Transcriptomic and metabolomic analysis provides insights into lignin biosynthesis and accumulation and differences in lodging resistance in hybrid wheat
2024-05-13WeibingYangShengquanZhangQilingHouJiangangGaoHanxiaWangXianchaoChenXiangzhengLiaoFengtingZhang2ChangpingZhao2ZhilieQin
Weibing Yang ,Shengquan Zhang ,Qiling Hou ,Jiangang Gao ,Hanxia WangXianchao ChenXiangzheng LiaoFengting Zhang2,Changping Zhao2#,Zhilie Qin#
1 Institute of Hybrid Wheat,Beijing Academy of Agriculture and Forestry Sciences,Beijing 100097,China
2 Beijing Municipal Key Laboratory of Molecular Genetics of Hybrid Wheat,Beijing 100097,China
Abstract The use of hybrid wheat is one way to improve the yield in the future.However,greater plant heights increase lodging risk to some extent.In this study,two hybrid combinations with differences in lodging resistance were used to analyze the stem-related traits during the filling stage,and to investigate the mechanism of the difference in lodging resistance by analyzing lignin synthesis of the basal second internode (BSI).The stem-related traits such as the breaking strength,stem pole substantial degree (SPSD),and rind penetration strength (RPS),as well as the lignin content of the lodging-resistant combination (LRC),were significantly higher than those of the lodgingsensitive combination (LSC).The phenylpropanoid biosynthesis pathway was significantly and simultaneously enriched according to the transcriptomics and metabolomics analysis at the later filling stage.A total of 35 critical regulatory genes involved in the phenylpropanoid pathway were identified.Moreover,42% of the identified genes were significantly and differentially expressed at the later grain-filling stage between the two combinations,among which more than 80% were strongly up-regulated at that stage in the LRC compared with LSC.On the contrary,the LRC displayed lower contents of lignin intermediate metabolites than the LSC.These results suggested that the key to the lodging resistance formation of LRC is largely the higher lignin synthesis at the later grain-filling stage.Finally,breeding strategies for synergistically improving plant height and lodging resistance of hybrid wheat were put forward by comparing the LRC with the conventional wheat applied in large areas.
Keywords: gene expression,lignin synthesis,lodging-resistance,hybrid wheat
1.lntroduction
Wheat (Triticum aestivumL.) accounts for about 20% of the total caloric intake of human beings (Chenet al.2020),and stem lodging is a major problem impairing its productivity (Shahet al.2017;Yuet al.2021;Donget al.2023).The yield reductions,which mainly depend on the occurrence period and lodging duration (Shahet al.2017),range from 10 to 57% in China (Luoet al.2019),and the grain quality can also be reduced by lodging (Nakajimaet al.2008).Semi-dwarfing genes,such asRht-B1b,Rht-D1b,Rht8,Rht24b,and others,are widely used in wheat dwarf breeding (Zhanget al.2020;Tianet al.2022;Xionget al.2022).However,further breeding for dwarfing is not conducive to further yield improvements due to the reduced plant biomass (Berryet al.2014).Therefore,appropriately improving plant height (PH) and stem lodging resistance may be an important aspect to consider in future high-yield breeding.
Owing to the outstanding heterosis of yield and stress resistance,hybrid wheat is expected to make great contributions toward meeting the needs of the growing population in the future (Longinet al.2012;Niet al.2017).The technical system of two-line hybrid wheat in China was first established in 2011 by our team,which offered prospects for its application on a commercial scale,and it has entered a relatively rapid development stage.To ensure higher biomass accumulation,the PH of the partial hybrid wheat currently approved is higher than that of conventional varieties,which may in turn increase the risk of lodging.Therefore,improving the lodging resistance ability is more important for hybrid wheat application in the future.
In addition to the stem-related traits,various agronomic measures are known to be closely related with lodging (Berryet al.2004;Penget al.2014;Shahet al.2017;Luoet al.2019).The stem-related traits such as the center of gravity height (CGH),stem diameter,and stem wall thickness contribute in determining the lodging susceptibility (Islamet al.2007;Wanget al.2013).Our studies showed that the breaking strength of the basal second internode (BSBSI) is closely related to the lodging of hybrid wheat (Yanget al.2022),which was consistent with studies on conventional varieties (Wanget al.2013;Penget al.2014).Therefore,the basal second internode (BSI) could be used as an important research marker for studying the lodging of hybrid wheat.A close correlation between lignin content and the rigid breaking strength of crop stems has been found (Wanget al.2015;Huet al.2017),and studies have shown that a lower lignin content can cause lodging (Wanget al.2014).The lignin content is largely affected by species and environmental factors (Luoet al.2019).Specifically,lignin may affect the strength and rigidity of the basal stem by affecting the cell wall stiffness (Chabanneset al.2001;Konget al.2013;Zhenget al.2017).Lignin is composed of three subunits (guaiacyl,syringyl andp-hydroxyphenyl) that are synthesized from the phenylpropanoid pathway (Yuet al.2021).The biosynthetic pathway has been clearly established and the different genes involved in the intermediate steps,such as phenylalanine ammonialyase (PAL),4-coumaric acid: CoA ligase (4CL),Caffeic acid 3-O-methyltransferase (COMT),cinnamyl alcohol dehydrogenase (CAD) and others,have been thoroughly studied (Ma 2009,2010;Penget al.2014;Nguyenet al.2016;Luoet al.2019).The lodging-resistance combination (LRC) showed higher expression levels of genes related to the lignin biosynthetic pathways when compared with the lodging-sensitive combination (LSC) (Yuet al.2021).So far,few studies have explored the relationships of lignin levels with the mechanical strength of hybrid wheat during the grain-filling stage,and studies on the molecular mechanism of lignin biosynthesis between LRC and LSC are also scarce.
In this study,we selected two hybrid combinations with differences in lodging resistance to analyze the variations in stem-related traits and lignin content,and then the physiological mechanism of the differences in lodging resistance between the two combinations was studied by transcriptome and metabolome analyses.Finally,a comparative analysis of internode lengths between conventional and hybrid varieties was carried out to provide a strong reference for the LRC breeding of hybrid wheat.
2.Materials and methods
2.1.Plant materials,experimental design and sampling
The experiments were carried out at the Experimental Station of Beijing Hybrid Wheat Research Institute in Haidian,Beijing,China (39°56´N,116°17´E) during two growing seasons from 2017 to 2018 and 2018 to 2019.According to previous results (Yanget al.2022),two hybrid combinations with differences in lodging resistance (i.e.,the lodging-resistant and lodging-sensitive combinations were BS1086×14GF7433-1 and BS366×14GF7453,respectively) were planted in a plot in a completely randomized block design with three replicates.A total of six plots were planted for the two combinations,and the plot size was 3 m×1 m with 0.25 m between rows.The materials were grown at a density of 180 plants m-2.In the growing seasons,120 kg N ha-1,60 kg P2O5ha-1,and 100 kg K2O ha-1were applied as basal fertilizer before planting,with 120 kg N ha-1applied as top-dressed at the jointing stage (GS31) (Zadokset al.1974).To ensure the consistency of the developmental process of the hybrid wheat,about 60 main stems whose spikes flowered on the same day were chosen and tagged in each plot.A total of 20 main stems in each plot were first taken for morphological agronomic investigation at 15 and 28 days post-anthesis (DPA),and then the BSI of the main stems were used for determining the contents of lignin and the different monomers.The 10 BSI of the tagged main stems in each plot at 15 and 28 DPA were immediately frozen in liquid nitrogen and stored at -80°C for transcriptome and metabolite analyses.The four sample types were: HL,high lodging-resistance combination at the later grain-filling stage;HM,high lodging-resistance combination at the mid grain-filling stage;SL,lodging-sensitive combination at the later grain-filling stage;and SM,lodging-sensitive combination at the mid grain-filling stage.
In addition,we also carried out a comparative experiment on lodging-related traits from 2019 to 2020 at the experimental station of hybrid wheat in Nanyang (34°40´N,112°21´E),Henan Province,China.A total of six conventional wheat varieties (Zhoumai 18,Jimai 22,Shannong 24,Jimai 211,Zhengmai 9023,and Zhongmai 30)with large planting areas in the Huang-Huai Rivers Valley Winter Wheat Zone and three hybrid wheat varieties (Jingmai 179,Jingmai 11,and Jingmai 183) were selected.Each material was planted in a plot of 1.5 m×2 m with 0.25 m between rows.Related data for about 15 main stems for each material were investigated at maturity.
2.2.Determination of morphological characteristics and lignin content
Samples were analyzed according to previously described methods (Zhenget al.2017;Chenet al.2018;Zhang Pet al.2022).A total of 20 representative main stems in each plot at 15 and 28 DPA were chosen for measurements of PH,CGH,the length of the basal second internode (LBSI),the fresh weight of a single stem (FWSS),the fresh weight from the bottom of BSI to the spike top (FWBS) and the stem pole substantial degree (SPSD).The diameter of the basal second internode (DBSI) and wall thickness of the basal second internodes (WTBSI) were measured at the internode mid-region using a digital calliper with an accuracy of 0.001 mm.BSBSI and rind penetration strength (RPS) were measured at 15 and 28 DPA following previous methods (Chenet al.2018;Zhanget al.2020).
The BSI of the 20 representative main stems at 15 and 28 DPA were used for lignin determination,and the lignin was measured by the concentrated sulfuric acid method accoding a previous report (Xionget al.2005).Briefly,0.1 g of crushed samples were placed in test tubes with 10 mL of 1% acetic acid solution.The precipitate was washed with 5 mL of 1% acetic acid,then soaked three times in a mixture of 3-4 mL of ethanol and ether,each time for 3 min.The precipitate was evaporated in a centrifuge tube to dryness,then 3 mL of 72% sulfuric acid was added to dissolve all the cellulose.A total of 10 mL distilled water was added to the test tube which was placed in a boiling water bath for 5 min.A total of 5 mL distilled water and 0.5 mL of 10% barium chloride solution were added after cooling.Subsequently,10 mL of 10% sulfuric acid and 10 mL of 0.1 mol L-1potassium dichromate solution were added to the lignin precipitate.The test tube was placed in boiling water for 15 min,and then cooled for later analysis.The sodium thiosulfate titration method was used to determine the lignin content,and the lignin was expressed as a percentage of dry matter.
The sodium hydroxide solution method to hydrolyze lignin monomers was used to determine the lignin monomer contents (i.e.,the S,G,and H subunits) (Mokochinskiet al.2015).Qualitative and quantitative analyses of the three lignin monomers were performed by the ultra-high performance liquid chromatography-mass spectrometry (UHPLC-MS,AB SCIEX,USA) method.About 0.1 g of powder was placed into a test tube,2 mL of 4 mol L-1sodium hydroxide solution was added,the tube was sealed and the sample was hydrolyzed at 95°C for 24 h.Then,1.6 mL of 6 mol L-1hydrochloric acid was added and the sample was thoroughly shaken to ensure complete neutralization.The reaction mixture was then centrifuged at 13,000 r min-1for 5 min,and 500 μL of the supernatant was transferred to a new centrifuge tube and extracted twice with 1 mL ethyl acetate.The organic phases were combined and dried with nitrogen gas.The samples obtained were resuspended and filtered using a 0.22 μm filter membrane,and finally a 2 μL sample was injected for HPLC-MS analysis.
2.3.Sample preparation,and metabolite extraction and quantification
The sample preparation,extract analysis,and metabolite identification and quantification were performed as previously described (Chenet al.2013) at Wuhan Metware Biotechnology Co.,Ltd.,Wuhan,China.The identified metabolites were annotated using the Kyoto Encyclopedia of Genes and Genomes (KEGG) compound database (http://www.kegg.jp/kegg/compound/),and annotated metabolites were then mapped to the KEGG pathway database (http://www.kegg.jp/kegg/pathway.html).Metabolites with VIP (variable importance in projection)≥1 and |log2FC (fold change)|≥1 were considered to be differential metabolites.
2.4.RNA extraction,cDNA library construction and transcriptome sequencing
The RNAprep Pure Plant Kit (TIANGEN,Beijing,China) was used to extract the RNA.Sequencing libraries were generated using NEBNext®UltraTMRNA Library Prep Kit for Illumina®(NEB,USA) and index codes were added to attribute the sequences to each sample.Referring to a previous report (Yuet al.2021),the library quality was assessed on the Agilent Bioanalyzer 2100 System.The clustering of the index-coded samples was performed on a cBot Cluster Generation System using TruSeq PE Cluster Kit v3-cBot-HS (Illumia).After cluster generation,the library preparations were sequenced on an Illumina Novaseq platform and 150 bp paired-end reads were generated.
2.5.Transcriptome data processing
Raw data (raw reads) in the fasta format were first processed using in-house perl scripts.In this step,clean data (clean reads) were obtained and mapped to the reference genome.A differential expression analysis was conducted on the various sample groups.The resultingP-values were adjusted using the Benjamini and Hochberg approach for controlling the false discovery rate.Genes with |log2FC|≥0 and an adjustedP-value<0.05 were considered differentially expressed.
2.6.Gene expression analysis
To verify the reliability of the transcriptome sequencing data,some genes related to the phenylpropanoid metabolic pathway were subjected to qRT-PCR in three biological replicates.The primers (Appendix A) used in the qRTPCR analysis were designed using Primer 5.0.The 2-ΔΔCtmethod was used to calculate the relative gene expression levels (Livak and Schmittgen 2001).The geneActinwas used as an endogenous control for normalization.
2.7.Statistical analysis
The multivariate analysis of variance was performed using Data Processing System (DPS) software (version 7.05,China),and significant differences were calculated by the least significant difference (LSD) test atP<0.05.The differential genes and metabolites in the same group were mapped to the KEGG pathway,and Pearson correlation analyses were performed to explore the correlations between phenylpropanoid biosynthesis related gene expression levels and lignin biosynthesis precursors.Those with a Pearson correlation coefficient |r|>0.9 and aP-value<0.05 were regarded as statistically significant.
3.Results
3.1.Analysis of stem characteristics
The index weight of BSBSI (Appendix B) indicated that BSBSI should be taken as the research perspective.BSBSI,RPS and SPSD varied with hybrid combinations and sampling times,and their values decreased during the grain-filling process (Table 1).The BSBSI,RPS,WTBSI,FWSS,FWBS and SPSD of LRC were significantly higher than those of LSC,while there were no significant differences in PH,CGHor LBSI between the two cultivars.According to this study,the years and developmental stages interacted significantly with BSBSI,and the combinations and development stages interacted significantly in RPS.BSBSI,RPS and SPSD exhibited interannual stability in the different years,indicating that they could be used as important indicators in lodgingresistance breeding.

Table 1 Main characteristics of the stem-related traits in two hybrid combinations with differences in lodging resistance1)
3.2.Lignin and monomer contents
The results showed that the DBSI of HL was higher than that of SL,and WTBSI showed the same trend (Fig.1-A),which was consistent with the data in Table 1.The lignin content increased during the filling process,and the total lignin content of HL and HM were higher than that of SL and SM,respectivily.The G monomers showed the largest proportion,followed by the S monomers,whereas the H monomers showed the lowest proportion in the same hybrid combination.The ratio of G+S lignin monomers in HL (HM) was higher than that of SL (SM),but the H monomers showed no significant difference between HL and SL (Fig.1-B).

Fig.1 Differences in phenotype and lignin content of the basal second internode.A,the appearance and cross-sections of the basal second internodes of high lodging-resistance (HL) and lodging-sensitive (SL) combination at the later grain-filling stage,respectively.Bar=1 cm.B,the changes in lignin and monomer contents.SM and HM,lodging-sensitive and high lodging-resistance combination at the mid grain-filling stage,respectively.Bars mean SE (n=3).Different letters indicate significant differences (P<0.05) among the different cultivars.
3.3.Analysis of widely targeted metabolomic and metabolic differences
The cluster heat map showed that all three biological replicates were clustered together (Appendix C).A PCA was conducted for the metabolomic data of the 12 analyzed samples to evaluate the overall metabolic differences between the samples in each group and the degree of variability.The repeatability within a group met the requirements,indicating that substance accumulations at the metabolomic level responded well to varieties and sample times (Appendix C).
A total of 432 differentially expressed secondary metabolites were identified.The numbers of differentially expressed metabolites (DEMs) that were downregulated and upregulated in HMvs.SM,HMvs.HL,HLvs.SL and SMvs.SL were 9 and 32,75 and 38,60 and 41,and 129 and 48,respectively (Fig.2-A).Within the KEGG categories,the terms related to “Phenylpropanoid biosynthesis” and “Phenylalanine metabolism” were significantly enriched in HLvs.SL (P<0.05) (Fig.2-D).In this experiment,a total of nine substances in the lignin biosynthesis pathway were identified (Fig.2-F).Three lignin related metabolites,coniferaldehyde,ferulic acid and coniferyl alcohol,were obviously accumulated in SL as compared to HL.Most of the metabolites related to lignin synthesis,particularly ferulic acid and L-phenylalanine,were higher in SL than in SM,whereas the metabolites in HL show a slight reduction when compared with HM (Appendix D).

Fig.2 Identification and functional characterization of the differentially expressed metabolites (DEMs) in hybrid combinations.A,the number of DEMs.B-E,Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of the DEMs in the comparisons of HM vs.SM,HM vs.HL,HL vs.SL and SM vs.SL.F,accumulation patterns of the secondary metabolites involved in lignin biosynthesis pathways.The rows in the heatmaps represent metabolites,and the columns indicate samples.The colors of heatmap cells indicate the accumulation levels of metabolites across different samples.HM and SM,high lodging-resistance and lodging-sensitive combination at the mid grain-filling stage,respectively;HL and SL,high lodging-resistance and lodging-sensitive combination at the later grain-filling stage,respectively.
3.4.Overview of the transcriptome sequencing and gene expression analysis
The RNA-seq results showed that about 143.3 Gb of clean data were obtained from the 12 samples,and the Q30 percentage was >94% (Appendix E).When the clean data were compared to those of the reference genome,the rate of correspondence was in the range of 75.25-95.05% (Appendix F).Moreover,the Pearson’s correlation coefficient (r) and PCA results demonstrated suitable biological reproducibility (Appendix G).Gene expression levels are expressed as fragments per kilobase of transcript sequence per millions base pairs sequenced (FPKM) (Appendix G).These results suggest an effective sampling procedure and a good-quality bioinformatics analysis of the RNA-seq results.
We further investigated the gene expression profiles by analyzing the differentially expressed genes (DEGs) of the different varieties and developmental stages.The total number of DEGs per group is shown in Fig.3-AF.There are 22,196 DEGs in HMvs.HL,of which,11,288 are upregulated and 10,908 are downregulated.A total of 17,300 DEGs exist in SMvs.SL,including 7,794 upregulated and 9,506 downregulated.A total of 7,673 DEGs exist in HMvs.SM,including 3,972 upregulated and 3,701 downregulated.There are also 7,300 DEGs in HLvs.SL,including 3,665 upregulated and 3,635 downregulated.Moreover,the KEGG analysis was carried out to identify the metabolic pathways of the DEGs.The DEGs involved in lignin biosynthesis pathways (phenylalanine metabolism,phenylpropanoid biosynthesis) were significantly enriched in HLvs.SL,while their enrichment in HMvs.SM was not significant.
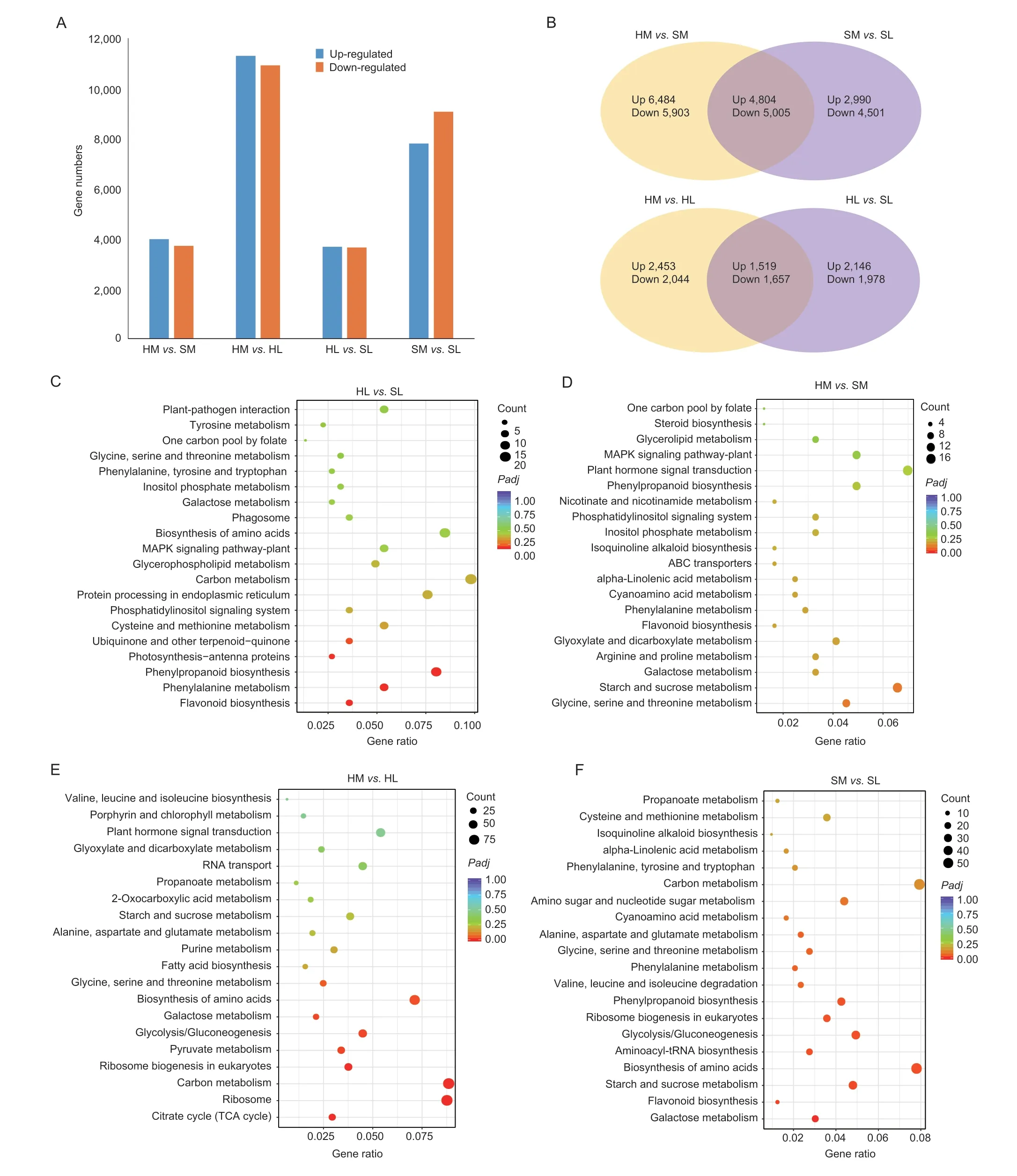
Fig.3 Identification and functional characterization of the differentially expressed genes (DEGs) in hybrid combinations.A,the number of DEGs.B,Venn diagrams showing DEGs between samples from different lodging-resistant hybrid wheat at two sampling times.C-F,Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of the DEGs in the comparisons of HL vs.SL,HM vs.SM,HM vs.HL,and SM vs.SL.HM and SM,high lodging-resistance and lodging-sensitive combination at the mid grain-filling stage,respectively;HL and SL,high lodging-resistance and lodging-sensitive combination at the later grainfilling stage,respectively.
3.5.The key lignin biosynthesis pathway related genes
According to the KEGG results,35 critical regulatory genes involved in the phenylpropanoid pathway (Fig.4-A),including fourPAL,threeC4H,twoCMOT,two4CL,fourCCR,threeCADand 17PER,were identified.Only 10% of the identified genes showed significant differential expression in HMvs.SM,while the genes with significant differential expression accounted for 42% in HLvs.SL,and more than 80% of the DEGs in HL were strongly upregulated when compared with SL.The expression levels of genes involved in the phenylpropanoid pathway in the same combination varied with the grain-filling stage.For example,among the 17 peroxidase coding genes,some are highly expressed at the mid grain-filling stage,while others are highly expressed at the later filling stage.Further,the expression patterns of five genes were also confirmed by qRT-PCR (Fig.4-B).Based on the results of a correlation analysis (Fig.4-C;Appendix H),a total of 19 vital regulatory genes were highly associated with substances (coniferyl alcohol,p-coumaraldehyde,L-phenylalanine,caffeic acid sinapinaldehyde,coniferaldehyde,and ferulic acid) and lignin monomers (G and S monomers).Notably,TraesCS3D02G189900,TraesCS5D02G168400andTraesCS7D02G347300displayed high correlations with the changes in essential lignin precursors (r>0.98,P-value<0.01).Moreover,TraesCS2D02G150400andTraesCS7B02G052600were positively and significantly correlated with the S and G monomers (r>0.98,P-value<0.01),respectively.
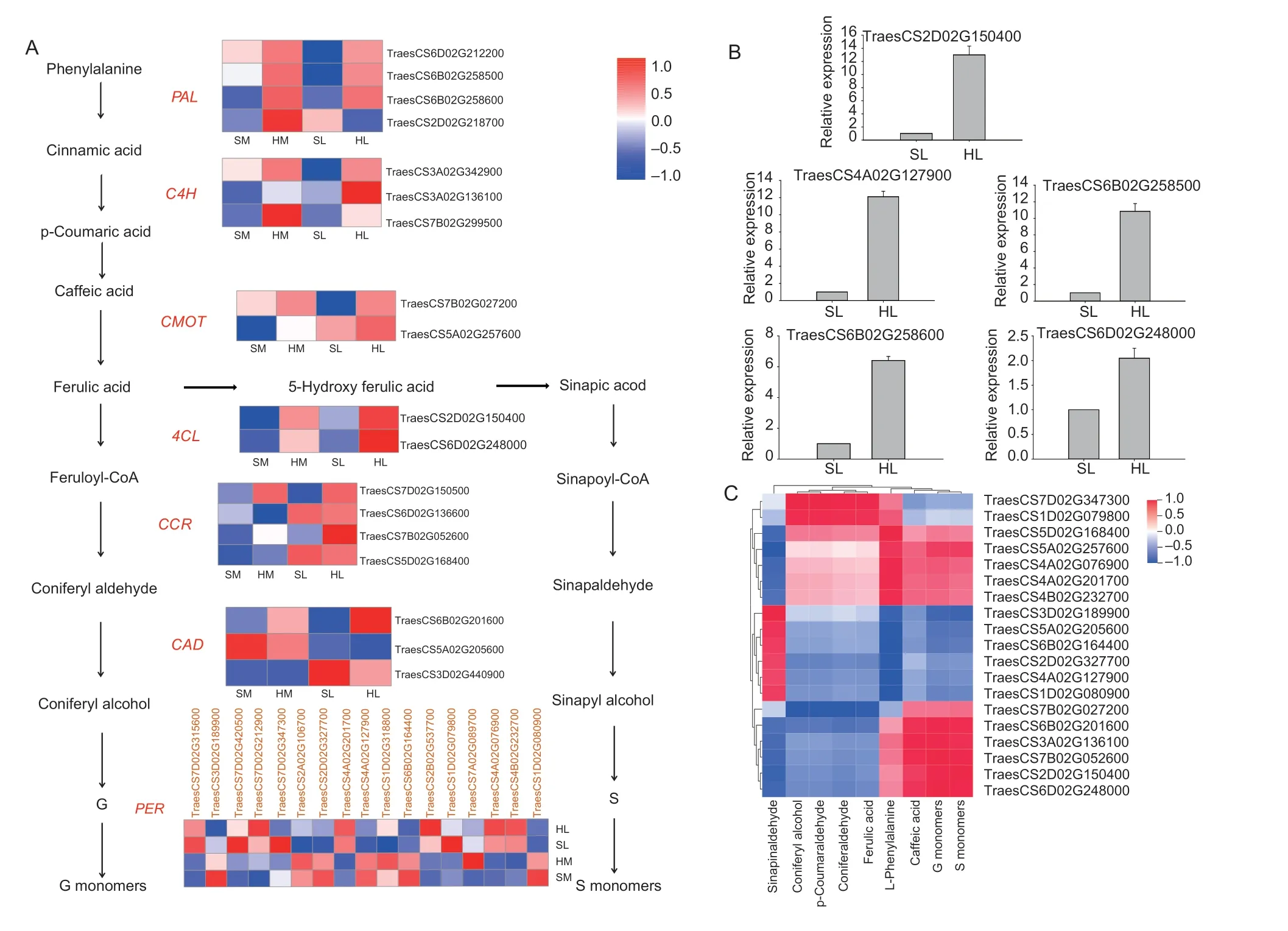
Fig.4 Expression levels of differentially expressed genes (DEGs) involved in the lignin biosynthesis pathway and its relations to intermediate metabolites.A,expression patterns of key lignin biosynthesis genes of different lodging-resistant hybrid combinations.The colors of heatmap cells indicate expression levels of genes across different samples.HM and SM,high lodging-resistance and lodging-sensitive combination at the mid grain-filling stage,respectively;HL and SL,high lodging-resistance and lodgingsensitive combination at the later grain-filling stage,respectively.B,qRT-PCR was used to verify the expression profiles of selected lignin biosynthesis genes.Bars mean SE (n=3).C,heatmap of the correlation coefficients between intermediate metabolites and expression levels of genes involved in lignin biosynthesis,with the red and blue blocks representing positive and negative correlations,respectively.
3.6.Comparative analysis of PH characters between hybrid wheat and conventional wheat
The production practice shows that the PH of applied hybrid wheat is higher than that of conventional wheat (Fig.5-B).In this experiment,a total of six conventional varieties with a large amount of application in the main wheat producing areas of China and three hybrid wheat varieties were used as materials to compare and analyze the variations in internode length.The results showed that compared with conventional wheat,the length of each internode and the spike of hybrid wheat showed an increasing trend,in which the BSI increased the most,with an average increase of 3.3 cm (Fig.5-A).Finally,considering with the above data (Table 1),the breeding strategies to improve lodging resistance of hybrid wheat can be put forward.Specifically,the results suggest that improving the DBSI,WTBSI and lignin synthesis ability is the key for improving the lodging resistance of hybrid wheat (Fig.5-C).

Fig.5 Comparative analysis of plant height characters between hybrid wheat and conventional wheat.A and B,analysis of length variations and plant height differences between conventional wheat and hybrid wheat.C,ways to improve lodging resistance of hybrid wheat.BSBSI (N),breaking strength of the basal second internode;RPS (N),rind penetration strength;SPSD (mg cm-1),stem pole substantial degree;DBSI (mm),diameter of the basal second internode;WTBSI (mm),wall thickness of the basal second internode.
4.Discussion
4.1.Relations of stem-related traits to lodging in hybrid wheat
The genetic improvement of stem-related traits provides one promising strategy for lodging-resistance breeding (Haiet al.2005;Huanget al.2006;Zhanget al.2020).The dwarfing genes such asRht1,Rht2andRht8had larger phenotypic effects on reducing PH,CGH and the length of BSI (Van De Veldeet al.2021;Chaiet al.2022).However,further dwarfing breeding led to reductions in photosynthetic capacity and plants biomass,which is not conducive to further yield improvement (Berryet al.2014),and the PH should be appropriately increased to improve the yield in modern wheat breeding programs (Zhanget al.2007;Wanget al.2014;Zhang Xet al.2022).At present,the PH of partial hybrid varieties is significantly higher than that of conventional varieties,and the increase in the BSI is the largest,which is consistent with the suggestion that the length of BSI was significantly and positively correlated lodging resistance (Zhanget al.2020).In this study,the BSBSI,DBSI and WTBSI of HL were significantly higher than those of SL (Fig.1-A),so they should be the focus for reducing the lodging risk caused by a greater PH in lodging-resistance breeding.Our results also showed that RPS and SPSD were closely related to the stem strength,indicating that they can be used as important indicators for lodgingresistance breeding of hybrid wheat,so this study proposes a phenotypic selection strategy to improve lodging resistance for hybrid breeding to some extent (Fig.5-C).Our previous results also indicated that the lodging resistance of hybrids was easily influenced by the male sterile line (Yanget al.2022).Therefore,the genetic improvement of the previously mentioned stem traits in male sterile line breeding is very important,and the marker-assisted selection related to stem strength (BSBSI,RPS,DBSI,WTBSI,etc.) may be used to improve the lodging resistance of male sterile lines in the future (Haiet al.2005;Huanget al.2006;Ookawaet al.2016).
4.2.Relationship of lignin synthesis and lodging
The stem strength,as a major trait for the improvement of lodging-resistance (Zhanget al.2020),was significantly correlated with lignin content and stem diameter (Zhenget al.2017).Due to the significant reduction of grain yield induced by lodging during the grain-filling stage (Penget al.2014;Luoet al.2019),the BSI values at the mid and later filling stages were used for studying the lodging-resistance mechanism of hybrid wheat.Our data showed that the BSI of HL had higher contents of lignin and the G and S monomers when compared to SL,which may increase the SPSD of HL,and thereby significantly increase the BSBSI and RPS.Due to the close relationship of lignin levels with lodging resistance (Penget al.2014;Liet al.2021;Yinet al.2022),we speculate that the higher lignin content in the later grain-filling stage may be the key to the higher lodging resistance of HL.Although the lignin biosynthetic pathway has been clearly established (Luoet al.2019;Yaoet al.2021),few studies have focused on the lignin biosynthesis of hybrid wheat during grain-filling.In this study,the transcriptome and metabolome data of the BSI at the mid and later grainfilling stages were analyzed to gain insights into the molecular basis for lignin biosynthesis.The metabolome analysis showed that the phenylpropanoid biosynthesis pathways were significantly enriched in HLvs.SL,which strongly proves its important role in lodging-resistance formation at the later grain-filling stage.The metabolome data of barley at the booting stage showed that the contents of lignin biosynthesis related intermediate metabolites in LSC were significantly lower than in LRC (Yuet al.2021).Interestingly,our study found that almost all lignin biosynthesis-related intermediate metabolites,particularly coniferaldehyde,ferulic acid and coniferyl alcohol,were obviously accumulated in SL as compared to HL,which may be related to the weakened synthesis from intermediate metabolites to lignin monomers.Our study also found that the contents of most of the intermediate metabolites in SL were higher than those in SM,while they showed an oppsite trend between HL and HM,indicating that LRC still has a strong lignin synthesis ability in the later grain-filling stage when compared to LSC.Studies have revealed the close relations between lignin synthesis related genes,such as4CL,PAL,andCAD,and lignin content (Nguyenet al.2016;Huet al.2017).The lignin content was reduced by 25% compared to the wild type when fourPALgenes inArabidopsis thalianawere knocked out (Huanget al.2010).Transcriptomic data showed that the phenylpropanoid biosynthesis pathway was also significantly enriched in HLvs.SL.From the results of our transcriptome and metabolome analysis,we concluded that the phenylpropanoid biosynthesis pathways is the key to higher lignin accumulation and lodging resistance at the later grain-filling stage of LRC.Therefore,it is important to identify the genes involved in the lignin biosynthesis in hybrid wheat.We identified a total of 35 DEGs participating in the lignin biosynthesis pathways,and the numbers of genes were different from those in barley (Yuet al.2021) and bamboo shoots (Houet al.2022).Only 10% of the identified genes showed significant differential expression between HM and SM,while the value accounted for as high as 42% in HLvs.SL.Among them,more than 80% of the DEGs in HL were strongly up-regulated when compared with SL,indicating that LRC has a higher lignin synthesis ability at the later grain-filling stage.Notably,the gene expression trend was almost the opposite of the metabolite contents between HL and SL,which is inconsistent with the results in barley (Yuet al.2021).We speculate that in the conversion of ferulic acid to coniferyl alcohol,4CL,CCRandCADare crucial genes (Nguyenet al.2016;Houet al.2022),and when their expression levels rise (such asTraesCS2D02G150400,TraesCS6D02G248000,TraesCS6B02G201600,andTraesCS4A02G127900),the flow of ferulic acid into G units increases,which in turn leads to the reduction of intermediate metabolites (ferulic acid,coniferylaldehyde and coniferyl alcohol).The results showed that the expression levels ofCAD(TraesCS6B02G201600),C4H(TraesCS3A02G136100),C C R(Traes CS7B02G052600),and4CL(Traes CS2D02G150400,TraesCS6D02G248000) were closely and positivley associated with the G and S monomers (|r|>0.9,P-value<0.05) (Fig.2-C;Appendix H),suggesting their potentially important roles in regulating lignin biosynthesis,and the higher expression of specific genes participating in the lignin biosynthesis pathways of LRC could improve its lignin accumulation and subsequently enhance its lodging resistance.The candidate gene resources identified in this study could be further functionally characterized in order to pinpoint the major genes that can be exploited in molecular hybrid breeding programmes.According to previous reports (Shahet al.2019;Yanget al.2022),the breeding strategy for improving the lodging resistance of hybrid wheat by improving the lignin synthesis ability of BSI was put forward (Fig.5-C).
5.Conclusion
In this study,we found that the BSI of LRC accumulated more lignin and showed higher BSBSI,RPS and SPSD as compared to the LSC.The transcriptome and metabolome analysis together revealed that the higher expression levels of genes related to the phenylpropanoid biosynthesis pathways represent the key mechanism leading to the higher lignin accumulation and lodging resistance at the later grain-filling stage of LRC.Ultimately,a selection strategy based on phenotypic traits (DBSI,WTBSI,BSBSI,etc.) and lignin synthesis ability was put forward for the lodging-resistance breeding of hybrid.
Acknowledgements
This work was supported by the Youth Fund Project from Beijing Academy of Agricultural and Forestry Sciences,China (QNJJ202225),the Germplasm Innovation and New Variety Breeding Project of Beijing,China (G20220628002) and the Training Programme Foundation for the Beijing Municipal Excellent Talents,China (2017000020060G130).
Declaration of competing interest
The authors declare that they have no conflict of interest.
Appendicesassociated with this paper are available on https://doi.org/10.1016/j.jia.2023.06.027
杂志排行

Journal of Integrative Agriculture的其它文章
- Invasion of fall armyworm led to the succession of maize pests in Southwest China
- Genome-wide and candidate gene association studies identify BnPAP17 as conferring the utilization of organic phosphorus in oilseed rape
- Impacts of agri-food e-commerce on traditional wholesale industry: Evidence from China
- Responses of growth performance,antioxidant function,small intestinal morphology and mRNA expression of jejunal tight junction protein to dietary iron in yellow-feathered broilers
- Genome-wide identification of the CONSTANS-LIKE (COL) family and mechanism of fruit senescence regulation by PpCOL8 in sand pear (Pyrus pyrifolia)
- ldentification of S-RNase genotype and analysis of its origin and evolutionary patterns in Malus plants