植被根系对土体水力学特性影响的现状与定量分析
2024-04-29杨几肖涛孙靓史亚瑞李萍
杨几 肖涛 孙靓 史亚瑞 李萍

收稿日期:2023-09-10
基金項目:国家自然科学基金青年项目(42007251)。
第一作者:杨几,男,从事生态岩土工程方面研究,yj811011@163.com。
通信作者:李萍,女,副教授,从事非饱和土力学研究,20175080@nwu.edu.cn。
摘要 在全球气候变暖和“双碳”战略的背景下,植被护坡成为一种潜在的碳负排放边坡加固和侵蚀控制手段。植被根系的发育可以改变土体结构和水力学特性,进而影响边坡的稳定性。虽然一些试验研究已经探讨了特定条件下植被对土体水力学特性的影响,但是对于植被根系改变土体水力学特性的一般规律尚不清楚。该文从植被蒸腾作用、根系物理作用以及根系生物化学作用3个方面入手,收集并梳理了近年来国内外相关研究,分析并总结了植被根系改变土体水力学特性的作用机理,且对已有试验数据进行了统计分析。指出植被根系对土体水力学特性的改变取决于植被蒸腾、根系物理作用和根系生物化学作用三者的相对贡献,同时受根系类型、土体类型及生长阶段等因素的影响。因此,植被根系对土体水力学特性的影响十分复杂,更全面广泛的研究对于揭示植被护坡机理是亟需的。最后,探讨了植被根系影响土体水力学特性的研究空白和未来趋势。
关键词 植被根系;SWRC;渗透系数;土体吸力;蒸腾作用
中图分类号: TU43 DOI:10.16152/j.cnki.xdxbzr.2024-01-007
Current state and quantitative analysis of the effects of vegetationroots on the soil hydraulic characteristics
YANG Ji, XIAO Tao, SUN Liang, SHI Yarui, LI Ping
(State Key Laboratory of Continental Dynamics, Department of Geology, Northwest University, Xian 710069, China)
Abstract Under the background of global warming and “carbon peaking and carbon neutrality” strategy, vegetation slope protection has become a potential means to reinforce the slope of negative carbon emission and errosion control. The development of vegetation roots can change the soil structure and hydraulic characteristics, and then affect the stability of slope. Although some experimental studies have explored the effects of vegetation on soil hydraulic characteristics under specific conditions, the general law that vegetation roots change soil hydraulic characteristics under different conditions is not clear.In this paper, the research on vegetation transpiration, root physical function and root biochemical function are collected and reviewed, the mechanism of the effect of vegetation roots on soil hydraulic characteristics is analyzed and summarized, and the available experimental data are statistically analyzed. It is pointed out that the change of hydraulic characteristics of soil by vegetation roots depends on the relative contributions of transpiration, physical and biochemical processes of roots, at the same time by the root type, soil type and growth stage and other factors. Therefore, the effect of vegetation roots on soil hydraulic characteristics is very complex, and more comprehensive and extensive researches are needed to reveal the mechanism of vegetation slope protection. Finally, the blank and future research trend of the effect of vegetation roots on soil hydraulic characteristics are discussed.
Keywords vegetation roots; SWRC; hydraulic conductivity; soil suction; plant transpiration
土体水力学特性在土壤-植物-大气连续体物质循环(如水循环和碳循环)过程有着至关重要的作用。它不仅控制着土中水的运移,而且影响径流的形成[1]。影响土体水力学特性的因素有很多,如土体结构(粒径组成、孔径分布、有机质含量等)、人类活动(农业实践和工程扰动)、季节变化和植被覆盖等[2-3]。其中,植被根系可以通过各种物理、化学和生物作用改变土体结构,进而改变土体的水力学特性。然而,植被根系对土体水力学特性的改变并非单纯地增强或减弱,这取决于植物类型、土体结构、土壤环境和大气环境等因素[4-5]。尽管目前大部分工程边坡和垃圾填埋场覆盖层都通过种植植被以达到美化环境、恢复生态以及提高土体稳定性的目的[6-7],然而,植被根系对土体水力学特性和基础设施稳定性影响的研究仍然较为缺乏。已有研究报道了植被根系对土体水力学特性的影响和机理。首先,植被蒸腾作用促使根系吸水,增加土体吸力,提高土体持水能力并降低土体渗透系数[8]。植被生长过程中根系的一系列扩张、填充土体孔隙以及穿透、包络集粒改变了土体的孔径分布和粒径分布,继而影响土体的水力学特性[9-10]。植被根系还可能通过改变土体物质(固态或液态)成分进而影响土体的水力学特性[11],如根系分泌物能够增加土体的水稳性、影响土体的浸润性和流体黏度等,从而影响土体的水力学特性[12]。尽管目前对特定植物类型、特定土壤和大气环境下植被根系对土体水力学特性的暂态影响有了一些定性或定量的认识,但是对于植被根系引起土体水力学特性变化的定量研究仍较为缺乏,这使得很多研究领域无法有效考虑植被根系引起的土体水力学特性变化[13]。
本文总结和分析了近年来的一些相关研究和数据,以期为水文、岩土、地质、生态和环境等相关领域了解植被根系影响土体水力学特性的研究现状提供便利,最后,探讨了植被根系影响土体水力学特性方向的研究空白和未来方向。
1 土体水力学特性
土体水力学特性控制土中水分运移,包括持水特征曲线(SWRC)、饱和渗透系数(Ks)和非饱和渗透系数(K)等。SWRC是描述含水率(或饱和度)与基质吸力(或吸力)的本構关系,是土体中水分与土体中能量平衡状态的定量表征。SWRC通常利用数学模型拟合有限实测数据点得到。饱和渗透系数是表征流体通过多孔介质难易程度的物理量,反映了多孔介质的渗透能力,一般通过常水头或变水头试验获得。而非饱和渗透系数与有效饱和度(或吸力、含水率)有关,描述二者的函数称为渗透性函数。在进行渗流分析时,SWRC和渗透性函数不可或缺,但是非饱和渗透系数的直接测量耗时费力,因此常常通过模型预测。目前,描述SWRC和预测渗透性函数最常用的模型是Van Genuchten-Mualem模型,其表达式如下[14],
θ(Ψ)=θr+θs-θr[1+(α(Ψ)n]m,Ψ<0
θs,Ψ≥0(1)
K(Ψ)=KsSle[1-1-S1mem]2(2)
Se=θ-θrθs-θr(3)
式中:θs和θr是饱和含水率和残余含水率;Ψ是吸力,单位为kPa;α(kPa-1)、m、n为拟合参数;m=1-1/n(n>1);K和Ks分别为非饱和渗透系数和饱和渗透系数,单位为m/s;Se为有效饱和度;l为土壤孔隙连通性参数。
2 植被根系的影响机理
植被根系对土体水力学特性的影响主要体现在3个方面。① 植被蒸腾引起的根系吸水。一方面,根系吸水增加根周围土体的吸力,影响土体的水力学特性;另一方面根系吸水使根系周围土体经历更多的干湿循环,可能导致土体结构发生变化,进一步影响土体水力学特性。②植被根系的物理作用,主要是根系通过扩张、穿透、孔隙填充和集粒包络等方式改变土体孔径分布和粒径分布,致使土体水力学特性发生改变。③植被根系的生物化学作用,即植物生理活动会影响土体的物质成分和微生物活性,如地上茎、叶和地下根系的腐败及根系分泌物的释放等使土中有机物含量(SOC)发生变化,土体水力学特性随之而改变。
2.1 植被的蒸腾作用
土体水分蒸发蒸腾损失总量(ET)主要分为地面的物理蒸发损失(E)以及植被的蒸腾损失(T),它是陆地水循环中最大的水通量,其中植被蒸腾是主要部分[15]。已有研究表明,ET约占陆地降水量的55%~65%,其中,T占ET的60%左右[16-17]。蒸腾作用是水通过管胞或导管运输,最终以水蒸气的形式从植物表面进入大气的过程[18]。对于大部分植物而言,绝大多数水分是通过叶片表面散失的,而叶片的气孔作为大气与植物之间的阀门,控制着叶片内外的碳水交换以及植物的蒸腾作用和光合作用[19]。相关报道表明,气孔蒸腾约占总蒸腾量的90%以上,因此,气孔的大小和数量是植物蒸腾作用强弱的决定性因素,气孔蒸腾也是植物叶片蒸腾的主要形式。此外,影响植物蒸腾作用的因素还有许多,如植物类型、土体类型、光照强度、大气湿度、CO2浓度等[20-22]。
植被蒸腾引起的根系吸水能够改变土体的水力学特性[23-24]。首先,植被根系从土中吸收水分,使土体含水率降低,土体吸力随之增加。研究结果显示有蒸腾作用的植被土的吸力明显大于裸土,吸力为裸土的1~8倍[25]。这意味着在相同吸力下,植被土能够吸收更多水分,即植被蒸腾引起的根系吸水提高了土体的持水能力。另外,由于吸力增加,土体的非饱和渗透系数减小[8]。事实上,土体吸力与根系吸水速率是息息相关的。如Feddes等人提出的蒸腾还原函数(TRF)描述了蒸腾速率和土体吸力的关系,是研究土壤-植物-大气之间水分运移的基础函数[26]。由于不同植物种类其TRF不同[27],因此,在使用植被进行护坡或保水时必须考虑土壤环境和植物种类,以最大限度地发挥植被的水力加固作用[4-8]。此外,植被蒸腾使根际土经历更多的干湿循环,可能导致土体开裂,且在干旱期间,植被根系收缩,导致根土分离(特别是直根型根系)。再次降雨后,这种收缩难以很快恢复,则可能形成优势通道,促进水分入渗[28]。简而言之,植被根系吸水可以增加土体吸力,直接改变土体水力学特性,或通过改变土体结构间接改变土体水力学特性[29-30]。
2.2 植被根系的物理作用
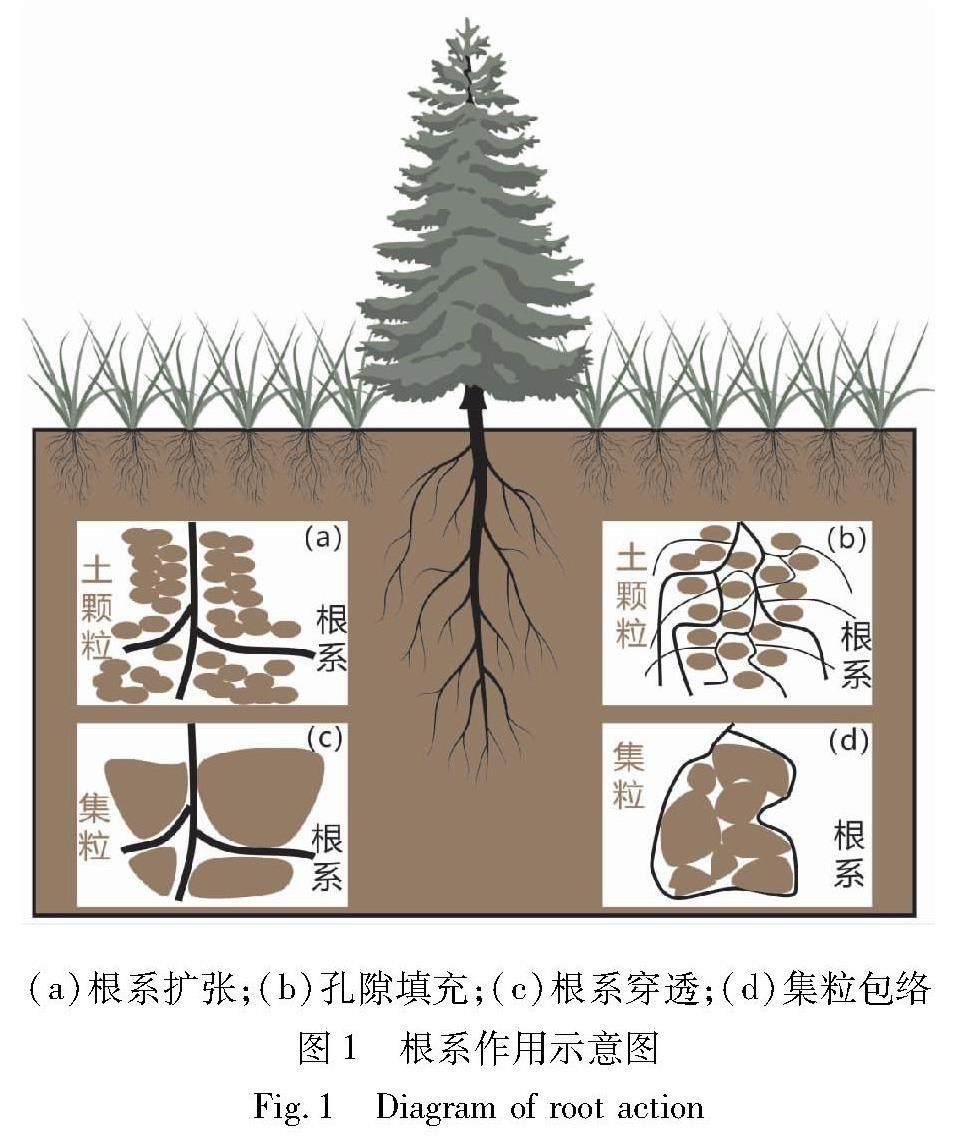
植被根系的物理作用主要体现在根系对土体孔径分布和粒径分布的改变,从而致使土体的水力学特性发生改变[9-10]。图1展示了根系物理作用的主要过程,包括根系穿透、根系扩张、孔隙填充和集粒包络等。根系扩张指随着植物的生长,根径增长,引起土颗粒重新排列或压缩周围土体的过程,如图1(a)所示。细根由于强度较低难以造成土颗粒的移动,往往沿着已有孔隙生长,最终导致孔隙率的减小,即孔隙填充[10],如图1(b)所示。根系扩张和孔隙填充作用主要通过改变土体孔径分布来影响土体水力学特性,而根系穿透和集粒包络则是通过改变集粒尺寸来改变土体水力学特性。根系穿透和集粒包络作用能够使颗粒(或集粒)破碎、分散或聚合,如图1(c)和图1(d)所示。随着根系生长,较大的集粒可能被穿透,分散为较小的集粒,而小集粒可能在细根的包络下聚合形成较大的集粒。
植被根系的物理作用对土体水力学特性的影响十分复杂,这与土体类型[31]、根系构型[4]以及生长阶段[13]密切相关。在粗粒土中根系填充作用较为显著,导致大孔减少,通常使土体渗透性降低[31]。而在细粒土中根系穿透和包络作用更为显著,可能导致土中大孔和微孔增多[9-10]。除此之外,不同的根型对土体水力学特性的影响不同。直根型根系往往向深层土体生长, 在经历多次干湿循环后,根系收缩和土体开裂形成的优势通道会导致土体渗透性增加。须根型根系则大多填充土体中原有的孔隙和裂缝,使得土体渗透性降低[32]。另外,根系对土体水力学特性的影响会随时间变化,植物在不同生长阶段表现出不同的根系物理作用,当根系不断伸长、数量增多,土体水力学特性会随根长密度(RLD)和根系生物量(RBV)的变化而变化[29]。
2.3 植被根系的生物化学作用
植被根系的生物化学作用是通过影响土体的物质成分和微生物活动进而影响土体水力学特性。植物会增加土的SOC,主要是以向土中释放根系分泌物(如糖类、氨基酸、有机酸、磷脂和果胶型多糖等)和植物器官(根、茎、叶)衰亡分解的方式。根据Tisdall and Oades提出的集粒多级形成机理[33]可知,有机物具有胶结土颗粒的作用,且有机物为土中微生物提供必要的养分,促进微生物产生菌丝,以形成更大、水稳性更强的集粒。此外,高分子有机聚合物可与土中矿物形成矿物有机物复合体(MAOM),从而改变土体粒径和孔径分布,提高土体的水稳性[32]。
值得注意的是根系分泌物,特别是黏液,是植物分泌的一种高分子量化合物、富含多糖的黏弹性凝胶,是影响根际土体物理化学性质的主要因素之一[34-35]。目前,绝大多数的研究表明根系分泌物会增加土体的持水能力、降低土体的渗透系数[36-39]。这些研究把根系分泌物改变土体水力学特性归因于以下4点:①黏液能够吸收大量水分,饱和时甚至能达到干重的数百倍[40],进而增强土体持水能力;②根系分泌物中黏液和多糖等物质会黏结土颗粒,改变土体粒径和孔径分布,增强土体水稳性[41];③根系分泌物会增加流体黏度,且影响土体的浸润性[40];④根系分泌物刺激土中微生物活性,微生物本身或代谢物可堵塞孔隙,即Bioclogging[42],或黏结土颗粒。然而也有一些研究认为根系分泌物会降低土体的持水能力[43-45],他们把这种负面作用归因于根系分泌物中的卵磷脂等脂质充当了表面活性剂,降低了弯液面的表面张力。但是随着土体基质吸力的增大,这种表面活性剂的作用会随之降低。因此,植被根系的生物化学作用对土体水力学特性的影响可能需要考虑有机物的物质成分、官能团种类、有机物之间的相互作用等。
3 植被根系影响的定量分析
如前所述,植被根系对土体水力学特性的影响是复杂多变的,不仅有植被蒸腾作用,还有根系的物理作用与生物化学作用。附表1总结了部分相关研究,着重考虑了根系构型、土体类型以及植物年龄3个影响因素,统计分析了相关试验数据。将砂粒含量大于50%的土划分为粗粒土,小于50%的则为细粒土[46]。根系构型分为直根型和须根型,大多数双子叶植物如乔木、灌木以及大豆等作物均属于直根型根系,而单子叶植物如大多数草本以及小麦等作物属于须根型根系。通过VG模型〔式(1)〕拟合SWRC数据得出θs、α、n这3个参数。
3.1 植被根系对SWRC的影响
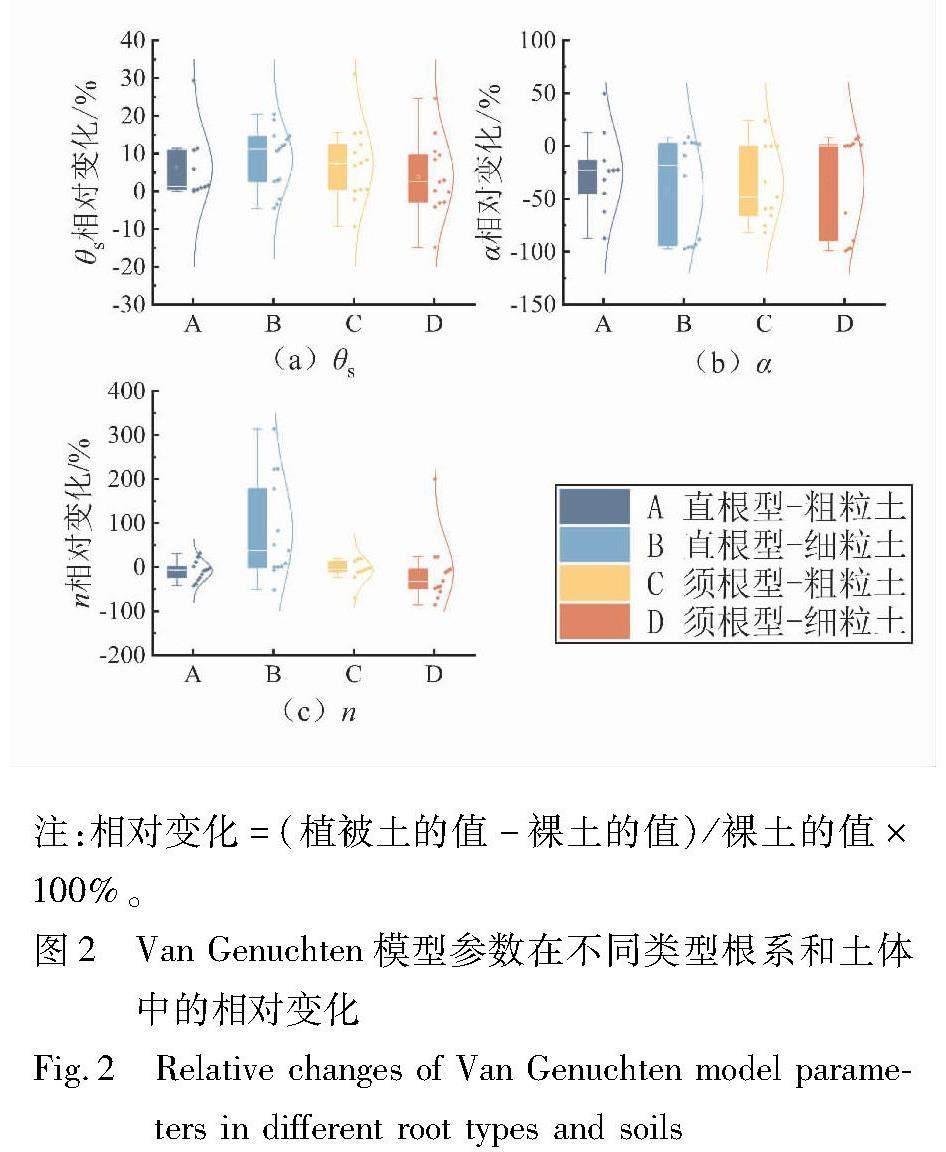
图2(a)~(c)分别展示了θs、α、n在考虑植物作用后其值的相对变化〔(植被土的值-裸土的值)/裸土的值×100%〕。从图2(a)可以看出,无论是直根型还是须根型,粗粒土和细粒土中θs相对变化的平均值都大于0,这意味着在所选择案例中根系造成的孔隙形成(pore-forming)作用较孔隙堵塞(pore-clogging)作用显著得多。这与Lu等人[29]和Leung等人[47]得出的结论一致。另外,植被土θs的增加也与根系限制因土体收缩而导致的孔隙損失有关。对于粗粒土中的直根型,所有案例中θs的相对变化都大于0,也就意味着植被根系使得土体孔隙体积增大。如前所述,很可能是因为直根型根系的收缩以及腐败导致大孔隙的形成,从而提高了土体孔隙率和θs。Bodner等人的研究表明,根系生长到一定密度后,密集的须根使大孔隙率增加了约16%,而直根型由于其较大的粗根比例使大孔隙率增加了近30%[48]。因此,相较于须根型对θs的增加(5%),直根型对θs的增加程度更高(7%)。另外,粗粒土中θs增加了约6%,且只有3组数据表现为减小,而细粒土中θs增加了5%,有8组数据呈现出减小的趋势。因此,总的来说,粗粒土中直根型根系能够增加土体的θs。
参数α与土体的进气值密切相关,α越小则进气值越大,能够反映土体的持水能力[29]。从图2(b)中可以看出,考虑植被根系的影响时,绝大多数α表现为减小,但在不同根系构型和土体类型中减小的程度不同。直根型和须根型根系使细粒土的α平均下降了40%和32%,在粗粒土中则下降了24%和36%。α减小(即进气值增大)的原因可能是:①根系的填充作用减小了土体的平均孔隙;②根系分泌物的强亲水性和膨胀使得孔隙体积减小。图2(b)中,B和D的α增加案例分别为5例和7例,A和C的则分别为1例和2例。也就是说不考虑根系类型时,粗粒土中根系使α降低的可能性更大。这是因为粗粒土的大孔隙较多,根系的生长减少了大孔隙,使得土体的平均孔径减小,从而进气值增大。而对于细粒土,由于植被蒸腾吸水会使土中产生裂缝,从而增加了土体的平均孔径,导致进气值减少[30]。不同根系类型对α值的影响也不同。有研究表明,粗根和细根生长到一定根系密度后会表现出对孔隙改变的不同效应[48]。相较于直根型,粗粒土中的须根型使得α值的下降幅度最大,原因是根系的填充使大孔隙向中小孔隙转化。
参数n对应SWRC过渡段的斜率,与孔径分布密切相关,n值越小意味着孔径分布较广,n越大则土体孔径更均一[49]。从图2(c)可以看出,考虑根系作用时,除了B的n值增加了81%之外,A、C、D的n值均表现为减小,分别减小了8%、7%和12%。B中n值的增加可能是因为根系吸水引发干湿循环和直根型根系收缩导致的大孔隙增加,使细粒土中孔径分布趋于均匀。而根系穿透、扩张和包络等物理作用以及生物化学作用使得土体的孔径分布更广,导致A、C、D的n值减小。多位学者观察到植被导致土中孔隙各向异性的增加[8,47,50],这都合理解释了n值的减小。
3.2 植被根系对饱和渗透系数的影响
Ks控制着降雨入渗量以及地表径流量,是水文模型中的关键参数。Ks的相对变化为-93%~1 425%之间,如图3所示,除了C之外,A、B和D的Ks均增加,增加幅度最大的为B。在植被根系作用下,15组数据中有11组的Ks增加。Ks增加一方面是因为蒸腾吸水引起干湿循环和根系收缩形成了优势通道,另一方面则是根系包络以及有机质促进土颗粒聚集形成集粒,集粒间孔隙也为水分快速迁移提供了通道。须根型粗粒土中Ks减小则是根系的填充使土体中大孔隙减小、连通性降低造成的,其次可能与根系分泌物的高黏度和生物堵塞有关。与须根型相比,直根型根系对Ks的增加更为明显。这是因为直根型根系能够导致更大的孔隙率。另外,细粒土中Ks增加更明显,这是因为细粒土中根系的孔隙形成作用胜过孔隙填充作用,导致了Ks的增加。
此外,植被在不同生长阶段对土体渗透性的影响是不同的。如图4所示,Ks随植物生长年龄表现出不同的变化,其中,图4(b)、 (c)、(f)中Ks随RLD逐渐增大。这可以归因于根系生长初期以蒸腾吸水引起干湿循环和根系收缩形成优势通道为主,而随着树龄的增加,根系腐败形成的大孔隙和有机质积累形成的集粒粒间孔隙发挥了主要作用,从而致使土体渗透性增加。然而RLD的增加并不总是意味着Ks的增加,它与根系构型和土体类型有关。图4(a)、(h)中Ks随RLD和RBV表现出先增大后减小的趋势,其中,图4(a)为直根型粗粒土,图4(h)为须根型细粒土。前期Ks增大的原因与第1种情况类似,而当RLD和RBV增加到某一个阈值时,Ks转而减小[30,51]。与第1种情况不同的原因是某些构型的根系会在原先穿透留下的孔隙通道以及包络形成的粒间孔隙生长,优势通道转变为孔隙堵塞,阻碍了水分的流动〔见图1(c)和(d)〕。
另一种情况是Ks随RLD和RBV表现出先减小后增大的趋势〔如图4(d)、(e)、(i)〕。出现这种趋势主要是因为图4(d)、(e)、(i)均为粗粒土,并且所研究的植被根系较细,在早期生长阶段以孔隙堵塞作用为主,然而随着植被生长,根系的孔隙堵塞作用由于可供填充的孔隙变少而减弱,根系收缩和腐败逐渐占主导地位。另外,由于生长时间长,土壤中有机物积累,微生物活性高,这些作用可以形成更稳定、复杂的土壤结构[11]。图4(g)中Ks随RBV持续降低,可能是由于含砾砂土本身孔隙较大,且植物根系为须根型,进而以孔隙填充作用为主。另外,根系分泌物导致的液相黏度增加也阻碍了水分的迁移。综上所述,这些复杂的变化都表明了植被对土体Ks的改变是植物蒸腾吸水、根系物理化学作用、根土相互作用等多方面综合作用的结果,因此,总结植被根系对土体水力学特性影响的一般规律仍然比较困难。
3.3 植被根系对非饱和渗透系数的影响
通常植被根系的生长区域处于包气带上部,包气带土体处于部分饱和或非饱和状态,因此,研究根系对非饱和渗透系数(K)的影响是十分有必要的[55]。许多研究表明,植物的生长能够提高相同吸力水平下土体的K(如附表1中案例1、4、5、 7、 11、 13)。但是,也有研究表明根系的存在会减小相同吸力水平下的K(如附表1中案例3、23、38)。还有研究表明根系主要影响低吸力水平(0~30 kPa)的K[30,56]。因此,为了探究植物生长对K的影响,需要考虑根系构型、土体类型和吸力大小等因素。由于K随着吸力的增大而减小,并且大部分研究只测定了低吸力下的K,如图5所示。因此,我们只考虑低吸力范围内K的相对变化。如图5(b)、(f)所示,相比于裸土,K由于直根型根系的生长而增加,即根系吸水虽然导致了土体吸力增加,但根系收缩和土体开裂形成的优势通道,以及根系一系列物理、生物化学作用产生的大孔隙为渗流提供了通道。而须根型根系通常导致K的降低〔见图5(c)、(d)、(e)〕,但在须根型-粗粒土中〔见图5(a)〕,K增加可能由于细根的迅速腐败[57]。
须根型根系使得土体K值降低,如图5(c)和5(d)所示,与Ks的变化一致〔见图4(g)〕,是须根型根系造成的孔隙填充和根系分泌物导致的孔隙液体黏度增加占主导地位的结果。同时,这种變化也与植物年龄有关,树龄越大,K下降的幅度越大〔见图5(c)、(d)〕,这可能是因为土壤结构复杂化导致渗流路径增长。图5(e)与之前分析不同的原因主要是相较于裸土,须根型根系显著限制了细粒土的开裂,导致K减小。除了土体类型影响外,直根型和须根型根系导致细粒土K值的变化也表现出相反趋势〔见图5(e)、(f)〕。由此可见,正是植被的蒸腾、根系的物理化学作用和吸力等因素的相互作用以及不同的土体类型、植被类型导致了K值的复杂变化。
4 结论与展望
本文对近年来植被根系对土体水力学特性影响的相关研究和数据进行了分析,总结了植被根系的作用机理,主要包括:植被蒸腾吸水;根系穿透、根系扩张、孔隙填充和集粒包络等根系物理作用;根系腐烂、根系分泌物等导致土体有机质含量变化的根系生物化学作用。通过统计已有试验数据发现直根型根系易通过根系穿透、扩张产生更多的大孔隙,且根系收缩或衰亡后产生较大的孔隙通道,这些都会导致θs、α和Ks增加。而须根型根系在不同土体类型和生长阶段有不同的作用。在粗粒土中,生长初期根系以孔隙填充和集粒包络作用为主,导致θs、α和Ks的降低;随着根系密度的增加,根系填充作用减弱,根系穿透、扩张导致的孔隙形成作用显著,导致了θs增加,Ks和α减小。而在细粒土中,细根扩大土体中已存在的中小孔,同时沿着已有孔隙生长产生连续通道,使得土体的θs和Ks增加。另外,对于渗透性,直根型根系的增强作用比须根型根系显著,且在细粒土中尤为明显。随着植物的生长,Ks在不同生长阶段表现出不同的变化趋势。由此可见,这种由植被根系引起的土体水力学特性复杂变化主要依赖于植被蒸腾、植被根系物理作用和植被根系生物化学作用三者的相对贡献,同时,受根系类型、土体类型及生长阶段等因素的影响。
目前為止,植被根系对土体水力学特性影响的相关研究和数据有限,致使难以得出植被根系影响的一般规律。同时,考虑到不同条件下根系发育不同,有必要在不同条件下开展全面的试验研究,以确定不同变量在改变土体水力学特性中的作用。因此,需要大量的试验研究以获取各种土体类型和植物类型的相关数据。其次,文中所探究的机理是从微观上理解植物对土体水力学特性的不同影响,其中一些作用可能是相互促进的,如植被蒸腾作用、孔隙填充和集粒包络作用,也可能是相互矛盾的,如根系穿透和孔隙填充以及根系分泌物的作用。因此,土体的水力学特性(SWRC、Ks和K)究竟表现出增加或减小以及各种作用对土体水力学特性改变的相对贡献目前仍不可知。定量植被蒸腾作用、根-土相互作用对于植被护坡以及水循环计算具有重要意义。另外,土体的水力学特性具有高度时间相关性。尽管目前部分研究探究了特定植物对土体水力学特性的影响,但是大多局限于某一生长年龄,并且是特定阶段的监测,对于某一种植物全生命周期的研究尚未有报道。因此,对植被土全生命周期的水力学特性监测仍需大量开展,这对建立植物水力作用数据库和能够精确反应植被作用的土体水力学模型是必要且亟需的。
参考文献
[1]SHI X Q, QIN T L, YAN D H, et al. A meta-analysis on effects of root development on soil hydraulic properties [J]. Geoderma,2021, 403:115363.
[2]GADI V K, TANG Y R, DAS A, et al. Spatial and temporal variation of hydraulic conductivity and vegetation growth in green infrastructures using infiltrometer and visual technique[J].Catena,2017, 155: 20-29.
[3]SCHWEN A, BODNER G, SCHOLL P, et al. Temporal dynamics of soil hydraulic properties and the water-conducting porosity under different tillage[J].Soil and Tillage Research,2011, 113(2): 89-98.
[4]WANG X H, MA C, WANG Y Q, et al. Effect of root architecture on rainfall threshold for slope stability: Variabilities in saturated hydraulic conductivity and strength of root-soil composite[J].Landslides,2020, 17(8): 1965-1977.
[5]LUO W Z, LI J H, SONG L, et al. Effects of vegetation on the hydraulic properties of soil covers: Four-years field experiments in Southern China[J].Rhizosphere,2020, 16:100272.
[6]RAHARDJO H, SATYANAGA A, LEONG E C, et al. Performance of an instrumented slope covered with shrubs and deep-rooted grass[J].Soils and Foundations,2014, 54(3): 417-425.
[7]NG C W W, GUO H W, NI J J, et al. Long-term field performance of non-vegetated and vegetated three-layer landfill cover systems using construction waste without geomembrane[J/OL].Géotechnique(Ahead of Print).(2022-06-09)[2023-09-10].https:∥www.icevirtuallibrary.com/doi/10.1680/jgeot.21.00238.
[8]NI J J, LEUNG A K, NG C W W. Unsaturated hydraulic properties of vegetated soil under single and mixed planting conditions[J].Géotechnique,2019, 69(6): 554-559.
[9]SCHOLL P, LEITNER D, KAMMERER G, et al. Root induced changes of effective 1D hydraulic properties in a soil column[J].Plant and Soil,2014, 381(1): 193-213.
[10]BACQ-LABREUIL A, CRAWFORD J, MOONEY S J, et al. Effects of cropping systems upon the three-dimensional architecture of soil systems are modulated by texture[J].Geoderma,2018, 332: 73-83.
[11]YANG F, ZHANG G L, YANG J L, et al. Organic matter controls of soil water retention in an alpine grassland and its significance for hydrological processes[J].Journal of Hydrology,2014, 519: 3086-3093.
[12]AHMED M A, KROENER E, HOLZ M, et al. Mucilage exudation facilitates root water uptake in dry soils[J].Functional Plant Biology: FPB, 2014, 41(11): 1129-1137.
[13]LEUNG A K, BOLDRIN D, LIANG T, et al. Plant age effects on soil infiltration rate during early plant establishment[J].Géotechnique, 2018,68(7):646-652.
[14]VAN GENUCHTEN M T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils[J].Soil Science Society of America Journal,1980, 44(5): 892-898.
[15]SCHLESINGER W H, JASECHKO S. Transpiration in the global water cycle[J].Agricultural and Forest Meteorology,2014, 189/190: 115-117.
[16]TRENBERTH K E, SMITH L, QIAN T T, et al. Estimates of the global water budget and its annual cycle using observational and model data[J].Journal of Hydrometeorology,2007, 8(4): 758-769.
[17]OKI T, KANAE S. Global hydrological cycles and world water resources[J].Science, 2006, 313(5790): 1068-1072.
[18]XU K, GUO L, YE H. A naturally optimized mass transfer process: The stomatal transpiration of plant leaves[J].Journal of Plant Physiology, 2019, 234/235: 138-144.
[19]LI Z K, LI X Y, ZHOU S, et al. A comprehensive review on coupled processes and mechanisms of soil-vegetation-hydrology, and recent research advances[J].Science China Earth Sciences,2022, 65(11): 2083-2114.
[20]GARG A. Effects of vegetation types and characteristics on induced soil suction[D].Hong Kong: Hong Kong University of Science and Technology,2015.
[21]NG C W W, TASNIM R, WONG J T F. Coupled effects of atmospheric CO2 concentration and nutrients on plant-induced soil suction[J].Plant and Soil,2019, 439(1): 393-404.
[22]LIU Q, SU L J, XIA Z Y, et al. Effects of soil properties and illumination intensities on matric suction of vegetated soil[J].Sustainability,2019, 11(22):6475.
[23]LEUNG A K, GARG A, NG C W W. Effects of plant roots on soil-water retention and induced suction in vegetated soil[J].Engineering Geology,2015, 193: 183-197.
[24]NI J J, LEUNG A K, NG C W W, et al. Investigation of plant growth and transpiration-induced matric suction under mixed grass-tree conditions[J].Canadian Geotechnical Journal,2017, 54(4): 561-573.
[25]NG C W W, WOON K X, LEUNG A K, et al. Experimental investigation of induced suction distribution in a grass-covered soil[J].Ecological Engineering, 2013, 52: 219-223.
[26]FEDDES R, KOWALIK P, ZARADNY H. Simulation of field water use and crop yield[J].Journal of Plant Nutrition and Soil Science, 1980, 143(2): 254-255.
[27]GARG A, COO J L, NG C W W. Field study on influence of root characteristics on soil suction distribution in slopes vegetated with Cynodon dactylon and Schefflera heptaphylla[J].Earth Surface Processes and Landforms,2015,40(12): 1631-1643.
[28]CARMINATI A, VETTERLEIN D, KOEBERNICK N, et al. Do roots mind the gap?[J].Plant and Soil,2013, 367(1):651-661.
[29]LU J R, ZHANG Q, WERNER A D, et al. Root-induced changes of soil hydraulic properties: A review [J]. Journal of Hydrology,2020, 589:125203.
[30]JOTISANKASA A, SIRIRATTANACHAT T. Effects of grass roots on soil-water retention curve and permeability function[J].Canadian Geotechnical Journal,2017,54(11): 1612-1622.
[31]MARCACCI K M, WARREN J M, PERFECT E, et al. Influence of living grass Roots and endophytic fungal hyphae on soil hydraulic properties[J].Rhizosphere,2022, 22:100510.
[32]SONG L, LI J H, ZHOU T, et al. Experimental study on unsaturated hydraulic properties of vegetated soil [J]. Ecological Engineering,2017, 103: 207-216.
[33]TISDALL J M, OADES J M. Organic matter and water-stable aggregates in soils[J].European Journal of Soil Science,1982,33(2): 141-163.
[34]AHMED M A, HOLZ M, WOCHE S K, et al. Effect of soil drying on mucilage exudation and its water repellency: A new method to collect mucilage[J].Journal of Plant Nutrition and Soil Science,2015, 178(6): 821-824.
[35]ZHANG W C, GAO W D, WHALLEY W R, et al. Physical properties of a sandy soil as affected by incubation with a synthetic root exudate: Strength, thermal and hydraulic conductivity, and evaporation[J].European Journal of Soil Science,2021, 72(2): 782-792.
[36]KROENER E, HOLZ M, ZAREBANADKOUKI M, et al. Effects of mucilage on rhizosphere hydraulic functions depend on soil particle size[J].Vadose Zone Journal,2018, 17(1): 1-11.
[37]DENG W N, HALLETT P D, JENG D S, et al. The effect of natural seed coatings of Capsella bursa-pastoris L.Medik.(shepherd′s purse) on soil-water retention, stability and hydraulic conductivity[J].Plant and Soil,2015, 387(1/2): 167-176.
[38]KROENER E, ZAREBANADKOUKI M, KAESTNER A, et al. Nonequilibrium water dynamics in the rhizosphere: How mucilage affects water flow in soils[J].Water Resources Research,2014, 50(8): 6479-6495.
[39]HAYAT F, ABDALLA M, MUNIR M U. Effect of chia seed mucilage on the rhizosphere hydraulic characteristics [J]. Sustainability,2021, 13(6):3303.
[40]CARMINATI A, ZAREBANADKOUKI M, KROENER E, et al. Biophysical rhizosphere processes affecting root water uptake[J].Annals of Botany,2016, 118(4): 561-571.
[41]BRAX M, BUCHMANN C, KENNGOTT K, et al. Influence of the physico-chemical properties of root mucilage and model substances on the microstructural stability of sand[J].Biogeochemistry,2020,147(1): 35-52.
[42]THULLNER M, SCHROTH M H, ZEYER J, et al. Modeling of a microbial growth experiment with bioclogging in a two-dimensional saturated porous media flow field[J].Journal of Contaminant Hydrology,2004,70(1/2): 37-62.
[43]DUNBABIN V M, MCDERMOTT S, BENGOUGH A G. Upscaling from rhizosphere to whole root system: Modelling the effects of phospholipid surfactants on water and nutrient uptake[J].Plant and Soil,2006, 283(1/2): 57-72.
[44]READ D B, BENGOUGH A G, GREGORY P J, et al. Plant roots release phospholipid surfactants that modify the physical and chemical properties of soil[J].New Phytologist,2003, 157(2): 315-326.
[45]WHALLEY W R, RISELEY B, LEEDS-HARRISON P B, et al. Structural differences between bulk and rhizosphere soil[J].European Journal of Soil Science,2005, 56(3): 353-360.
[46]南京水利科
