因组DNA 的检测
将提取的89 份香蕉样品基因组DNA 采用0.8%琼脂糖凝胶电泳进行检测,结果显示香蕉基因组DNA 主带明显,呈现一条完整发亮的谱带,无降解拖尾现象。经核酸蛋白仪检测后,所得DNA母液OD260/OD280 在1.8 左右,OD260/OD230≥2.0,浓度在1000~1800 ng/μL 之间,表明所提取的香蕉基因组DNA 完整性较好,纯度高,無小分子物质和蛋白质的污染,能够满足后续试验要求。将DNA 分装至PCR 管中备用,用时稀释至30 ng/μL。
2.2 香蕉SRAP 引物筛选及PCR 扩增
利用表2 所列的正反引物各10 条,共100 对引物组合进行筛选,以其中扩增条带易于识别、带型清晰、多态性高的13 对引物对供试材料进行分析。结果表明,13 对引物共扩增出170 条条带,平均每对引物13.08 条,其中多态性带共140 条(图1,图2)。各引物产生的多态性带比例为64.3%~92.9%,平均为79.0%(表3)。
2.3 遗传相似性系数分析
在遗传相似系数矩阵中,89 份香蕉种质资源的相似系数的变化范围在0.241~1.000 之间,在这些香蕉种质中,8818 与天宝野蕉、8818 与罗坝大蕉、罗坝大蕉与夫人指、畦头大蕉与红研2 号、矮大蕉与粤丰1 号的相似系数最低,为0.241。海贡和贡蕉相似性系数最高,为1.000,其SRAP 分子标记带型基本一致,但在田间表现中,海贡和贡蕉是有一定差异的,海贡具有枯萎病抗性,而贡蕉是感病品种。究其原因一方面可能我们采用的SRAP 分子标记位点还比较少,不足以区分这2 个品种,另一方面也有可能样品混淆,需要进一步实验证明。此外,分别来自于云南和广西的云南那邦和广西凭祥相似性系数为0.983,同时来自于云南的野蕉1 号与这2 个种质资源的相似性系数也在0.900 以上,这3 个品种均为BB 类野蕉,亲缘关系较近。五指山1 号和五指山2 号相似性系数为0.983,五指山3 号与五指山1 号和2 号的相似性系数相对较低,分别是0.81 和0.828,亲缘关系也比较近,五指山1 号、五指山2 号及五指山3 号是课题组在海南五指山不同地点采的样品,均为阿宽蕉类野生蕉。可见这些种质资源的遗传多样性非常丰富,香蕉种质资源的亲缘关系很复杂,而并不是简单的平行关系。
2.4 亲缘关系聚类分析
采用NTSYSpc 2.10 t 软件对SRAP 标记统计后的综合结果进行数据分析,建立树状图(图3)。在相似性系数为0.49 时,可以将89 份香蕉种质资源分为六大类群。
第一大类群:主要包括香牙蕉、贡蕉、龙牙蕉、AA 型野蕉,基因型主要为AAA 型、AAB型和AA 型,共33 份种质资源,在相似性系数为0.65 处可以将此大类分为四大亚类。第一亚类包括G30、NK4-1、海贡、贡蕉、河口龙牙(贵妃蕉)、夫人指、1442、漳选2 号、漳选8-2、1443、8818、巴西、天宝矮蕉、大蜜舍、泰国、菲律宾、粤优抗1 号、泰国贡蕉、catas1、佳丽、LF、飞亚(飞亚-25)、粤丰1 号、catas5、金指、紫茎蕉、金沙香、玫瑰蕉、红研2 号;第二亚类包括佛手蕉、过山香;第三亚类是千指蕉;第四亚类是紫花蕉(美叶芭蕉)。
第二大类群:主要包括粉蕉、粉大蕉、BB型野蕉,基因型主要为ABB 型,共21 份种质资源;在相似性系数为0.64 处可以将此大类分为二大亚类。第一亚类包括粉杂1 号、孟加拉粉大蕉、千蒡培、灰大蕉、增城野蕉、云南那邦、广西凭祥、野蕉1 号、孟加拉菜、五山野蕉、马来西亚粉蕉、BB 野蕉、南海粉大蕉、东莞粉大蕉、银丰1 号、棱指蕉、四方大蕉、大粉蕉;第二亚类包括天宝野蕉、东莞细粉、沙巴。
第三大类群:主要包括大蕉、阿宽蕉类野蕉以及芭蕉类野蕉,共32 份种质资源。在遗传距离为0.64 处可以将此大类分为五大亚类。第一亚类包括云南牛角蕉;第二亚类包括矮大蕉、罗坝大蕉、饭芭蕉、三江大蕉、番禺大蕉、广西中把大蕉、封开大蕉、东莞高把大蕉、兴隆酸蕉、木棉蕉、牛芭蕉、中山大蕉、潮州大蕉、海南芭蕉、柴蕉、巴马高把大蕉、五指山1 号、五指山2 号、五指山3 号、防城大蕉、海南牛角蕉、畦头大蕉;第三亚类包括阿宽野蕉、龙岩野蕉、三明野蕉、云南漾濞、广宁野蕉;第四亚类包括宾阳野蕉、龙州野蕉;第五亚类包括盛2、芭蕉。
第四、五、六类群:都只包含了1 份种质资源,分别是红花蕉、地涌金莲、蝎尾蕉。
3 讨论
香蕉种质资源丰富,种类品种繁多,然而由于特殊的生物学特性如多倍体、无性繁殖、营养结实、小染色体等[18],使其种质资源的分类及系统学研究一直是个难题。本研究以香牙蕉、龙牙蕉、大蕉、粉蕉和不同类型的野蕉,基因型主要包括AA、BB、AAA、AAB、ABB 和AAAA 等类型的89 份香蕉种质资源为材料,运用SRAP 分子标记技术研究不同香蕉种质资源之间的亲缘关系。由10 个正向引物和10 个反向引物两两组合成100 对引物组合,从中筛选出13 对扩增条带易于识别、带型清晰、多态性高的SRAP 引物,这13 对引物在89 个香蕉种质资源间扩增出170 条可辨认条带,其中表现出多态性的带有140 条,多态性比率为79.00%。
在相似性系数为0.49 时,可以将这89 份香蕉种质资源分成六大类,包括:香牙蕉、贡蕉、龙牙蕉、AA 型野蕉为主的第一大类(AAA、AA、AAB 基因型型),粉大蕉、粉蕉和BB 型野蕉为主的第二大类(ABB、BB 基因型),大蕉、阿宽蕉类野蕉为主的第三大类,红花蕉、地涌金莲、蝎尾蕉、分别所属的第四、五、六类群,该方法的分类结果与传统的基于形态的分类结果基本一致,说明用SRAP 标记对香蕉种质资源进行分类是可行的。但该研究中发现部分香蕉种质的分类分组并没有与品种的基因组组成一致,例如,以巴西为代表的AAA 型香牙蕉、以贡蕉为代表的AA 型及以龙牙蕉为代表的AAB 型聚类群体并不明显,均分散在第一类。三者在遗传上没有太大的差异,说明栽培类香牙蕉、贡蕉和龙牙蕉具有相同的祖先,与AA 基因型野蕉亲缘关系近,这与宁淑萍等[20]利用SSR 技术分析的结果一致。粉蕉及粉大蕉(ABB 型)聚为一类,可见二者遗传背景相似,与BB 基因型的野蕉亲缘关系较近,但无法明显区别是否含A 基因组,这与郭计华[21]的研究结果相似。不同基因组成的香蕉品种聚到一起,也说明香蕉品种的亲缘关系可能与起源有关。前人利用细胞遗传学方法[22]、RFLP 分子标记[23-24]和SSR分子标记[25-26]方法对香蕉品种资源进行分类分析,也发现与应用传统SIMMONDS[3]分类法命名的基因组型结果不一致。香牙蕉(AAA)、大蕉(ABB)、粉蕉(ABB)和龙牙蕉(AAB)都是根据形态学性状分类而笼统地确定基因组类型,难免会造成同物异名、同名异物的现象[27],也说明使用单一的分类方法无法明确地把香蕉种质区分,还需要从形态学、细胞学、分子生物学方面作进一步的研究和鑒定。
早期研究认为大蕉基因组为ABB 或者BBB,项目组前期研究发现大蕉倍性值(或DNA 相对含量)平均高于其他ABB 基因型资源,而且利用大蕉与长梗蕉人工杂交的后代均为四倍体,而粉蕉与长梗蕉的杂交后代为三倍体或四倍体[28],说明大蕉与ABB 基因型资源有所差别。但是,项目组在进行B 基因组相关的SCAR 标记的研究中又发现大蕉是含有A 和B 基因组的[29]。在本研究中21 份大蕉和10 份阿宽蕉聚到同一类,说明大蕉和阿宽蕉亲缘关系较近,但是从整个聚类图中无法看出大蕉是否含有A 和B 基因组,可见,大蕉的遗传背景及起源比较复杂,有待进一步研究。
