植物种子萌发行为和子叶功能的资源利用和能量输出的权衡研究
2024-05-10葛加荣栾艳新季晓静葛雨祺栾志慧
葛加荣,栾艳新,沙 悦,王 媛,丁 鹏,潘 红,季晓静,葛雨祺,栾志慧
种子萌发是高等植物正常生长发育的重要起点,种子能否顺利发芽,对植物种群的扩散、占领新的分布区并通过有性繁殖更新种群及延续物种均起着重要作用[1].植物种子萌发是一个快速和精密的过程,种子萌发调控机制与植物生长有着密不可分的关系.对种子萌发调控机制进行深入研究,不仅可以更加深入地了解植物的适应性和生存策略,也可以为植物种子加工和改良奠定基础,并为生物学多方位的应用研究提供素材.
植物在种子至幼苗生活史阶段存在着两大权衡:一个是种子繁殖和子叶生长之间的权衡,主要表现为植物对自身繁殖的成本.另一个则是种子萌发和子叶功能的资源利用和能量输出的权衡,表现为幼苗建植的适合度[2].一株植物从种子萌发到子叶幼苗建成,经历的一系列生命活动中,由于资源存在定量的限制性,因此准确合理地衡量,以及最佳地配置此阶段的资源尤为重要[3-4].
环境影响着种子萌发及幼苗出土,而种子埋深又是影响种子萌发和幼苗出土的诸多环境因素中的主要因素之一,埋深对种子的大小、发芽率、幼苗出土、定居等都有较强的选择压力,合适的埋深,可以为种子发芽和幼苗的出土提供一定的保护[5].
松嫩草地的优势植物多为一年生或多年生单子叶和双子叶草本植物,随着全球温度的升高,松嫩草地的生境破坏、盐碱化日趋严重,本文特选取该生境优势植物,即一年生或多年生单子叶和双子叶草本植物羽茅、麻花头和飞廉作为研究对象,以不同埋深和不同培养天数的种子萌发行为和子叶生长作为研究内容,探析植物在保持生存的情况下对自身发展进行适应性的调整,实现对有限的资源进行合理分配的过程,为研究种群的生态适应机制提供理论基础[6].
1 材料及方法
1.1 实验材料及处理方式
用30~35℃温水将羽茅、麻花头、飞廉的种子浸泡2~6 h,使种子吸水充分.每30颗种子为一组放在一个透明的塑料杯中,分别进行3种埋深处理,分别是0 cm、0.5 cm、3 cm,每种处理设置3组重复,并放在培养箱内培养,培养箱条件设置为变温25/15℃,光照/黑暗为12/12 h[7-8].
待种子萌发的子叶露出土面,分别在种植5 d、15 d、30 d后,将新鲜子叶从实验沙中拔出,进行子叶的动态形态监测和酶活性监测.
1.2 种子活力的检测
以浓度为9.078 g/L的KH2PO4为母液Ⅰ,浓度为9.472 g/L的Na2HPO4为母液Ⅱ,将母液Ⅰ和母液Ⅱ以2∶3的比例配制,得到500 mL pH为7.5的磷酸盐缓冲液后,称量TTC(2,3,5-氯化三苯基四氮唑)0.5 g,将TTC溶解于以上磷酸盐缓冲液中,得到浓度为0.1%的TTC溶液,最后调节pH为7.0.
随机取10粒种子并沿种胚中央小心切开,将其中一半放入培养皿中,皿中倒入TTC溶液,浸泡种子,在恒温箱(35℃)中保温30 min后,弃TTC溶液,并用清水冲洗种子2次,观察并记录种胚染色的程度[9].
1.3 种子干重的测定
设置50粒种子为一组,称量滤纸重量并记录后,再放上种子,通过称量和计算得到种子鲜重.用笔在称量滤纸上标清种子类别,包裹后放入烘箱中在130℃下烘烤40 min停止,重复至少3次,直至数据不再发生变化为止[10].
1.4 种子发芽
将新鲜的种子在变温25/15℃,光照/黑暗为12/12 h条件的培养箱中进行培育,用蒸馏水湿润,记录每天种子的发芽情况.计算0~30 d的发芽率,对比不同埋深对发芽的影响.
1.5 SOD酶活性
取新鲜的羽茅、麻花头、飞廉种子各0.5 g,及5 d、15 d、30 d的羽茅、麻花头和飞廉的新鲜子叶,每组至少3株,放入研钵中.
每个研钵按每克鲜叶加入3 mL浓度为0.05 mol/L、pH为7.8的磷酸钠缓冲液的标准加入磷酸钠缓冲液,并加入适量的石英砂,置于冰上研磨成匀浆,然后将匀浆液转移到5 mL离心管中,8 500 rpm、4℃条件下离心30 min,所得上清液即为SOD酶粗提液[11].
向每组依次加入磷酸钠缓冲液,蛋氨酸-磷酸钠缓冲液、NBT溶液、酶液及核黄素溶液,并充分混匀.对照组试管置于暗处,剩余7个试管均放在25℃,光强为4 500 Lux的光照箱内照光15 min立即遮光,终止反应,于560 nm波长下测定吸光值[11].
1.6 淀粉酶活性
取新鲜的羽茅、麻花头、飞廉种子各0.5 g,及5 d、15 d、30 d的羽茅、麻花头、飞廉的新鲜子叶,每组至少3株,放入研钵内.
加入2 mL蒸馏水并放入少许石英砂,研磨成匀浆,再用3 mL蒸馏水清洗研钵,将清洗得到的液体一同倒入离心管并摇匀,室温中放置15 min,在4℃、10 000 rpm条件下离心30 min,留上清液.
取3支试管,1支为对照管,2支为测定管,每管各加0.5 mL以上提取所得酶液,于70℃恒温水浴锅中先加热15 min,随后取出迅速转移到冰中,待其彻底冷却后,向各管再加入柠檬酸缓冲液.对照管中加入2 mL的3,5-二硝基水杨酸,并于40℃恒温水浴中保温15 min,然后向各管加入预热40℃的1%的淀粉溶液摇匀,继续放置在40℃水浴锅中,接着向测定管中快速加入3,5-二硝基水杨酸以终止酶的活力,并于100℃煮5 min,冷却后加入10 mL蒸馏水,立即于540 nm波长下测定吸光值[12].
1.7 数据分析
使用Microsoft Excel 2013计算最终发芽率.数据分析采用GraphPad Prism 9处理,结果表示为平均值和标准差(P<0.05).
2 结果与分析
2.1 不同埋深对种子萌发行为的影响
羽茅、麻花头和飞廉种子含水量会影响种子的基础萌发能力,羽茅种子含水量12.49%±0.02%,麻花头种子含水量13.10%±0.04%,飞廉种子含水量15.25%±0.03%.飞廉种子含水量较高,其萌发率相对较低.
土壤埋深是植物生长的一个重要环境因素,不同的埋深条件可能导致植物采取不同的生长策略.种子萌发行为因为埋深不同而不同,结果显示,当埋深处于0.5 cm时,种子萌发行为较突出,发芽率较高,此时飞廉种子发芽率为50%,在0 cm埋深处理及3 cm埋深处理的发芽率分别为10%和3.3%.羽茅在0.5 cm埋深时的发芽率最高,达93.3%,其发芽率最低的是0 cm埋深处理,为26.7%.而麻花头比较特殊,在埋深0 cm时发芽率最高,为60%,与0.5 cm埋深的差异不大.
由以上结果可见,种子埋过深或过浅都不利于发芽,埋深过浅种子可能缺乏保护,容易被风干,埋深过深可能阻碍种子吸收营养和水分,使种子遭受过重的压力,导致发芽失败.所以只有选择合适的埋深深度,才能确保种子顺利发芽.种子萌发是植物群落演替和更新的基础,对维持生态系统的稳定性和健康至关重要.理解种子在不同深度的土壤中的萌发行为有助于预测和控制植被的分布和丰度,进一步影响整个生态系统的结构和功能.此外,种子萌发是植物生命周期的重要阶段.研究种子在不同深度的土壤中的萌发行为有助于揭示植物如何适应不同的土壤环境.
2.2 不同埋深对幼苗形态指标的影响
在种子萌发成幼苗的初期,子叶发挥着至关重要的作用.检测子叶的根、茎长度对于评估幼苗的生长状况、健康状况、预测生长潜力等方面具有重要意义.因此,本研究测量不同埋深羽茅、麻花头和飞廉幼苗根、茎长度,结果分别如图1、图2、图3所示.随着埋深的增加,羽茅、麻花头和飞廉子叶的根长和茎长都呈现出显著的增长趋势(P<0.05).
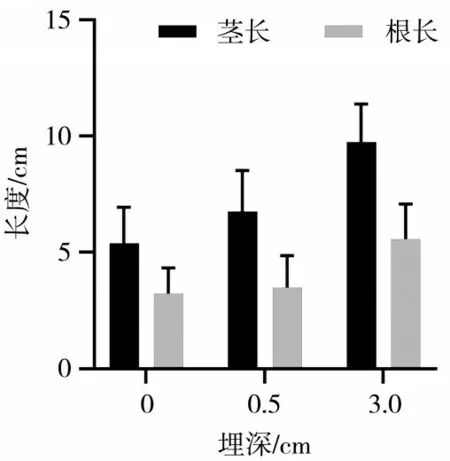
图1 不同埋深羽茅子叶根、茎长度差异(P<0.05)
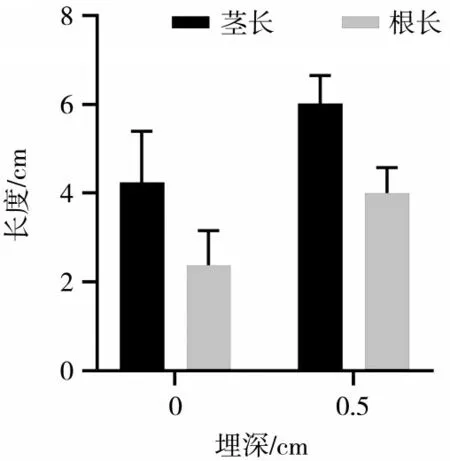
图2 不同埋深麻花头子叶根、茎长度差异(P<0.05)

图3 不同埋深飞廉子叶根、茎长度差异(P<0.05)
通过观察子叶根、茎长度的变化,发现一定的基质压力对子叶的生长具有积极的促进作用,可以有效地促进子叶根、茎的生长,从而帮助子叶顺利破土而出.了解不同埋深对植物子叶根、茎长度的影响,可以为种植者提供科学依据,指导他们合理安排种植深度.
2.3 不同时期不同埋深对子叶功能的影响
子叶可为幼苗提供必要的营养物质,确保幼苗正常生长,子叶还能通过自身的生长促进幼苗的发育,为其提供一个安全、适宜的生长环境.通过对种子淀粉酶活性及SOD酶活力进行测定,羽茅种子淀粉酶活力为4.65 mg/g,种子SOD酶活力为60 000 U/g,麻花头种子淀粉酶活力为2.706 mg/g,种子SOD酶活力为240 000 U/g,飞廉种子淀粉酶活力为14.49 mg/g,种子SOD酶活力为333 333.33 U/g.
子叶和种子中的淀粉酶活性可以提供有关植物生长和发育的重要信息.淀粉酶是植物中负责分解淀粉的酶,其活性水平可以反映植物对淀粉的利用效率和生长状况.羽茅、麻花头和飞廉子叶淀粉酶活力检测结果如图4、图5、图6所示.
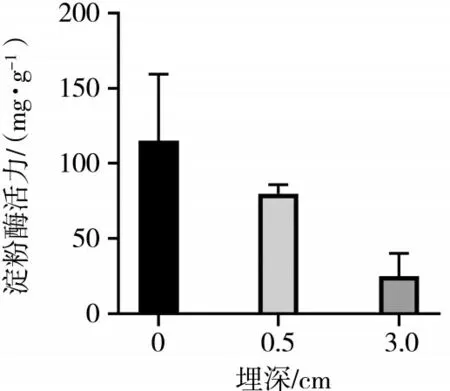
图4 不同埋深不同时期的羽茅子叶淀粉酶活性差异(P<0.05)
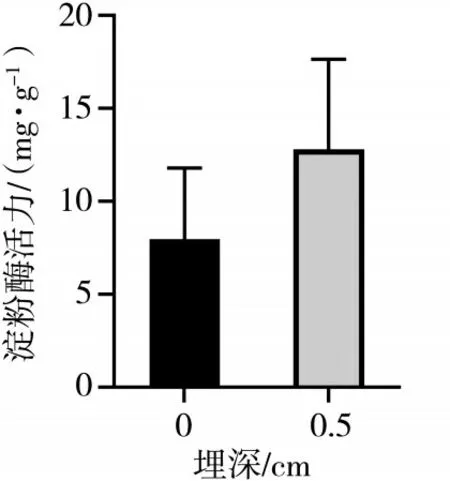
图5 不同埋深不同时期的麻花头子叶淀粉酶活性差异(P<0.05)
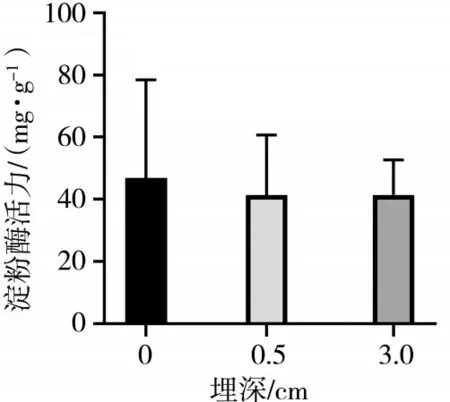
图6 不同埋深不同时期的飞廉子叶淀粉酶活性差异
从图4、图5、图6可以看出,羽茅子叶在浅层土壤中的淀粉酶活性相对较高,但随着土壤埋深的增加,这种活性逐渐降低.该变化表明,羽茅子叶在较深的土壤环境中,由于氧气缺乏、湿度增加等环境压力,导致淀粉酶的活性降低,从而减少了对淀粉的分解和利用.相反,麻花头子叶的淀粉酶活性随着土壤埋深的增加而显著升高.这表明麻花头子叶为了适应深层的低氧、高湿环境,是通过提高淀粉酶的活性增加对淀粉的分解和利用,以获取更多的能量和营养物质.飞廉子叶的淀粉酶活性变化趋势与羽茅相近.值得注意的是,无论是羽茅、麻花头还是飞廉,其子叶的淀粉酶活性均高于种子阶段的淀粉酶活性.这一结果进一步凸显了子叶在植物生长过程中的重要作用.在种子萌发和幼苗生长阶段,子叶作为重要的能量来源,需要更高的酶活性支持其快速生长和发育.
SOD酶是植物代谢的关键酶,其可以保护细胞免受氧化损伤,维持植物正常的代谢和生长.羽茅、麻花头和飞廉子叶SOD酶活力检测结果如表1、表2、表3所示.
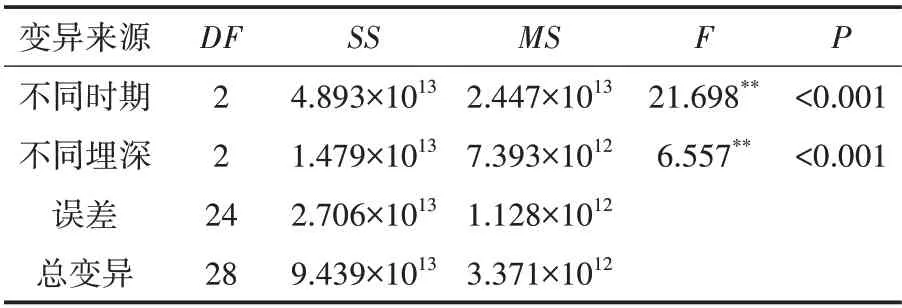
表1 不同埋深不同时期羽茅子叶SOD酶活力影响的方差分析表

表2 不同埋深不同时期麻花头子叶SOD酶活力影响的方差分析表
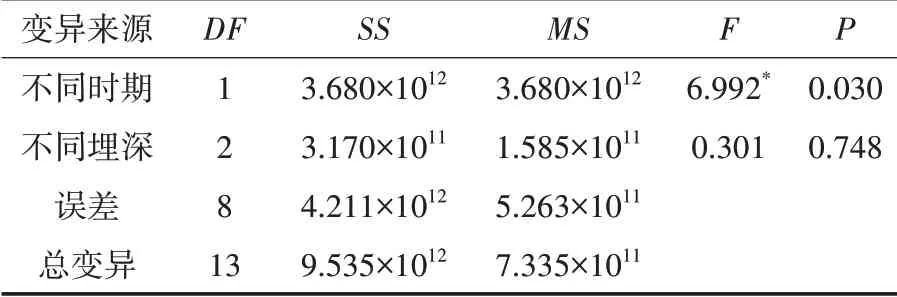
表3 不同埋深不同时期飞廉子叶SOD酶活力影响的方差分析表
从表1、表2、表3可以看出,羽茅和麻花头子叶的SOD酶活力受时间和埋深的影响极为显著.飞廉子叶的SOD酶活力在不同时期也有显著的变化,但与埋深的关系不大.
此外,幼苗的根、茎长度与淀粉酶活性大小之间存在反比关系.即初期的种子萌发行为越强,子叶的淀粉酶活性越弱.以上变化趋势提示,随着萌发的进行,子叶开始生长,需要更多的能量和营养物质支持其发育.如果淀粉酶活性过高,会导致淀粉过度降解,产生的葡萄糖过多,这可能会对子叶的生长产生负面影响.因此,子叶可能通过调节淀粉酶活性来平衡能量和营养物质的供应,以满足自身生长的需求.这种平衡机制对于种子的正常萌发和幼苗的生长具有重要意义.
3 结论与讨论
种子需要吸收和利用有限的资源,例如水分、养分和能量,以完成萌发并成长为健康的植物[13].在种子萌发的过程中,子叶中的养分和能量被用于支持种子的生长和发育.因此,在资源有限的情况下,种子萌发和幼苗生长之间存在一种资源竞争的关系.在这种情况下,如何合理地分配资源成为一个重要的问题.种子和子叶之间的资源分配可能会受到多种因素的影响.例如,在干旱条件下,种子可能需要更多的水分支持萌发,子叶中的水分可能会被更多地用于支持种子的生长.此外,种子的萌发和子叶的功能之间也存在相互作用.子叶中的养分和能量不仅被用于支持种子的生长,同时也对种子的萌发过程产生影响.
因此,本研究以羽茅、麻花头、飞廉等子叶出土型单子叶和双子叶植物为研究对象,设置不同生境如设置埋深为0 cm、0.5 cm、3 cm条件下,分别监测不同发育时期种子、子叶和幼苗的形态和生理指标,监测酶活性的变化,探讨相关植物如何通过合理的资源管理来提高种子的萌发率和植物的生长质量.结果发现,一定的基质压力可促进种子萌发,压力过小或过大皆不利于子叶破土.除麻花头外,子叶茎长与淀粉酶活性大小成反比关系:种子萌发行为越强,对应子叶的淀粉酶活性越弱.淀粉酶活性反应植物体内能量代谢的水平,这一结果表明,在有限的资源条件下,植物种子的萌发行为和子叶功能在资源利用和能量输出之间存在一定的权衡关系,即子叶通过调节淀粉酶活性来平衡能量和营养物质的供应,以满足自身生长的需求.
资源的权衡涉及到植物如何根据环境条件和自身需求在能量供应上进行优化和平衡.子叶作为种子内储存养分的器官,对于调节资源的分配和供应起着关键作用.该结论强调了子叶通过调节淀粉酶活性来平衡能量和营养物质的供应,从而满足自身生长的需求.这种调控机制可能涉及多种因素,如环境信号、激素调节和基因表达等,这需要进一步研究来深入了解其背后的机制和影响因素.该结论不仅增加了对子叶功能的理解,而且提供了研究资源权衡过程的新视角,同时具有一定的实际应用价值.了解子叶如何通过调节淀粉酶活性来平衡能量和营养物质的供应,可以更好地理解植物的生长规律和应对环境变化的能力,为通过改进种子处理、调整种植条件、开发新型肥料,提高作物的生长和产量等农业生产的优化提供依据.此外,通过监测植物各阶段幼苗形态及生理学指标,分析不同种子萌发和子叶生长的相关性,有助于认识和阐明物种进化及其生态适应,对于预测植物的发芽和出土、全球变暖对植物生长发育及分布的影响、认识野生植物资源利用策略等均具有理论指导意义和生产实践价值.
