毛竹向杉木林扩张过程中叶功能性状季节变化1)
2024-05-11张淑洁顾娇周秀梅王楠白尚斌
张淑洁 顾娇 周秀梅 王楠 白尚斌
(浙江农林大学,杭州,311300) (浙江农林大学暨阳学院)
植物入侵严重威胁生物多样性,破坏生态系统的功能和服务,是当前国际公认的环境问题[1]。当外来入侵物种进入本地生态系统时,它们会受到来自本地物种的竞争压力,这种竞争可以通过比较外来种类和当地种类在某些特征上的不同来发现[2]。已有研究发现,植物的入侵受其自身特征(如生长、传播、繁殖等)的影响,这些特征是植物入侵成功与否的关键[3]。对于某些入侵性较强的植物而言,其生长速率和繁殖能力普遍高于普通植物[4]。植物的相对生长速率与叶功能性状有着直接的关系,如叶干物质质量分数、比叶面积及N、P、K等元素质量分数与植物生长速率呈正相关。叶营养元素(如N)作为光合作用的必需物质,与光合速率呈显著正相关[5-6]。较高质量分数叶元素与利用效率有利于提高植物光合速率[7-8]。较高的比叶面积、N质量分数和光合速率皆对植物的相对生长速率有提高作用,促进植物入侵[9]。
毛竹(Phyllostachysedulis)与众多木本植物相比,特性显著,开花时间长、茎秆寿命短,拥有较强的无性繁殖能力。此外,毛竹具备极强的繁衍潜力及营养体分化特性,其地下茎繁殖分生能力极强,主要功能为养分贮藏和运输,一片竹林就像是一棵巨大的竹树[10]。毛竹属于我国的本地种,其强大的扩鞭能力,使其具有类似入侵种的特征,常常扩张进入周边常绿阔、针叶树林。
有相关研究表明,毛竹在扩张过程中,其根系功能性状发生了一定的变化[11-12],毛竹向撂荒地扩张中表现出了叶性状的改变[13],但关于毛竹向周围针叶林扩张的过程中,其叶功能性状的季节性变化研究相对较少。毛竹向杉木(Cunninghamialanceolata)林扩张时,是否会改变其叶功能性状,以及是否存在季节性差异来实现或加速其扩张进程尚不清楚。本研究以毛竹-杉木混交林为研究对象,以毛竹纯林为对照,对比分析不同季节,毛竹扩张杉木林的过程中其叶片功能性状的变化,以阐明毛竹扩张时毛竹叶片功能性状的季节变化特征,旨在揭示毛竹在扩张过程中采取的叶功能对策,有利于其扩张进程,从而为更好地理解毛竹扩张机制和合理地经营调控毛竹扩张提供科学参考。
1 研究区概况
研究区域位于浙江省杭州市临安区於潜镇南山村实验林场(119°13′12″E,30°2′6″N),地处浙江西北部,亚热带季风气候,光照充足,温暖湿润,雨量充沛,四季分明。年均降水量1 613.9 mm,年均降水日158 d,年平均无霜期为237 d。低山丘陵地貌,海拔100~200 m,土壤为黄壤。植被主要以杉木次生林和毛竹林为主,大部分毛竹林是由人工种植后自然扩张形成的。故而,在研究区内存在毛竹纯林和毛竹-杉木混交林两种林分。毛竹林中的林下优势植被主要有盾叶莓(Rubuspeltatus)、茅莓(Rubusparvifolius),其他植被还有日本野桐(Mallotusnepalensis)、渐尖毛蕨(Cyclosorusacuminatus)、三脉紫菀(Asterageratoides)等。毛竹与杉木为乔木层,紫珠(Callicarpabodinieri)和日本野桐等为灌木层,金线草(Antenoronfiliforme)及姬蕨(Hypolepispunctata)等为草本层。
2 试验方法
试验设计:研究区内毛竹林是人工种植经营后抛荒形成。皆伐(2008年)后的杉木林通过自然更新形成次生林。毛竹林随即向杉木次生林扩张,形成竹杉混交林。选取立地条件基本相似的区域布置2种林分样地,分别为毛竹-杉木混交林(MF)和毛竹纯林(BF)。样地内无人为干扰,为自然生长竞争状态。每种林分设置10 m×20 m的样方各3个,且每个样地中心外围均设置大于10 m的缓冲区。于生长季(7月)对样地内的树木进行树高、胸径、密度等样地调查,基本情况如表1。

表1 样地基本概况
叶片采集法:于2021年1月(冬季)、4月(春季)、7月(夏季)、10月(秋季),在连续晴朗3~5 d后的上午,于两种林分6个样地中各随机选取平均胸径和树高接近的毛竹样株5株,每种林分共15株,两种林分共计30株,用高枝剪采集位于毛竹树冠4个方向(东南西北)的叶片,每株共采集40片完整叶片。将采集的叶片放入已编号自封袋中,带回实验室,用于叶功能性状的测定。
叶结构性状的测定:将采集的毛竹叶片平展后,用水平仪相机进行拍摄,获得图像后,用Image J软件进行叶长、叶宽和叶面积的计算。在叶片上、中、下3个位置,用精度为0.01 mm的游标卡尺测定叶厚,取平均值。用电子天平(精度0.01 g)称取叶鲜质量,后将叶浸泡于清水中,4 ℃黑暗环境浸泡12 h,取出,吸干表面水分,称得叶饱和鲜质量。再将叶片置于烘箱(70 ℃)中,烘干至恒质量,称得叶干质量。根据已测得指标进行其他指标计算。计算公式如下:
比叶面积=叶面积/叶干质量;
叶干物质质量分数=叶干质量/叶饱和鲜质量;
叶相对含水量=[(叶鲜质量-叶干质量)/(叶饱和鲜质量-叶干质量)]×100%;
叶组织密度=叶干质量/(叶面积×叶厚);
叶形态指数=叶长/叶宽。
叶养分质量分数的测定:毛竹叶片烘干后进行研磨处理,过筛(孔径为0.15 mm),测定养分质量分数。采用重铬酸钾加热法测定叶碳质量分数,凯式定氮法测定叶氮质量分数,钼锑抗比色法测定叶磷质量分数,火焰光度计测定叶钾质量分数[14]。
叶光合色素质量分数的测定:采用混合液浸提-分光光度计法测定。将鲜叶剪碎混匀,称取样品0.2 g,放入研钵,加入碳酸钙、石英砂便于研磨,叶片碾碎后,按体积比4.5∶4.5∶1.0加入丙酮、无水乙醇和水,配成混合提取液,将研钵中绿色液体倒入具塞比色管,定容至25 mL(记为V),标号,置于黑暗环境中24 h,滤纸过滤,将过滤完的液体倒入比色皿中,以丙酮、乙醇和水的混合液为参照,用分光光度计测定645、663 nm波长的光密度(D),根据Arnon公式计算叶绿素质量分数[15]。
叶绿素a质量分数=(12.72D663-2.59D645)×V/(1 000×0.2);
叶绿素b质量分数=(22.88D645-4.67D663)×V/(1 000×0.2);
总叶绿素质量分数=(8.05D663+20.29D645)×V/(1 000×0.2)。
叶片解剖结构的观察:采用石蜡切片法[16]。将固定好的叶片制成透明装片,放入显微镜(NIKON ECLIPSE E100,尼康株式会社,日本)观察,用成像系统(NIKON DS-US,尼康株式会社,日本)进行图像采集分析。
数据处理:用SPSS 22.0(IBM,美国)进行数据统计分析。采用重复测量方差分析比较不同林分间毛竹叶功能性状季节变化差异(显著性水平为P<0.05),分析时间、林分以及两者之间的交互作用影响。采用单因素方差分析比较两林分间毛竹叶解剖结构变化差异(显著性水平为P<0.05)。
3 结果与分析
3.1 毛竹叶结构性状季节变化特征
由表2可知,毛竹-杉木混交林内的毛竹各季节叶厚从大到小依次为春、夏、冬、秋,而毛竹纯林内的毛竹各季节叶厚从大到小依次为夏、秋、春、冬;毛竹-杉木混交林内的毛竹叶厚在春季达到最大值,为0.086 mm,而毛竹纯林在夏季达到最大值,为0.093 mm;在夏秋两季,毛竹-杉木混交林内毛竹叶厚显著低于毛竹纯林(P<0.05)。毛竹-杉木混交林中,毛竹比叶面积夏秋大于春冬,在夏季达到最大值,为255.085 cm2/g;而毛竹纯林中的毛竹比叶面积则为秋冬大于夏春,在秋季达到最大值,为200.569 cm2/g;除冬季外,毛竹-杉木混交林内毛竹叶片比叶面积均大于同季毛竹纯林内的毛竹叶片,且在夏季达到显著水平(P<0.05)。毛竹-杉木混交林内的毛竹叶干物质质量分数随季节变化,从大到小依次为春、冬、秋、夏,而毛竹纯林内毛竹叶干物质质量分数从大到小依次为春、冬、夏、秋,两种林分均在春季达到了最大值;但毛竹纯林毛竹叶干物质质量分数在夏季显著高于毛竹-杉木混交林(P<0.05),而在秋季显著低于毛竹-杉木混交林(P<0.05)。毛竹-杉木混交林中的叶相对含水率季节变化趋势为,夏冬大于春秋,最大值出现在夏季,为92.491%,毛竹纯林则是冬春大于秋夏,最大值出现冬季,为92.209%,两种林分变化趋势不同;而在夏季时,毛竹-杉木混交林内毛竹叶片含水率显著高于毛竹纯林(P<0.05)。毛竹-杉木混交林内,毛竹叶组织密度变化趋势为冬高夏低,从大到小依次为冬、秋、春、夏,毛竹纯林内,毛竹叶组织密度变化趋势为冬高秋低,从大到小依次为冬、春、夏、秋,两者的变化趋势不同;在冬季时,两者间的毛竹叶组织密度呈现显著差异,表现为毛竹-杉木混交林显著低于毛竹纯林(P<0.05)。两种林分中,毛竹叶形态指数变化趋势大致相同,均为秋夏大于冬春;但在夏秋两季,毛竹-杉木混交林内的毛竹叶形态指数显著低于毛竹纯林(P<0.05)。
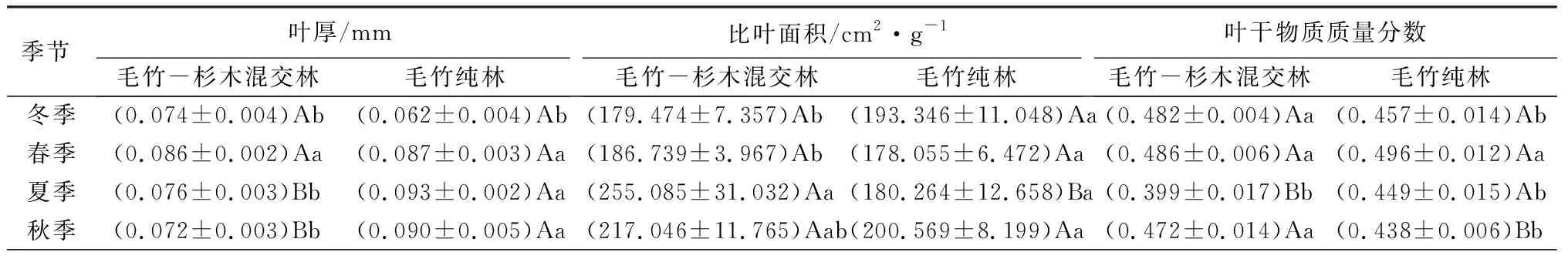
表2 四季中不同林分毛竹的叶结构性状
3.2 毛竹叶养分质量分数季节变化特征
如表3所示,毛竹-杉木混交林内的毛竹叶碳质量分数各季节从大到小依次为秋、春、冬、夏,而毛竹纯林内依次为冬、秋、春、夏;其中,毛竹-杉木混交林在冬季显著低于毛竹纯林(P<0.05);两种林分最低值均出现在夏季,分别为427.327、423.227 g/kg。两种林分内的毛竹叶氮质量分数季节变化趋势一致,从大到小均依次为冬、秋、春、夏。两种林分内的毛竹叶磷质量分数季节变化趋势一致,从大到小均依次为秋、冬、春、夏;而在春季,毛竹-杉木混交林内的叶磷质量分数显著低于毛竹纯林(P<0.05)。毛竹叶钾质量分数季节变化趋势与叶磷一致,均为秋冬高于春夏,但存在一定的差异性;与毛竹纯林相比,毛竹-杉木混交林毛竹叶钾质量分数在春夏两季增加,而秋冬两季减少,并在春季时,毛竹-杉木混交林毛竹叶钾质量分数显著高于毛竹纯林(P<0.05)。两种林分毛竹叶碳、氮质量分数比四季变化一致,从大到小均依次为夏、春、秋、冬;毛竹-杉木混交林和毛竹纯林毛竹叶碳、氮质量分数比夏季为冬季的1.3倍,且差异显著(P<0.05);冬季时,毛竹纯林内毛竹叶碳、氮质量分数比显著高于毛竹-杉木混交林(P<0.05)。两种林分中,毛竹叶氮、磷质量分数比均呈现出夏高冬低的趋势,并在秋季达到最低;与毛竹纯林相比,毛竹-杉木混交林内毛竹叶氮、磷质量分数比在夏秋两季较低,而冬春两季较高,在春季,毛竹-杉木混交林毛竹叶氮、磷质量分数比显著高于毛竹纯林(P<0.05)。
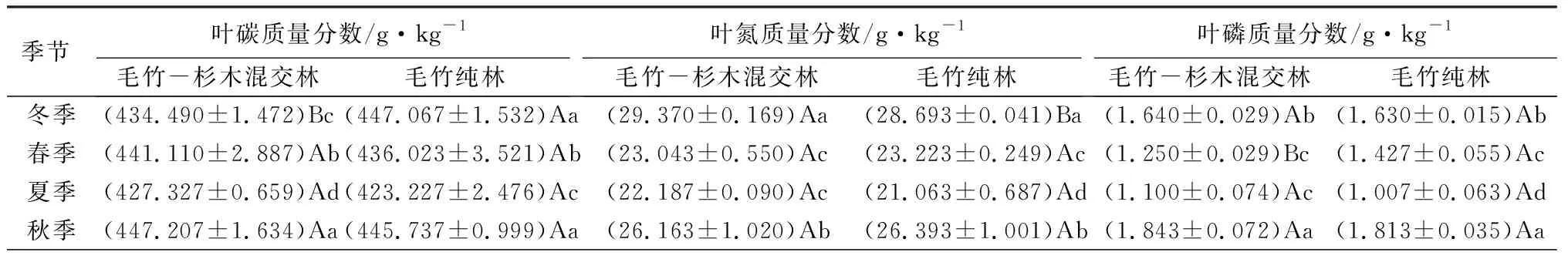
表3 四季中不同林分毛竹的叶养分质量分数
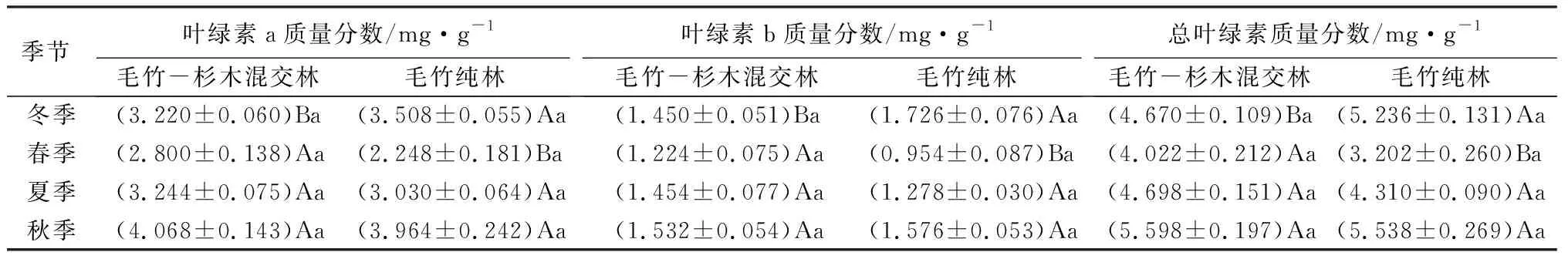
表4 四季中不同林分毛竹光合色素质量分数
3.3 毛竹叶光合色素质量分数季节变化特征
毛竹-杉木混交林内,毛竹叶片的叶绿素a、叶绿素b和总叶绿素质量分数从大到小均依次为秋、夏、冬、春;而毛竹纯林内毛竹叶片内叶绿素a和总叶绿素质量分数的季节变化从大到小依次为秋、冬、夏、春;叶绿素b质量分数各季节从大到小依次为冬、秋、夏、春。两种林分的叶绿素a质量分数都在秋季达到最大值,但毛竹-杉木混交林内为4.068 mg/g,而毛竹纯林内为3.964 mg/g;叶绿素b质量分数都在春季达到最小值,毛竹-杉木混交林内为1.224 mg/g,而毛竹纯林内为0.954 mg/g。毛竹-杉木混交林内总叶绿素质量分数在春夏秋三季均高于毛竹纯林,而在冬季低于毛竹纯林;叶绿素a、叶绿素b和总叶绿素质量分数在冬季均呈现为毛竹-杉木混交林显著低于毛竹纯林(P<0.05),而在春季则相反,显著高于毛竹纯林(P<0.05)。
3.4 不同林分毛竹叶解剖结构变化特征
对毛竹-杉木混交林和毛竹纯林叶片解剖结构进行比较(图1、表5),发现两种林分中毛竹叶片解剖结构类似,但存在一定的差异性,毛竹-杉木混交林内的毛竹叶片韧皮部截面积显著低于毛竹纯林(P<0.05)。与毛竹纯林相比,毛竹-杉木混交林中毛竹叶片角质层厚度、上表皮厚度、下表皮厚度均有所增加,泡状细胞和木质部导管也较为发达。而毛竹纯林中毛竹叶片解剖结构显示,梭形细胞和韧皮部细胞较发达。

A为毛竹-杉木混交林,B为毛竹纯林;UE为上表皮,LE为下表皮,bc为泡状细胞,FVB为细脉维管束,VBS为维管束鞘(两层细胞),fc为梭形细胞,V为木质部导管,Ph为韧皮部,Me为叶肉细胞。图1 不同林分毛竹叶片解剖结构特征(10X)

表5 不同林分毛竹叶片的形态解剖特征
4 讨论
4.1 扩张中毛竹叶功能性状的季节变化特征
植物获取和利用资源的能力体现在植物叶功能性状上,主要表现为,植物适应环境变化的能力及生存对策[17-18],因此常被作为研究植物生态适应性和生存策略的首选指标[19]。叶厚通常用来衡量叶片资源获取、水分保存的能力。毛竹-杉木混交林内,毛竹叶厚在夏秋两季显著低于毛竹纯林(P<0.05),说明毛竹扩张后,毛竹-杉木混交林中的生态环境发生改变,叶片对水分、养分的保存能力变弱,毛竹在扩张过程中也受到了一定的生长阻力。毛竹-杉木混交林和毛竹纯林内,毛竹叶片叶厚不同季节变化趋势说明,毛竹在扩张杉木林后改变其生存策略,在竞争环境中(毛竹-杉木混交林),于生长季加速获取资源,而在独自生长环境中(毛竹纯林),则是选择水热条件最好的夏季进行快速生长。叶片比叶面积一定程度上反映了叶片截获光资源的能力,比叶面积越大,越有利于增强植物捕获资源的能力。刘广路等[13]研究发现,毛竹向撂荒地扩张前期,毛竹叶片比叶面积变大。春、夏、秋三季内,毛竹-杉木混交林叶片比叶面积均大于毛竹纯林,在夏季,毛竹-杉木混交林比叶面积显著高于毛竹纯林(P<0.05),说明毛竹与杉木竞争,提高了自身捕获资源的能力以加强竞争力,加速其自身的发展速度。Leishman et al.[20]研究表明,入侵植物具有较高的比叶面积,能够促进自身生长繁殖。两种林分内的叶片干物质质量分数和组织密度均呈现出与比叶面积相反的趋势,大致呈现为冬春高于夏秋。这是由于两种林分内的毛竹在夏秋两季时,吸收更多的养分用于加速生长,为此,需消耗更多的养分才能增强对资源的竞争力,故导致养分积累能力降低[21]。毛竹纯林中叶干物质质量分数在夏季显著高于毛竹-杉木混交林(P<0.05),而在秋季显著低于毛竹-杉木混交林(P<0.05),说明由于竞争条件的影响,毛竹的生存策略随着竞争力的改变而做出了适应性的调整。但在冬季,两种林分内的毛竹叶片叶组织密度均达到了最大值,并且毛竹-杉木混交林显著低于毛竹纯林(P<0.05),说明两者在冬季有较多的碳储备,而毛竹纯林内碳储备显著高于毛竹-杉木混交林。叶含水量体现植物生存环境的水热条件,较高的叶含水率说明植物对水分的保持能力较强。两种林分内的毛竹叶片含水率变化趋势不同,毛竹纯林内最大值出现在冬季,而毛竹-杉木混交林则是出现在夏季。夏季是试验区内降水量最大的季节,此时,毛竹-杉木混交林叶相对含水率显著大于毛竹纯林(P<0.05),这也表明毛竹在竞争条件时,拥有了更好的水分保持能力。但是,毛竹纯林则在降雨较少的冬季,拥有四季中最好的水分保持力。许凯扬等[22]研究表明,同为入侵植物的喜旱莲子草(Alternantheraphiloxeroides)体现出一定的表型可塑性,具体表现为以不同的形态学特征来适应环境因子变化。两种林分内的毛竹叶形态指数除冬季外,均为毛竹纯林大于毛竹-杉木混交林,并在夏秋两季,达到显著差异水平(P<0.05),这说明在毛竹扩张过程中,其叶片形状特征发生一定的改变。毛竹能够根据所处的环境不同而改变自身的叶片形状,具有表型可塑性。
叶片养分质量分数是衡量植物光合同化能力和生长发育的重要指标。叶片碳元素是构成部分次生代谢产物的重要物质,合成这些物质要耗费较高的能量[23]。当植物将较多能量投入合成这些次生代谢产物,植物生长发育所获得的碳就会减少。许培瑜[24]研究表明,瘤突苍耳(Xanthiumstrumarium)与洋野黍(Panicumdichotomiflorm)等入侵植物,其叶片碳质量分数显著低于本地共生植物。本研究发现,两种林分内的毛竹叶碳质量分数各季节变化趋势不同,毛竹-杉木混交林的毛竹叶碳质量分数在冬季显著低于毛竹纯林(P<0.05),但其余三季,毛竹-杉木混交林中毛竹叶碳质量分数均高于毛竹纯林,说明毛竹在竞争环境,仍旧维持着原本的碳吸收和利用能力,适当增强了自身的抗逆性,能够更好的适应复杂多变的环境。叶片氮元素大部分位于叶绿体中,光合酶的质量分数和活性与其有着直接关系。叶氮质量分数的增加,意味着叶片具有较强的光合能力。两种林分的毛竹叶氮质量分数变化趋势一致,均在冬季达到最大值,且冬季时,毛竹-杉木混交林叶氮质量分数显著大于毛竹纯林(P<0.05)。冬夏两季,毛竹-杉木混交林叶氮质量分数大于毛竹纯林。马可心等[25]相关研究表明,外来入侵植物曼陀罗(Daturastramonium)比本地植物具有更高的氮质量分数,说明在任何环境中,毛竹在冬季都会提高自身资源捕获能力,用于度过自然环境资源相对匮乏的冬季。在竞争环境中,毛竹适当增强了自身的光合能力和碳同化能力,这与毛竹叶组织密度在冬季增加现象相一致。植物叶片碳、氮质量分数比的高低代表植物同化碳及吸收营养元素的能力。植物叶片内碳、氮质量分数比较低时,表明其具有较快生长速率与较强入侵能力[26]。两种林分的毛竹叶碳、氮质量分数比从大到小均呈现为夏、春、秋、冬,但毛竹-杉木混交林在冬季显著低于毛竹纯林(P<0.05),说明在竞争条件中,毛竹在生长季增强其自身的生长速率和入侵能力,从而增加入侵速度。磷在能量代谢中起特殊作用,有提高植物抗逆性和适应外界环境条件的能力。王满莲等[27]研究表明,飞机草(Chromolaenaodorata)与紫茎泽兰(Eupatoriumadenophorum)等入侵植物,叶片氮、磷质量分数显著高于本地植物。本研究中,两种样地内的毛竹叶磷质量分数变化趋势相同,但毛竹-杉木混交林中,毛竹叶磷质量分数在春季显著低于毛竹纯林(P<0.05),说明毛竹在竞争环境中,对磷捕获能力较弱,竞争能力略有降低,但四季中对磷的捕获策略并没有改变。植物叶片的氮、磷质量分数比可作为判断植物生长养分是否受限的重要依据。Koerselman et al.[28]研究表明,植物生长受氮、磷限制的氮、磷质量分数比阈值为14与16,即氮、磷质量分数比在14~16时,受氮、磷共同限制;氮、磷质量分数比<14时,群落中植物生长受氮限制较多;氮、磷质量分数比>16时,植物生长受磷限制较多。两个样地内的毛竹叶片氮、磷质量分数比,秋季时平均值均在14~16,其余三季均大于16,说明样地内的毛竹生长在秋季受到了氮和磷的共同限制,但大多数时间还是受到磷的限制,这也说明了毛竹并没有因为竞争环境的改变而改变养分获取策略。钾主要参与植物体内的新陈代谢,增强植物抗性。两种林分内毛竹叶钾季节变化,在春季和夏季,毛竹-杉木混交林叶钾质量分数大于毛竹纯林,并在春季达到显著差异(P<0.05),说明毛竹竞争过程中适当加强了自身的抗性,能够承受更多的环境压力。闫雅楠等[29]研究表明,加拿大一枝黄花(SolidagocanadensisL.)等入侵植物的叶片钾质量分数高于其伴生植物。
叶绿素是植物吸收、传递、分配和转化光能用于光合作用的色素。光合作用中,叶绿素a参与电子传递和捕捉可见光,叶绿素b具有光能的吸收和传递功能。两种林分内,除毛竹纯林内叶绿素b质量分数呈冬高、春低外,其余毛竹叶片叶绿素a、叶绿素b以及总叶绿素质量分数均呈现秋高、春低的趋势。陈建华等[30]关于毛竹叶片生理特性的研究也发现,毛竹叶绿素在9月份(秋季)达到最大值。同时也说明,毛竹在竞争条件没有改变自身叶绿素季节变化趋势。两种林分毛竹叶绿素的变化反映出,毛竹纯林在冬季光合作用能力大于毛竹-杉木混交林,而毛竹-杉木混交林在春季光合作用能力大于毛竹纯林。
4.2 扩张中毛竹叶组织结构变化特征
植物叶组织结构是植物长期适应环境而表现出的特征,可用其研究植物对环境的特定适应性[31]。毛竹作为禾本科单子叶植物,其解剖结构主要包括表皮、叶脉和叶肉三部分。毛竹-杉木混交林内,毛竹表皮相较毛竹纯林均有一定程度的加厚,这说明毛竹在竞争环境中,一定程度上提高自我保护能力,这能较大程度地减缓蒸腾作用,避免夏季高温灼伤[32]。毛竹-杉木混交林内,毛竹表皮外的角质层也变厚,这起到有效防止蒸腾失水的功能[33]。毛竹-杉木混交林毛竹木质部导管较毛竹纯林更发达,说明其输导能力在竞争条件会适度增强。有研究表明,单子叶植物主要通过增大导管直径提高水分运输能力[34]。泡状细胞具有储藏水分的功能[35],而毛竹-杉木混交林中,毛竹叶片内的泡状细胞较发达,说明毛竹扩张后,其储水能力增强。
5 结论
毛竹扩张进入杉木林后,与杉木形成竞争,除光合色素质量分数外,其叶结构性状和叶养分质量分数在不同季节内均呈现出一定的显著性差异。说明毛竹在向杉木林扩张过程中,表现出一定的表型可塑性,改变了自身的生存策略,以达到生长扩张的目的。同时,毛竹加厚叶片表皮、表皮外角质层,木质部导管和泡状细胞变得较为发达等,叶组织结构也发生了改变。这些结果意味着毛竹扩张后形成的杉竹混交林中,毛竹改变了叶功能性状以在竞争中获取资源。
