Effects of wind speed, underlying surface, and seed morphological traits on the secondary seed dispersal in the Tengger Desert, China
2024-04-22QUWenjieZHAOWenzhiYANGXinguoWANGLeiZHANGXueQUJianjun
QU Wenjie, ZHAO Wenzhi, YANG Xinguo, WANG Lei, ZHANG Xue, QU Jianjun*
1 Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China;
2 Key Laboratory of Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education,Ningxia University, Yinchuan 750021, China;
3 University of Chinese Academy of Sciences, Beijing 100049, China;
4 School of Ecology and Environment, Ningxia University, Yinchuan 750021, China
Abstract: The maintenance of sand-fixing vegetation is important for the stability of artificial sand-fixing systems in which seed dispersal plays a key role.Based on field wind tunnel experiments using 11 common plant species on the southeastern edge of the Tengger Desert, China, we studied the secondary seed dispersal in the fixed and semi-fixed sand dunes as well as in the mobile dunes in order to understand the limitations of vegetation regeneration and the maintenance of its stability.Our results indicated that there were significant variations among the selected 11 plant species in the threshold of wind speed (TWS).The TWS of Caragana korshinskii was the highest among the 11 plant species, whereas that of Echinops gmelinii was the lowest.Seed morphological traits and underlying surface could generally explain the TWS.During the secondary seed dispersal processes, the proportions of seeds that did not disperse (no dispersal) and only dispersed over short distance (short-distance dispersal within the wind tunnel test section) were significantly higher than those of seeds that were buried (including lost seeds) and dispersed over long distance (long-distance dispersal beyond the wind tunnel test section).Compared with other habitats, the mobile dunes were the most difficult places for secondary seed dispersal.Buried seeds were the easiest to be found in the semi-fixed sand dunes, whereas fixed sand dunes were the best sites for seeds that dispersed over long distance.The results of linear mixed models showed that after controlling the dispersal distance, smaller and rounder seeds dispersed farther.Shape index and wind speed were the two significant influencing factors on the burial of seeds.The explanatory power of wind speed, underlying surface, and seed morphological traits on the seeds that did not disperse and dispersed over short distance was far greater than that on the seeds that were buried and dispersed over long distance, implying that the processes and mechanisms of burial and long-distance dispersal are more complex.In summary, most seeds in the study area either did not move, were buried, or dispersed over short distance, promoting local vegetation regeneration.
Keywords: seed dispersal; seed morphological traits; wind speed; vegetation regeneration; wind tunnel; Tengger Desert
1 Introduction
Revegetation is an effective measure to prevent the expansion of desertification (Wang and Zhu,2001).Since the 1950s, bioengineering measures such as sand barriers and planting xerophytic shrubs have been carried out to combat desertification in China (Jiang et al., 2002).The introduction of xerophytic shrubs altered the distribution patterns of precipitation and atmospheric dust, while the accumulation of litter and the formation of cryptogam accelerated the process of soil formation (Li et al., 2018).A large amount of cryptogam has propagated, and annual and perennial herbs have colonized revegetated areas (Chen et al., 2017).However, with the drying of the deep soil in the fixed sand dunes, the dominant position of shrub species gradually weakened,and the vegetation structure and underlying surface conditions have also changed significantly(Yang et al., 2019).At present, maintaining the stability of revegetation and realizing natural regeneration and succession from artificial intervention vegetation systems to near-zonal vegetation has become a major issue in environmental protection and ecological management (Li et al., 2013, 2016).
Vegetation regeneration involves the entire life cycle of seed germination, seedling establishment, reproduction, and diaspore dispersal by mature plants; diaspore dispersal is a key process in vegetation regeneration, since it signifies the end of the reproductive cycle and the establishment of offspring (Wang and Smith, 2002; Huang et al., 2012; Han et al., 2021).Diaspores refer to the parts of the plant that are dispersed, including seeds, fruits, infructescence,and other dispersal units (Zhu and Liu, 2012).The propagules of the mother plant that fall to the surface are again dispersed through the wind; this is called secondary wind dispersal (Schurr et al., 2005).Diaspore dispersal includes four categories: no dispersal, short-distance dispersal,burial, and long-distance dispersal (Liang, 2019).These modalities will lead to differences in the distribution patterns of seedlings and plants (Poulsen et al., 2012; Liang et al., 2019).
Diaspores have evolved multiple morphological structures to facilitate the capacity for dispersal (Mamut et al., 2023).Balloon, hairy, and winged diaspores can promote dispersal, and awned diaspores are beneficial for both dispersal and burial (Liang, 2019; Cavanagh et al., 2020).Diaspores without appendages tend to resist long-distance dispersal, except for extremely light dust diaspores (Batty et al., 2006; Liang, 2019).In addition, other diaspore traits may also affect dispersal by wind (Muller-Landau et al., 2008).For example, diaspores with less mass or longer length are more likely to be dispersed over longer distance, and a discoidal shape has an advantage over a spherical shape in terms of interacting with wind currents and achieving a greater displacement (Cousens and Rawlinson, 2001; Thomson et al., 2011; Zhu et al., 2016;Wyse and Hulme, 2021; Guzmán-Vázquez et al., 2023).Studies have suggested that projection area, shape index, wing loading, and the ratio of projection area to volume of diaspores will affect dispersal ability (Dorp et al., 1996; Nathan and Muller-Landau, 2000; Aavik and Helm, 2018;Liang et al., 2019).However, there are some conflicting results.For example, some studies have indicated that seed mass has no impact on seed dispersal distance (e.g., Jenkins et al., 2007), in contrast to previous research (Greene and Johnson, 1993; Zhu et al., 2016).It is unclear whether other morphological traits that have not been considered in previous studies would alter the importance of the originally considered diaspore attributes.
The interaction between wind speed and habitat has an impact on diaspore dispersal (Higgins et al., 2003).It has been suggested that gentle winds produce short-distance dispersal, while strong winds may induce long-distance dispersal (Nathan et al., 2002).However, some studies have found the opposite pattern (e.g., Tackenberg, 2003).Different habitats can produce different airflow patterns.Thermal updrafts are dominant in open landscapes (inhibiting long-distance dispersal) (Tackenberg, 2003), while shear-induced updrafts are dominant in forests (facilitating long- distance dispersal) (Nathan et al., 2002; Higgins et al., 2003).Underlying surface, plant height, and slope may be also important for wind dispersal (Higgins et al., 2003; Thomson et al.,2011; Liang, 2019).Heterogeneity of the habitat is widespread in nature, and thus the interaction among the diaspore attributes, wind speed, and habitat can be quite complex.
Field wind tunnel experiments are more conducive to capturing the patterns of secondary seed dispersal under various habitat characteristics (Thompson and Katul, 2013; Teller et al., 2016;Zhou et al., 2019; Zhou et al., 2020).However, due to complex equipment operation and high costs, this approach is rarely applied in ecological research.In general, although secondary seed dispersal is an important process affecting vegetation regeneration, field empirical research still lags behind theoretical research due to the limitations of study systems and methods (Liang et al.,2019).
The research on secondary seed dispersal started relatively late in arid areas.For example,present studies in the Tengger Desert, China, have focused on soil seed banks, the germination and settlement of short-lived plants, and the relationships between plants and biological crusts (Li et al., 2014, 2018; Liu et al., 2014; Liu et al., 2018; Song et al., 2019; Wang et al., 2019).As a result, there is a paucity of research on secondary seed dispersal.Taking the Tengger Desert as an example, apart from external factors such as soil moisture, can secondary seed dispersal mechanisms partly explain the present lack of progress in vegetation regeneration? Understanding the potential for vegetation regeneration via sand fixation in the Tengger Desert is a key point.Therefore, the main purposes of this study were to determine (1) how seed morphological traits and underlying surface characteristics affect the threshold of wind speed (TWS) of diaspores; (2)how seed morphological traits, underlying surface characteristics, and wind speed interact to regulate the destinations of seeds; and (3) the relationship between secondary seed dispersal and vegetation regeneration based on the characteristics of secondary seed dispersal and its influencing factors in the Tengger Desert.The overall goal is to deeply understand the limiting factors for community regeneration and stability of sand-fixing vegetation, and the results of this study hold significant importance for guiding the sustainable management of artificial sand-fixing vegetation in arid areas.
2 Materials and methods
2.1 Study area
The study area is located in the transitional zone from desert to oasis on the southeastern edge of the Tengger Desert in Ningxia Hui Autonomous Region, China (37°25′58′′-37°37′24′′N,104°49′25′′-105°09′24′′E; 1339 m; Fig.1).The mean annual temperature of the study area is 10.6°C, the mean annual potential evaporation is 3000 mm, and the mean annual precipitation is only 186 mm.Approximately 80% of the total annual precipitation occurs between May and September.The annual mean wind speed is 2.90 m/s; the wind frequency in the range of 5.00-11.00 m/s is 98.48%, and the main windy season is from March to July (Qu et al., 2007; Li et al., 2014).
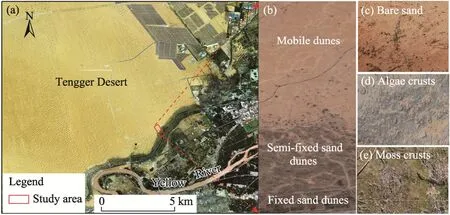
Fig.1 Location of the study area in the Tengger Desert (a) and photos showing its spatial patterns (b-e).The images were obtained from the Beijing No.2 Satellite in July 2021.
In 1956, a vegetation system combining xerophytic shrubs and straw checkerboard barriers was established in the mobile sand dunes to protect the Shapotou section of Baotou-Lanzhou Railway against wind erosion and sand burial hazards.With the gradual formation of a sand-fixing zone,the surface gradually became stable.The spatial distribution pattern of revegetation in different(construction) succession stages formed an artificial sand-fixing vegetation system perpendicular to the railway.
2.2 Design of the field experimental plots
According to the distance from the artificial vegetation area of railway, the degree of dune stabilization, the type of crust coverage, and the composition of dominant vegetation, we divided the study area into three types of habitats, namely fixed sand dunes (60%-85% moss crusts),semi-fixed sand dunes (60%-70% algae crusts), and mobile dunes (bare sand without crusts) (Fig.1).The basic characteristics of each habitat are shown in Table 1.We selected three experimental plots (20 m×20 m for each) from these three habitats for field wind tunnel experiments conducted in June 2022.
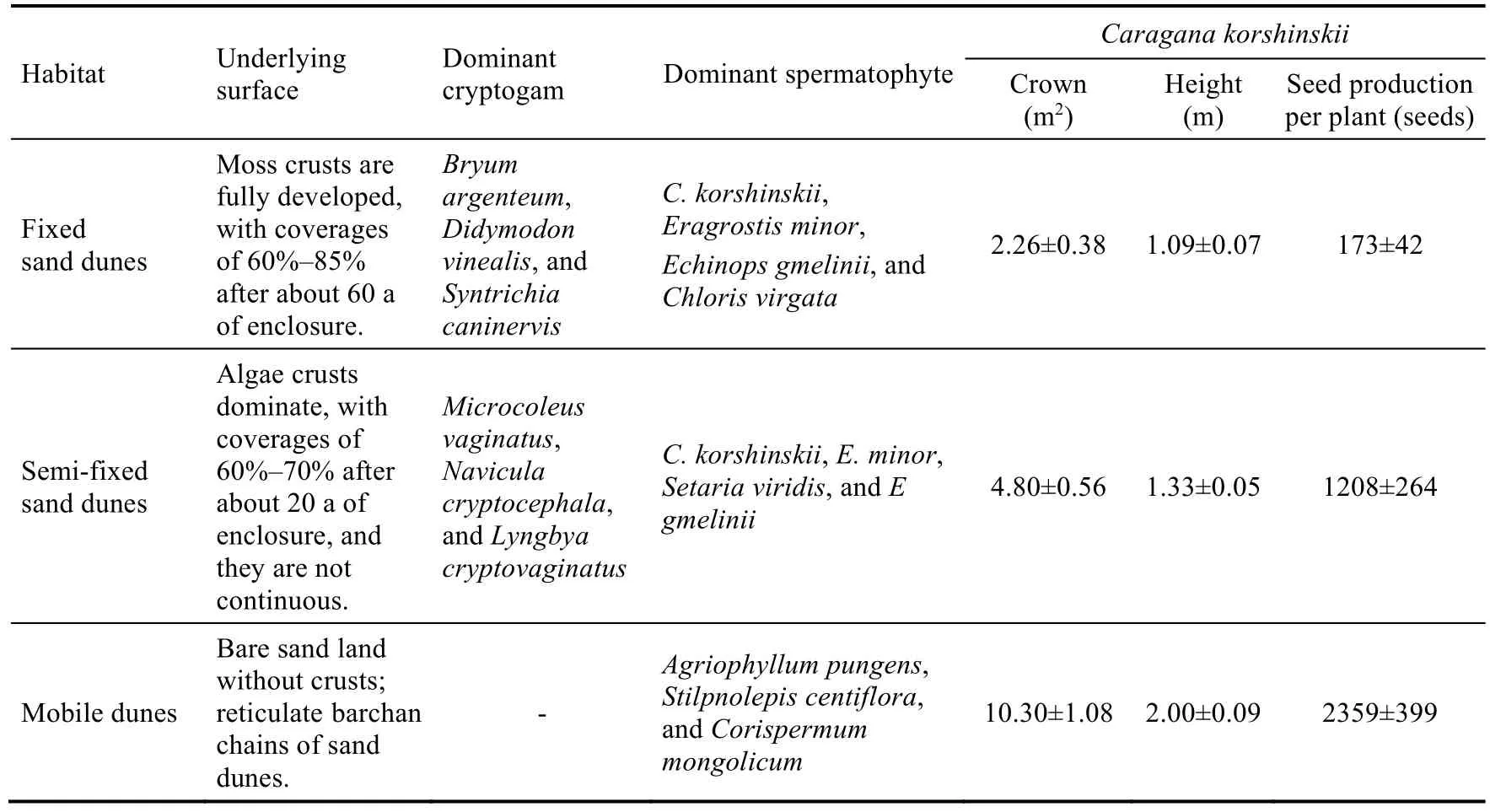
Table 1 Basic characteristics of the three habitats
2.3 Seed collection and measuring methods of seed morphological traits
We selected 11 common plant species based on their dominance, seed size, and suitability for field wind tunnel experiments:Caragana korshinskii(shrub),Corethrodendron scoparium(shrub),Ammopiptanthus mongolicus(shrub),Corispermum mongolicum(annual herb),Agropyron cristatum(perennial herb),Allium mongolicum(perennial herb),Hippophae rhamnoides(shrub),Echinops gmelinii(annual herb),Nitraria tangutorum(shrub),Corethrodendron fruticosum(shrub), andCalligonum mongolicum(shrub).These selected plant species represented the main dominant species in the study area, excluding those with seeds too small to be conducive to wind tunnel tests and data analysis.Additionally, we also selected some common plant species with seeds that significantly differ from those of the dominant species,which enhanced the applicability of wind tunnel experiments.For shrub species, 20-30 plants were selected from each species, and for herb species, 30-50 plants were selected from each species, ensuring the collection of 3000-4000 seeds for each plant species in the study area, and then the collected seeds were dried in the shade.The characteristics of each plant species and seed morphological traits are listed in Tables S1 and S2 and Figure S1.In April 2022, the more evenly sized and plump seeds were selected as the experimental materials.To facilitate the observations and the search for seeds during the field wind tunnel experiments, we lightly sprayed the surfaces of seeds without appendages using aerosol coatings of different colors.The spraying color did not change the quality or morphology of seeds; the colors were used only for identification and did not affect the experimental results.During the painting process, the seeds were separated from each other and dried and preserved naturally.
The seed morphological traits (length, width, thousand seed weight (TSW), projection area, and wing loading) are important indices used to measure secondary seed dispersal by wind.Fifty seeds with intact appearance were selected for each plant species and then painted.The morphological traits of the collected seeds were measured using a Plant Seed Analyzer (OPTO-Agri,Optomachines, Riom, France), namely length, width, TSW, projection area, perimeter, diameter,ratio of length to width, circularity, compacity, and rugosity.The thickness (height) of the collected seeds was measured with Vernier calipers (0.01 mm accuracy).We obtained wing loading and shape index using the following equations (Matlack, 1987; Thompson et al., 1993):
whereωis the wing loading (g/mm2); TSW is the thousand seed weight (g);Sis the projection area (mm2);Vsis the shape index;nis the total dimensions of seed (n=3); andxiis theithdimension of seed (x1,x2andx3in this study).It is noted thatx1was calculated as the ratio of seed length to seed length,x2as the ratio of seed width to seed length, andx3as the seed height to seed length.
2.4 Field wind tunnel experiments
The portable field wind tunnel experiments can simulate the physical relationship between horizontal wind and underlying surface to examine the secondary seed dispersal process by wind(Liu et al., 2015).The field wind tunnel equipment was developed by the Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, China (Fig.2).The wind tunnel was 0.6 m high and 0.6 m wide with a 6.0 m long test section, consisting of six movable segments(one segment size: 1.0 m×0.6 m×0.6 m).A transformer was used to change the wind speed,meanwhile a Pitot tube (160-60, Dwyer Instruments Inc., Michigan City, Indiana, USA) was applied to monitor the wind speed in the field wind tunnel in real time.The wind speed in the field wind tunnel test section had a positive linear correlation with the frequency of the transformer motor.By adjusting the frequency of the transformer motor, the wind speed could be increased or decreased.The transformer motor could produce stable wind speeds in the range of 0.00-15.00 m/s.The Pitot tube was 5.5 m away from the input end, and the wind speed was measured at 0.3 m above the surface.The other end of the Pitot tube was connected to a laptop for real-time wind speed monitoring.
Three habitats, namely, fixed sand dunes (60%-85% moss crusts), semi-fixed sand dunes(60%-70% algae crusts), and mobile dunes (bare sandy land), in the sand-fixating vegetation system along the Shapotou section of Baotou-Lanzhou Railway on the southeastern edge of the Tengger Desert were selected for field wind tunnel experiments (Table 1).For the field wind tunnel experiments, we selected a 20 m×20 m sample plot with flat area in each habitat.The wind tunnel equipment was placed in an area with relatively flat terrain without vegetation and perpendicular to the environmental wind direction to prevent the wind from blowing into the wind tunnel test section.The connection of the wind tunnel test section was sealed with tape, and the area where the wind tunnel equipment is in contact with the ground was sealed with sand to prevent air leakage.After each set of wind tunnel tests in the fixed and semi-fixed sand dunes, the wind tunnel test section was moved away from the experimental area, and the secondary seed dispersal was measured.The surface in the test section was not trampled during the test process to ensure that the underlying surfaces of moss and algae crusts were in good conditions.For the mobile dunes, the sand surface needed to be restored and leveled before the next set of wind tunnel tests.

Fig.2 Photo of field wind tunnel equipment (a) and diagram of wind tunnel test section (b)
2.4.1 Measurement of TWS
The drag or shear force exerted by the wind for seed initial motion is called the threshold shear stress, whereas the corresponding wind speed is called TWS for critical wind speed (Zhou et al.,2020).In this study, we measured the TWS of the selected 11 plant species.Ten seeds of each plant species were randomly released each time from 10.0 cm above the surface to ensure the randomness of the initial orientation of seeds.The transformer motor frequency started from zero and increased by 0.01 Hz in each time step, and the wind speed shown by the computer (i.e., TWS)was recorded when two or more seeds were displaced (Liang et al., 2020).The experiment comprised five replicates for each plant species, with a total of 1650 seeds (11 plant species×3 underlying surfaces×10 seeds×5 replicates).
2.4.2 Measurement of secondary seed dispersal
According to the results of K-means cluster algorithm (a classic algorithm of partition clustering)(Sinaga et al., 2021) of seed morphological traits from 11 plant species, we selectedC.scoparium,C.korshinskii,E.gmelinii,C.fruticosum,andCalligonum mongolicumto carry out the wind tunnel experiments of secondary seed dispersal.To make more seeds spread in the test section, we set the wind speed using different multiples of TWS.According to the TWS of seeds and the maximum wind speed that could be stably controlled by the pressure transformer (15.00 m/s), we designed the experimental wind speed gradients as 1.0, 1.2, and 1.4 times the TWS.When the transformer reached a stable wind speed, it was held for 2 min and then stopped.An interceptor net was set at the air outlet to collect seeds.Ten seeds of each plant species were selected for each set of wind tunnel tests, and each test was repeated three times.Therefore, a total of 1350 seeds were released (5 plant species×3 underlying surfaces×3 wind speed gradients×10 seeds×3 replicates).
After each set of wind tunnel tests, we investigated the secondary seed dispersal.There were four possibilities for the final destinations of seeds: no dispersal, short-distance dispersal (within the test section), burial (including lost seeds), and long-distance dispersal (beyond the test section).We adopted a distance of 6.0 m to distinguish short-distance dispersal and long-distance dispersal by considering the parameters of our wind tunnel equipment and referring to the studies of Venable et al.(2008), Liang (2019), and Qin (2021).The numbers of seeds that dispersed over long distance and did not no disperse were counted initially, and the number and distance of seeds that dispersed over short distance were recorded; finally, the number of lost seeds was calculated.In the mobile dunes, the seeds that were completely buried in the sand were counted as buried seeds (burial), whereas those were not completely buried in the sand were counted as short-distance dispersal seeds.
2.5 Data analysis
The 11 plant species were grouped by the K-means clustering algorithm using the seed morphological traits.One-way analysis of variance (ANOVA) was used to compare and analyze seed morphological traits and TWS for different underlying surfaces of the 11 plant species.To address the non-independence of data and avoid pseudoreplication in the experiments, we used linear mixed models to evaluate the individual effects of seed morphological traits and underlying surfaces on TWS and the interaction effect of seed morphological traits and underlying surfaces on secondary seed dispersal (no dispersal, short-distance dispersal, burial, and long-distance dispersal) at different wind speeds using the R package "lme4" (Bates et al., 2015).We used plant species as random effect in the linear mixed models, and all seed morphological traits, underlying surface characteristics, and wind speed as fixed effects.First, a collinearity test was performed on the explanatory variables, and variables with variance inflation factor>5 were removed.The linear mixed model analyses identified significant variables and calculated the marginalR2(whereR2represents the variance of the fixed effects) and conditionalR2values (models 1-5; Table 2).To calculate the variance explained uniquely by each significant predictor, we refitted the linear mixed models based on the preliminary significant variables in the R package 'partR2' (models 6-10; Table 2) (Stoffel et al., 2021).Across the analyses, omnibus tests were conducted with the''ANVOA'' function in the R package 'car' (Fox and Weisberg, 2019).The other data were analyzed and mapped with R software (R Core Team, 2023).
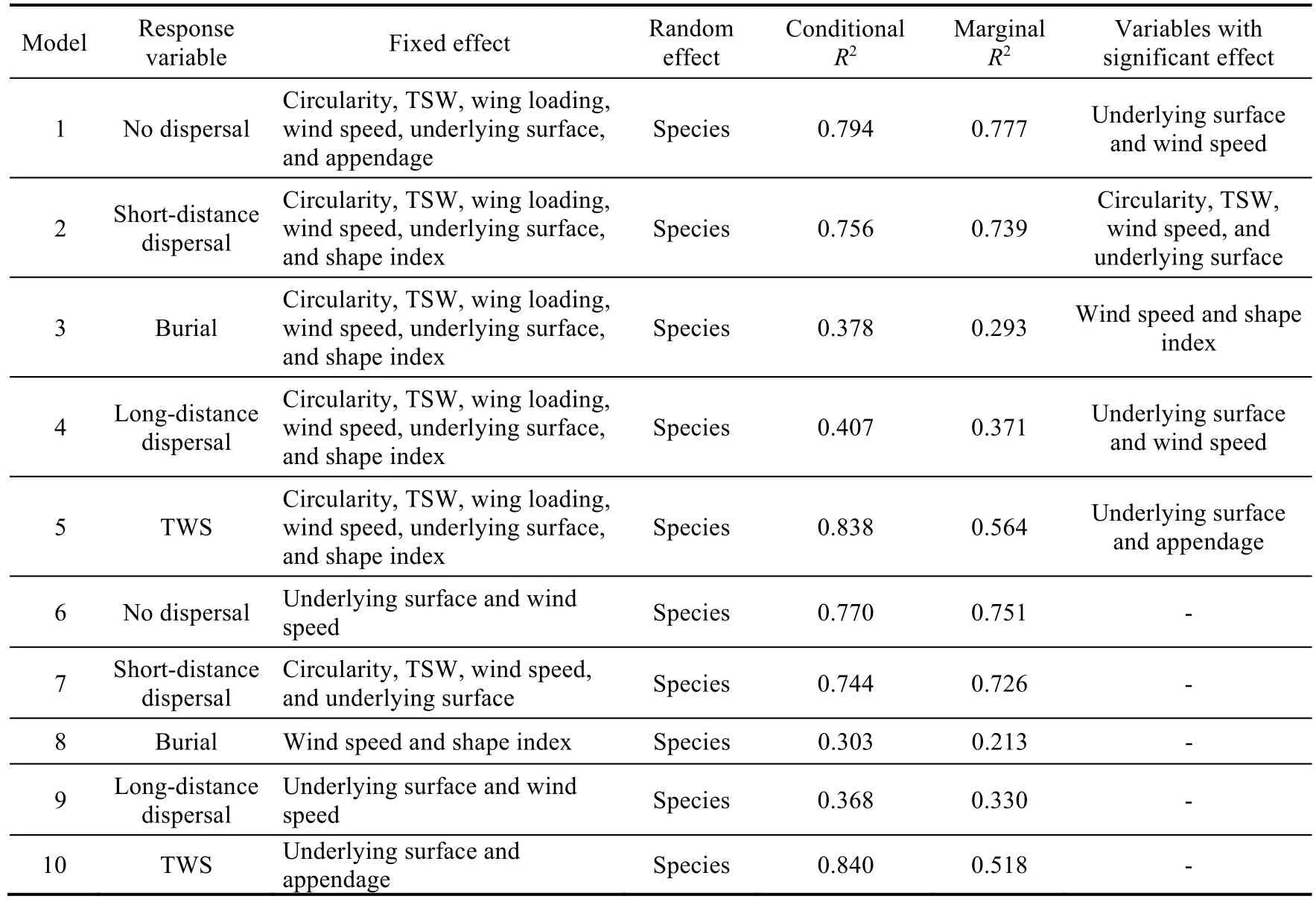
Table 2 Linear mixed models implemented to assess the effects driving TWS and the destination of seeds
3 Results
3.1 Seed morphological traits and cluster analysis
The seed morphological traits of the 11 selected plant species in the study area were determined using the Plant Seed Analyzer (Tables S1 and S2).Overall, there were clear differences in seed morphological traits; for example, the differences in the life forms of plant species such as shrubs and herbs were highly significant.
Based on the results of the K-means cluster analysis shown in Figure 3, we found that Cluster I was composed ofCorispermum mongolicum,H.rhamnoides,A.cristatum,A.mongolicum,andE.gmelinii, among whichH.rhamnoidesis a shrub species, while others are herbs.The width of seeds varied from 1.07 to 2.39 mm; the perimeter varied from 9.29 to 16.27 mm; the projection area ranged from 3.37 to 6.32 mm2; the diameter varied from 2.07 to 2.83 mm; and the TSW ranged from 1.38 to 4.20 g.These indices were significantly lower than those of the other three clusters that were dominated by shrubs.Cluster IV only hadCalligonum mongolicum(width of 6.85 mm, projection area of 41.90 mm2, and perimeter of 45.93 mm).The diameter (7.28 mm) ofCalligonum mongolicumwas the largest among the 11 plant species, and its TSW (64.96 g) was also the highest.Cluster III (includingC.scoparium,C.korshinskii,andA.mongolicus) and Cluster II (includingN.tangutorumandC.fruticosum) were also composed of shrubs, and the values of their seed morphological traits were moderate.
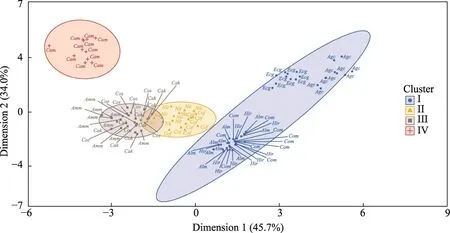
Fig.3 Diagram showing the K-means cluster analysis of plant species based on seed morphological traits.Ecg,Echinops gmelinii; Cam, Calligonum mongolicum; Cos, Corethrodendron scoparium; Cof, Corethrodendron fruticosum; Nit, Nitraria tangutorum; Agc, Agropyron cristatum; Hir, Hippophae rhamnoides; Com, Corispermum mongolicum; Cak, Caragana korshinskii; Alm, Allium mongolicum; Amm, Ammopiptanthus mongolicus.The abbreviations are the same in the following figures.
3.2 TWS
The TWS values of the 11 plant species in descending order were as follows:A.mongolicus>C.korshinskii>A.mongolicum>N.tangutorum>H.rhamnoides>Corispermum mongolicum>A.cristatum>C.fruticosum>C.scoparium>Calligonum mongolicum>E.gmelinii(Fig.4).We divided these 11 plant species into five groups according to their TWS values.Specifically, the first group consisting ofA.mongolicushad the highest TWS (11.45 m/s).The second group includedC.korshinskiiandA.mongolicum, and the TWS values were 8.22 and 9.45 m/s,respectively.The third group consisted ofN.tangutorum,H.rhamnoides, andCorispermum mongolicum, and their TWS values were 7.94, 7.51, and 7.49 m/s, respectively.The fourth group includedA.cristatum,C.fruticosum, andC.scoparium, with the TWS values of 5.59, 6.04, and 6.37 m/s, respectively.The fifth group includedCalligonum mongolicumandE.gmelinii, which had the lowest TWS values, i.e., 3.09 and 2.40 m/s, respectively.
The TWS of the 11 plant species differed among the habitats.In the fixed sand dunes with moss crusts, the maximum TWS was associated withA.mongolicus, ranging from 10.26 to 12.63 m/s,with an average of 11.14 m/s.The TWS values ofCalligonum mongolicumandE.gmeliniiwere the lowest, in the ranges of 1.80-2.62 and 2.98-3.42 m/s, respectively.In the habitats of semi-fixed sand dunes with algae crusts and fixed sand dunes with moss crusts, the plant species with the lowest TWS were the same, namelyCalligonum mongolicumandE.gmelinii.For these two plant species, the changes in their TWS values were less than that ofA.mongolicus, which had the highest TWS value.In the mobile dunes, the plant species with the highest TWS wereA.mongolicusandC.korshinskii, with the ranges of 8.83-9.56 and 8.38-9.29 m/s, respectively, and the average values of 9.11 and 8.77 m/s, respectively.The TWS values ofCalligonum mongolicumandE.gmeliniiwere the lowest, ranging from 1.27 to 2.29 and 1.80 to 4.12 m/s,respectively (averages of 1.95 and 3.12 m/s, respectively).

Fig.4 Comparison of TWS of the 11 plant species in different habitats.(a), fixed sand dunes with moss crusts;(b), semi-fixed sand dunes with algae crusts; (c), mobile dunes.TWS, threshold of wind speed.Bars represent standard errors.Different lowercase letters within the same habitat indicate significant differences in TWS among different plant species at the P<0.05 level.
3.3 Secondary seed dispersal process
As shown in Figure 5, the proportion of seeds that did not disperse was the highest in the mobile dunes (proportion of 74.8%), followed by the semi-fixed sand dunes with algae crusts (52.2%)and fixed sand dunes with moss crusts (48.2%).The proportion of seeds that dispersed over short distance was the highest in the fixed sand dunes with moss crusts (40.2%), followed by the semi-fixed sand dunes with algae crusts (39.3%).There was no significant difference in the proportion of seeds that dispersed over short distance between the two habitats, but the proportions were significantly higher than that in the mobile dunes, indicating that the difficulty of secondary seed dispersal was significantly increased in the mobile dunes compared with the fixed and semi-fixed sand dunes.The order of the proportions of seeds that were buried in the three habitats was as follows: semi-fixed sand dunes with algae crusts (12.0%)>mobile dunes(7.1%)>fixed sand dunes with moss crusts (4.9%).The proportion of seeds that dispersed over long distance was the highest in the fixed sand dunes with moss crusts, followed by the mobile dunes.
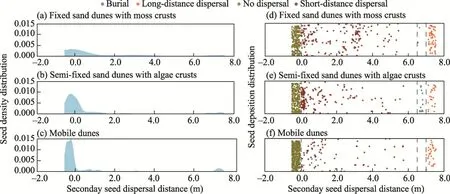
Fig.5 Distributions of seed density and seed deposition during the secondary seed dispersal processes in different habitats.(a and d), fixed sand dunes with moss crusts; (b and e), semi-fixed sand dunes with algae crusts;(c and f), mobile dunes.The dots to the left of the gray dashed line at the 0.0 scale value represent the seeds that did not disperse.The dots between the two gray dashed lines at the 6.5 and 7.0 scale values represent the lost seeds.The dots to the right of the gray dashed line at the 7.0 scale value represent the seeds that dispersed over long distance.
Figure 6 shows the distributions of secondary seed dispersal by wind for different major plant species in the three habitats.In the fixed sand dunes with moss crusts, the seeds ofC.korshinskii,E.gmelinii,C.scoparium,Calligonum mongolicum, andC.fruticosumwere dispersed more evenly.In the mobile dunes, the secondary seed dispersal distances ofC.scopariumandC.fruticosumwere less than those of the other plant species.In the three habitats, the secondary seed dispersal distance ofCalligonum mongolicumwas the longest.There was no significant difference in the proportions of seeds that did not disperse among different plant species in the three habitats.In the fixed sand dunes with moss crusts, the highest proportion of seeds that were buried wasC.fruticosum(2.2%), while in the semi-fixed sand dunes with algae crusts and mobile dunes, the highest proportion of seeds that were buried wasC.scoparium(5.6% and 7.7%,respectively).Compared with other plant species,the seeds ofCalligonum mongolicumandE.gmeliniitended to spread farther away.
3.4 Drivers of TWS
Linear mixed model 5 showed that TWS could be modeled as a function of circularity, TSW, wing loading, underlying surface, appendage, and shape index, in addition to the random effect (Table 2).The marginalR2and conditionalR2values were 0.564 and 0.838 in this model, respectively.Species (random effect) explained 27.4% of the total variance in TWS.Both appendage and underlying surface were significant factors driving TWS (P<0.05).Appendage explained 42.6%of the total variance in TWS, while underlying surface only explained 9.2% (Fig.7).The TWS of seeds with conductive appendages was lower than that of seeds without conductive appendages.
3.5 Drivers of secondary seed dispersal pattern
Linear mixed models 1-4 showed that secondary seed dispersal could be modeled as a function of circularity, TSW, wing loading, underlying surface, wind speed, and shape index, in addition to the random effect (Table 2).No dispersal and long-distance dispersal were strongly correlated with wind speed and underlying surface; short-distance dispersal was related with wind speed,underlying surface, and seed morphological traits (circularity and TSW); and burial was correlated with shape index and wind speed.The most variance in the fixed effects of no dispersal, short-distance dispersal, burial, and long-distance dispersal patterns can be explained by wind speed (Fig.8).In addition, the explanatory power of wind speed, underlying surface, and seed morphological traits on the seeds that did not disperse and dispersed over short distance was far greater than that on the seeds that were buried and dispersed over long distance.
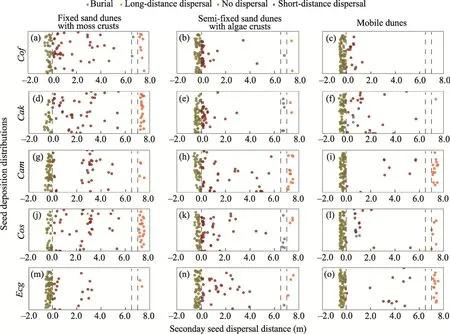
Fig.6 Secondary seed dispersal patterns of major plant species in the fixed sand dunes with moss crusts,semi-fixed sand dunes with algae crusts, and mobile dunes.(a-c), Cof; (d-f), Cak; (g-i), Cam; (j-l), Cos; (m-o),Ecg.The dots to the left of the gray dashed line at the 0.0 scale value represent the seeds that did not disperse.The dots between the two gray dashed lines between the 6.5 and 7.0 scale values represent the lost seeds.The dots to the right of the gray dashed line at the 7.0 scale value represent the seeds that dispersed over long distance.
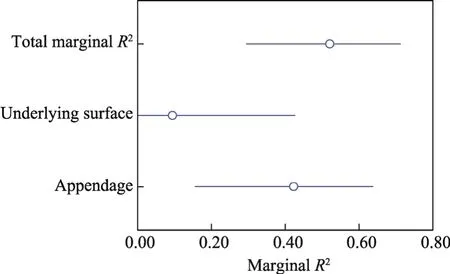
Fig.7 Variance partition of the significant drivers for TWS.R2, variance of the fixed effects.The circles represent the marginal R2 value for each driver and the total marginal R2 value.The black lines represent the 95%confidence intervals of the marginal R2 values.
4 Discussion
4.1 Seed morphological traits, underlying surface configuration, and TWS

Fig.8 Variance partition of the significant drivers for the secondary seed dispersal.The circles represent the marginal R2 value for each driver and the total marginal R2 value.The black lines represent the 95% confidence intervals of the marginal R2 values.
Wind-driven dispersal is the primary factor in secondary seed dispersal in desert areas.TWS refers to the minimum wind speed at which the diaspore moves.Different underlying surface conditions such as near-surface turbulence changes caused by land cover and micro-topography as well as seed morphological traits including shape, mass, projection area, wing loading, and appendage may all be the key factors affecting TWS (Green, 1980; Andersen, 1993; Nathan and Muller-Landau, 2000; Horn et al., 2001; Lemke et al., 2009; Liang et al., 2019).Among the 11 studied plant species,C.korshinskiihad the highest TWS (Fig.4), indicating that its dispersal ability was relatively poor, a result that was related to its high resistance and low susceptibility to wind force caused by its flat shape and smooth surface.The seeds ofE.gmeliniihad feathery appendages that make the seeds easier to move, and this species had the lowest TWS (Fig.4).Feathery or bristle appendage is a much stronger predictor of TWS for seeds (Liang, 2019).It has been suggested that the pappus of Asteraceae contributes to secondary seed dispersal and increases the probability of long-distance dispersal (Mandel et al., 2019).The bristle appendages could raise the height of the seed from the ground and increase the area of the seed in contact with the wind, thus promoting horizontal movement (Liang et al., 2019).This could explain the results of the lower TWS values ofE.gmelinii(feathery appendages) andCalligonum mongolicum(bristle appendages).
Underlying surface also had a significant influence on the TWS of seeds, but its explanatory power was lower than that of appendage.The effect of underlying surface was reflected in the different absolute values of TWS.The TWS values of the same plant species in the semi-fixed sand dunes with algae crusts were relatively higher than those in the fixed sand dunes with moss crusts and mobile dunes, partly because the underlying surface of this habitat was more likely to form cracks or micro-pits and other micro-surfaces that restricted seed movement (He et al., 2023).
The interaction between seed morphological traits and underlying surface often determines the variation and adaptation of the TWS of seeds.Most common plant seeds with feathery, bristle,awn, and hard spiny appendages, or light seeds in the fixed sand dunes with moss crusts and semi-fixed sand dunes with algae crusts might facilitate the dispersal of seeds to new locations(Cavanagh et al., 2020; Zhou et al., 2020).Most plant seeds without appendages in the mobile dunes have higher TWS values, such asCorispermum mongolicum, which could reduce TWS by taking advantage of the terrain (Liang, 2019).
4.2 Secondary seed dispersal pattern
In general, there are four patterns of secondary seed dispersal: no dispersal, short-distance dispersal, burial, and long-distance dispersal.The final pattern of secondary seed dispersal (the proportional relationship of the four probabilities) will be determined by the combined effect of wind speed, underlying surface, and seed morphological traits (Chambers and MacMahon, 1994;Nathan and Muller-Landau, 2000).
In this study, no dispersal and short-distance dispersal were the main patterns of secondary seed dispersal.The probability of seeds that dispersed over long distance was very low, which is consistent with the results of other simulated wind tunnel experiments (Liang et al., 2019; Zhou et al., 2020).Horizontal wind is an important factor driving secondary seed dispersal; the stronger the wind, the easier the seed dispersal, and the longer the dispersal distance (Tackenberg, 2003).Underlying surface could usually change the wind speed by interacting with the near-surface wind speed, and its surface microstructure also affects the ability of seeds to roll and move, thus influencing the dispersal distance (Field and Pelletier, 2018).During high winds, the seeds roll or jump faster, and underlying surface has less impact on the seeds, allowing them to disperse farther.Therefore, the interaction between wind speed and underlying surface determines the secondary seed dispersal.
Short-distance dispersal was also correlated with the circularity and TSW of seeds.Circularity refers to the degree of roundness.When it is close to 1, the seed is rounder and can spread farther(Liang, 2019).In addition, the larger the seed mass, the shorter the dispersal distance.However,these two drivers (circularity and TSW) had a significant but low contribution to short-distance dispersal.Some previous studies indicated that the relationship between seed mass and dispersal distance may be scale-dependent: at a small scale, seed mass is negatively correlated with dispersal distance, but with the increase of scale, this correlation does not exist (Muller-Landau et al., 2008; Thomson et al., 2011).Furthermore, species with small seeds spread more widely than species with large seeds, thereby increasing the likelihood of survival (Hyatt et al., 2003).In general, small and round seeds are more easily dispersed.
Long-distance dispersal is vital for seed dispersal and population regeneration, and existing models have verified that updrafts could significantly increase the distance of secondary seed dispersal (Maurer et al., 2013).Long-distance dispersal primarily depends on the high wind speed, possibly because the strong horizontal wind could generate upward vertical wind near the surface and increase the lifting force on the seeds (Higgins et al., 2003).In this study, underlying surface was also a major factor that affected long-distance dispersal, because the probability of long-distance dispersal in the fixed sand dunes with moss crusts was much higher than that in the other two habitats.Few studies have explicitly indicated that certain seed morphological traits may affect short-distance dispersal or long-distance dispersal (Thomson et al., 2011; Cavanagh et al., 2020).Winged and hairy diaspores, or diaspores with high wing loading may have a tendency for long-distance dispersal (Liang, 2019); the longer the wings, the farther the long-distance dispersal (Guzmán-Vázquez et al., 2023).Furthermore, seeds with high mass tend to disperse over short distance, whereas seeds with low or even extremely low mass tend to disperse over long distance (Batty et al., 2006; Zhou et al., 2019).
Burial is also one of the patterns of secondary seed dispersal.Seed burial is seen as the beginning of a soil seed bank and as a means of germplasm resource conservation (Fenner and Thompson, 2005).In this study, the proportion of seeds that were buried varied among the three habitats, and the semi-fixed sand dunes with algae crusts had the highest proportion (12.0%).This was because algae crusts were more likely to form cracks or micro-pits, and then seeds were more likely to be intercepted (He et al., 2023).However, the layer of moss crusts was tight and thick,making it difficult for the large seeds to be buried after they were dispersed on the surface (Chen et al., 2017).In the mobile dunes, the proportion of seeds that were buried also increased with the increasing wind speed, a result that was related to the movement of the sand itself.
Although the proportion of seeds that were buried varied in different habitats, linear mixed model 3 (Table 2) showed that shape index and wind speed had a significant effect on the burial of seeds.Shape index (variance of 0.11) explained more to seed burial than wind speed (variance of 0.09)(Fig.8c).Shape index refers to the degree to which a seed approaches a sphere.Our result indicated that when the wind speed is less than or equal to 1.4 times TWS, the relationship between shape index and seed burial is positive.This differs from the results reported by Liang et al.(2020), and these inconsistent results may be caused by different selected plant species and wind speed settings.The variance of the burial model was only 0.30, and the marginalR2was 0.21.However, compared with no dispersal (conditionalR2=0.794 and marginalR2=0.777) and short-distance dispersal(conditionalR2=0.756 and marginalR2=0.739), the results of linear mixed model analysis showed that the process of seed burial and its influencing factors may be more complex.
The process and pattern of secondary seed dispersal are crucial for the seeds.In our study, most seeds did not move or only dispersed over short distance, indicating that most seeds were retained within the study area.The seeds of most short-lived plants in this study area have dormant periods(Liu et al., 2004;Wang et al., 2019).Previous studies have found that plant species with high dormancy rates usually have shorter spatial dispersal distance (Chen et al., 2020).Some research also suggested that small, short-lived plant species with persistent seed banks and low mean dispersal distance tend to disperse through time rather than space (Venable et al., 2008; Thomson et al., 2011).Therefore, short-distance dispersal is also the adaptive strategy for the main vegetation (annual herbs) in our study area.It has been suggested that the seeds of most desert annual plants in the Sonoran Desert, USA, do not travel very far (Venable et al., 2008), and the distances of seed dispersal are only 0.49-0.92 m for herbaceous plants in Europe (Willson, 1993).These findings are similar to our research results.In addition, due to the large topographic relief in our study area, seeds tended to accumulate at the bottom of the slope along the windward slope,and shrubs and litter also had a blocking effect on the seeds, all of which limited the long-distance dispersal.Hence, it can be inferred that the soil seed bank in the study area was formed by the short-distance dispersal andin situburial of seeds produced by local plant species.
5 Conclusions
In this study, by conducting field wind tunnel experiments, we preliminarily studied the secondary seed dispersal process induced by wind and its influencing factors in the three habitats on the southeastern edge of the Tengger Desert.The secondary seed dispersal by wind was generally a complex and multi-factor interactive driving process.
The TWS of seeds was affected by the interaction between seed morphological traits and underlying surface.Among the 11 selected plant species, the TWS ofC.korshinskiiwas the highest, and that ofE.gmeliniiwas the lowest.Under the field controlled experimental conditions, most seeds did not disperse or dispersed over short distance; the proportion of seeds that were buried was the lowest, and long-distance dispersal often depended on the high wind speed.For the three habitats, mobile dunes were the most difficult places for seed dispersal, the proportion of seeds that were buried was the highest in the semi-fixed sand dunes, and seeds were most prone to disperse over long distance in the fixed sand dunes.The influencing factors of the four secondary seed dispersal patterns were distinct.Wind speed, underlying surface, and seed morphological traits had a stronger explanatory power for no dispersal and short-distance dispersal than burial and long-distance dispersal.The explanatory power of wind speed,underlying surface, circularity, and TSW for short-distance dispersal decreased successively.Wind speed and underlying surface had a significant impact on the long-distance dispersal, and shape index and wind speed were correlated with seed burial.
Overall, due to the dispersal limitations, most seeds were retained in the study area, which is the main source of vegetation regeneration in the local community.In view of the long-term and complex nature of natural vegetation succession in artificial sand fixation areas, it is necessary to further study the seed dispersal limitation mechanism to better guide the management practice of vegetation regeneration.
Conflict of interest
ZHAO Wenzhi is an editorial board member of Journal of Arid Land and was not involved in the editorial review or the decision to publish this article.All authors declare that there are no competing interests.
Acknowledgements
This research was supported by the Key R & D Program of Ningxia Hui Autonomous Region, China(2021BEG03008) and the Natural Science Foundation of Ningxia Hui Autonomous Region, China(2021AAC03083).We are grateful to the editors and anonymous reviewers for their insightful comments and suggestions in improving this manuscript.
Author contributions
Conceptualization: QU Jianjun, ZHAO Wenzhi, YANG Xinguo; Data curation: QU Wenjie, ZHANG Xue;Formal analysis: QU Wenjie, WANG Lei; Funding acquisition: WANG Lei; Investigation: QU Wenjie, ZHANG Xue, WANG Lei; Methodology: QU Wenjie; Visualization: QU Wenjie; Writing - original draft: QU Wenjie,YANG Xinguo; Supervision: ZHAO Wenzhi, QU Jianjun, YANG Xinguo.All authors approved the manuscript.
Appendix
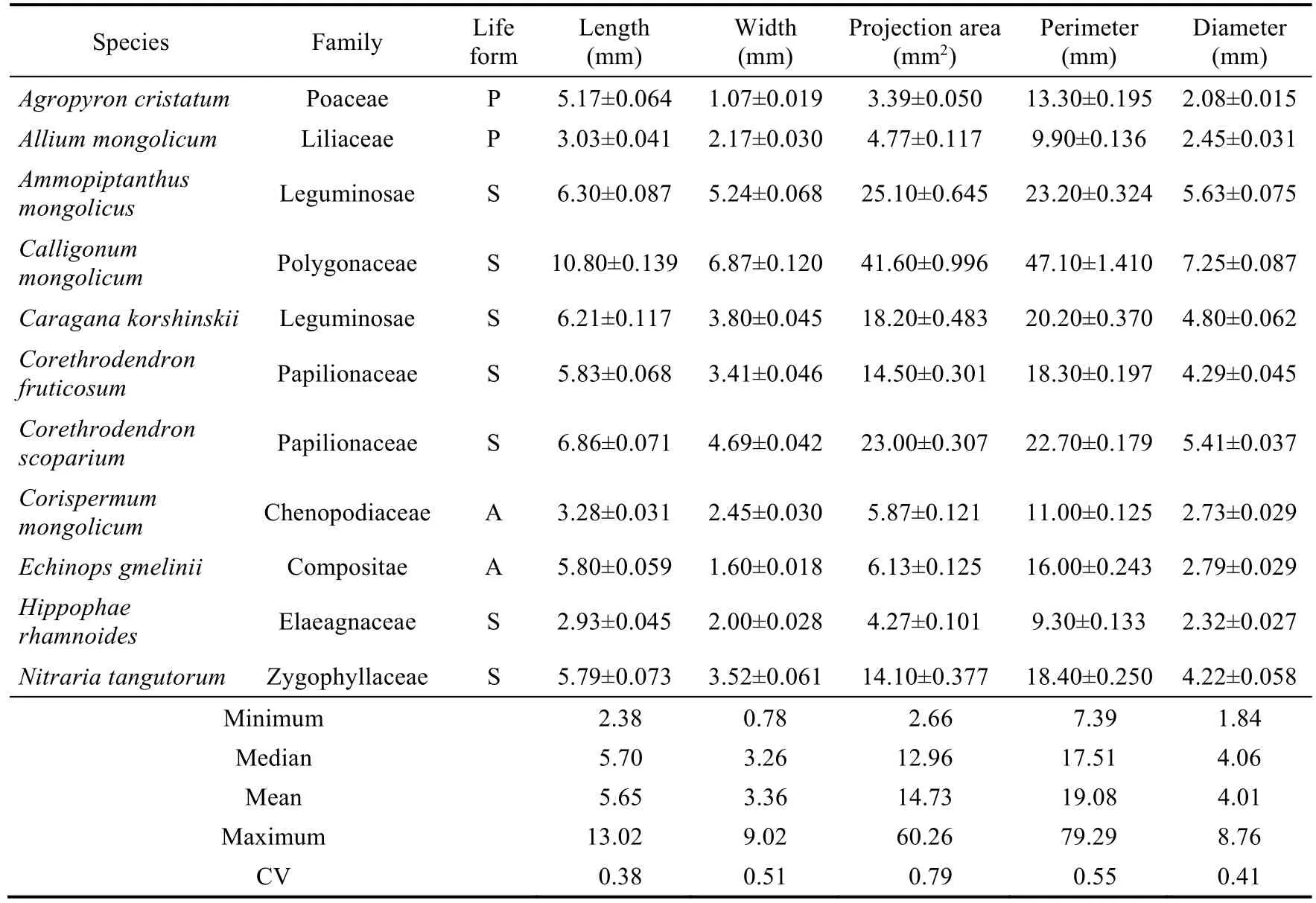
Table S1 Seed morphological traits (length, width, projection area, perimeter, and diameter) of the selected 11 plant species

Table S2 Seed morphological traits (length/width, circularity, compacity, rugosity, TSW, Wing loading, and shape index) of the selected 11 plant species

Fig.S1 Pictures showing the seeds of the selected 11 plant species
杂志排行

Journal of Arid Land的其它文章
- Assessment of runoff changes in the sub-basin of the upper reaches of the Yangtze River basin, China based on multiple methods
- Effect of coir geotextile and geocell on ephemeral gully erosion in the Mollisol region of Northeast China
- Spatiotemporal variability of rain-on-snow events in the arid region of Northwest China
- Urban growth scenario projection using heuristic cellular automata in arid areas considering the drought impact
- Afforestation with an age-sequence of Mongolian pine plantation promotes soil microbial residue accumulation in the Horqin Sandy Land, China
- Spatiotemporal characteristics of seed rain and soil seed bank of artificial Caragana korshinskii Kom.forest in the Tengger Desert, China