马铃薯WOX 基因家族的鉴定及在离体再生和非生物胁迫中的表达分析
2024-04-17张玉石磊巩檑聂峰杰杨江伟刘璇杨文静张国辉颉瑞霞张丽
张玉 石磊 巩檑 聂峰杰 杨江伟 刘璇 杨文静 张国辉颉瑞霞 张丽,2
(1.宁夏农林科学院农业生物技术研究中心 宁夏农业生物技术重点实验室,银川 750002;2.甘肃农业大学农学院,兰州 730070;3.甘肃农业大学生命科学技术学院,兰州 730070;4.甘肃农业大学省部共建干旱生境作物学国家重点实验室,兰州 730070;5.宁夏农林科学院固原分院,固原 756000)
马铃薯(Solanum tuberosum L.)是世界上第四大粮菜兼用作物,分布范围广、适应性强、种植面积大,在保障我国粮食安全、促进经济战略性调整、全面推进乡村振兴等方面发挥重要支撑作用。马铃薯为同源四倍体无性繁殖作物,基因组高度杂合,且自交不亲和,加快生物育种创制优良种质势在必行。但马铃薯基因型依赖性严重,部分品种转化效率较低。因此,提高马铃薯遗传转化效率、克服基因型依赖性问题,对充分发挥生物技术在马铃薯遗传育种中的作用意义重大。
WUSCHE‑ 相 关 同 源 盒(WUSCHEL‑related homeobox, WOX)基因家族是一种植物特异性转录因子家族,其保守序列是由约60 个氨基酸以“螺旋-环-螺旋-转角-螺旋(helix‑loop‑helix‑turn‑helix)”构成的同源异型结构域(homeo‑domain, HD),WOX转录因子通过HD 与特定DNA 序列结合发挥生物学功能,广泛参与植物生长发育、干细胞分裂分化调控、胚胎和器官形成等过程[1]。通过对不同植物中WOX 蛋白的系统发育分析,WOX 基因家族被分为3 个分支:WUS 进化分支(WUS clade)、中间进化分支(intermediate clade)、古老进化分支(ancient clade)[2]。1996 年,Laux 等[3]从拟南芥(Arabidopsis thaliana)中发现了WUS 基因,首次揭示了WOX 基因家族成员在植物发育中的重要作用。随后,WOX基因家族在拟南芥、水稻、高粱、玉米等多种植物中进行了充分的研究[4],其家族成员在植物发育和生长过程中发挥着重要调控作用。在拟南芥中,WUS 基因主要在茎尖分生组织(shoot apical meris‑tem, SAM)表达,对SAM 的形成和维持具有重要意义[3],研究表明玉米WUS 基因与再生基因BBM 在植物组织培养过程中发挥了关键作用,二者共转化可显著提高玉米、高粱、甘蔗的转化效率[5]。WOX1和WOX3 在拟南芥、黄瓜、柳枝稷等植物的器官侧向生长中起重要作用[6-8]。WOX2 在早期胚胎细胞发育过程中发挥功能,云杉PaWOX2 可作为胚胎发生的潜力标记[9],同样地,WOX8 和WOX9 也是胚胎发育所必需的[10]。WOX4 能够促进拟南芥和番茄维管形成层的分化和维持[11]。WOX5 在根尖分生组织(rootapical meristem, RAM)中表达,并维持RAM 中干细胞的稳定性[12],同时过表达TaWOX5 能够显著提高小麦转化效率、再生能力[13]。研究证明WOX6主要调控胚珠和种子发育,在珠被和卵细胞形成的过程中防止细胞提前分化[14]。拟南芥WOX7 在侧根发育的所有阶段表达,并以糖依赖性的方式抑制侧根发育,过表达AtWOX7 将显著减少植株侧根原基数量[15]。WOX11 和WOX12 在调控根系发育上功能冗余,过表达PagWOX11/12a 可促进银腺杨根系伸长、生物量增加[16]。WOX13 和WOX14 与花器官的形成和发育相关,过表达OsWOX13 可使水稻开花期提早7-10 d[17],AtWOX14 能够促进拟南芥花序轴中维管细胞的分化和木质化[18]。此外,WOX 基因家族成员还参与逆境胁迫应答,其中,AtWOX6 在根尖的维持和根发育中起着重要作用,在冷胁迫应答中扮演重要角色[19],茶树CsWOXs 在PEG 诱导的干旱胁迫、盐胁迫处理下存在差异表达,响应非生物胁迫[20],过表达水稻OsWOX13 能够提高水稻耐旱性[17],敲除棉花GhWOX4 导致转基因棉花耐旱性显著降低[21],过表达PagWOX11/12a 可显著增强银腺杨耐旱性与耐盐性[16,22]。
WOX 基因家族成员参与调控植物生长发育、响应逆境胁迫等重要过程,已在多种植物中得到鉴定,但目前对马铃薯WOX 基因家族的鉴定及基因功能的研究尚未见报道。
本研究利用马铃薯基因组信息,通过生物信息学方法对马铃薯WOX 基因家族成员进行鉴定,分析其基因结构、转录因子结合位点、染色体分布、表达模式等。通过愈伤组织培养与RT‑qPCR 试验,证明StWOX5 在离体再生方面的功能,为进一步解析马铃薯StWOXs 功能及调控植物生长发育分子机制,以及利用目标基因在马铃薯育种研发与应用方面提供理论依据。
1 材料与方法
1.1 材料
试验材料为马铃薯品种(系)大西洋(DY)、84、85、Z291,由宁夏农林科学院农业生物技术研究中心(宁夏农业生物技术重点实验室)提供。
1.2 方法
1.2.1 马铃薯WOX 家族成员蛋白鉴定 使用HM‑MER 3.0 软件对拟南芥、番茄、烟草、水稻 WOX 蛋白序列构建隐马尔可夫模型,并以此模型搜寻马铃薯所有编码蛋白序列,再从中找出潜在的WOX 家族序列。使用BLASTP[23]将马铃薯所有蛋白序列与获取的WOX 家族参考序列进行比对(e‑value 设为1e-20),保留比对成功的同源序列作为候选WOX家族蛋白序列。利用软件pfamscan 和Pfam A[24]数据库对获得的候选序列进行结构域注释,将含有PF00046 结构域的序列作为最终马铃薯WOX 序列。
1.2.2 马铃薯WOX 成员共线性分析 使用MCS‑canX 软件,采用默认参数(MATCH SCORE: 50;MATCH SIZE: 5; GAP PENALTY: ‑1; OVERLAP WIN‑DOW: 5; E VALUE: 1e-05; MAX GAPS: 25),在马铃薯WOX 家族种内和种间进行基因共线性分析。
1.2.3 马铃薯WOX 基因家族成员理化性质分析、亚细胞定位 利用在线软件ExPASy 分析马铃薯WOX家族蛋白的氨基酸长度、分子量、等电点、不稳定指数等理化性质。通过ProtComp 9.0 对马铃薯WOX基因家族成员进行亚细胞定位预测。
1.2.4 马铃薯WOX 系统发育进化树构建及分析 使用mafft 软件对马铃薯、拟南芥、番茄、烟草、水稻WOX 家族蛋白序列进行多序列比对,然后,将比对结果导入MEGA 软件,采用邻接法(neighbor‑joining method, NJ)构建系统发育进化树(参数设置:模型为p‑distance,缺失数据方法为Partial deletion,cutoff 为50%,Bootstrap 设 置 为1 000)。 在 软 件iTOL 中对进化树进行注释。
1.2.5 StWOXs 基因结构、蛋白motif、转录因子结合位点分析 使用软件GSDS 分析马铃薯WOX 基因家族成员的内含子/外显子结构分布。利用MEME软件分析家族成员的motif 结构(motif 寻找数量为15)。截取基因结构上游2 000 bp 区域作为启动子调节序列,利用PlantTFDB 软件预测启动子上的转录因子结合位点(transcription factor binding site,TFBS),在启动子物理图谱中标记和显示数量排名前12 的结合位点,并统计分析StWOXs 启动子上TFBS 的类型和数量。
1.2.6 StWOXs 表达模式分析及愈伤组织诱导分化培养 基于PGSC 下载的马铃薯转录组(RNA‑seq)数据,对StWOXs 表达量数据做层次聚类分析,将具有相同或相似表达行为的基因进行聚类。根据聚类图,选择与马铃薯离体再生相关的基因StWOX5进行功能分析。以马铃薯大西洋(DY)、84、85、Z291 四个具有不同再生能力品种(系)的无菌脱毒苗作为试验材料,取茎段置于MS 基础培养基中,每皿初始放置25 个茎段,共3 皿。外植体接种10、20 和30 d 后,统计不同品种(系)愈伤组织个数,计算出愈率。待愈伤组织诱导成功后转入分化培养基,统计10、20 和30 d 后不同品种(系)的分化率,并通过实时荧光定量PCR(RT‑qPCR)测定不同品种StWOX5 在分化阶段的相对表达量。愈伤组织诱导率(%)=诱导出愈伤组织的茎段数/茎段总数×100%,分化率(%)=分化茎段数/茎段总数×100%。荧光定量PCR 试验步骤为:使用试剂盒法提取不同马铃薯品种(系)分化外植体RNA,反转录得到cDNA 测定StWOX5 的表达情况。以EF1α 为内参,采用荧光染料法进行实时荧光定量PCR(引物见表1),应用2-△△CT法计算StWOX5相对表达量[25],每个检测样品进行3 次重复试验。PCR 反应程序设置为95℃ 30 s;95℃ 10 s,60℃ 10 s,72℃ 15 s,40 个循环;72℃ 10 min。
表1 实时荧光定量PCR 引物Table 1 Primers used in real-time quantitative PCR
1.2.7 数据分析 StWOX5 RT‑qPCR 试验数据均为3次生物学重复的平均值。使用Graphpad Prism 8.0 软件作图、SPSS 23.0 软件进行显著性分析。
2 结果
2.1 马铃薯WOX基因家族成员
经比对、分析,共获得11 个马铃薯WOX 基因家族成员,它们随机分布在第2、3、4、6、11 染色体上,其中,第2 和6 染色体上分别分布了3 个WOX 成员,数量最多,第4 染色体上WOX 成员分布最少(1 个)。基于邻接树进化关系,对马铃薯WOX 基因家族成员按分支结果进行命名。理化性质及亚细胞定位信息如表2 所示,11 个马铃薯WOX蛋白的氨基酸数量为157-391,相对分子质量为18.5-44.1 kD;等电点处于5.53-9.97,其中5 个成员的理论等电点小于7.0,占比45%;预测所有马铃薯WOX 成员均定位于细胞核内。

表2 马铃薯WOX 基因家族成员信息Table 2 Information of WOX gene family members in S.tuberosum
2.2 马铃薯WOX成员共线性分析
为探究马铃薯WOX 家族的基因重复事件,对马铃薯WOX 成员之间的种内共线性,以及拟南芥、番茄、水稻、马铃薯WOX 的种间共线性进行分析。结果(图1‑A)表明,马铃薯StWOX3c和StWOX3b 发生了序列复制事件,二者具有共线性。马铃薯StWOXs 和拟南芥AtWOXs 中有7 个同源基因对,分别为StWUS/AtWUS、StWOX1/AtWOX1、StWOX2/AtWOX2、StWOX3a/AtWOX3、StWOX4/AtWOX4、StWOX5/AtWOX5 和StWOX13/AtWOX13;马铃薯StWOXs 和番茄SlWOXs 中有10 个同源基因对,分 别 为StWUS/SlWUS、StWOX1/SlWOX1、StWOX2/SlWOX2、StWOX3b/SlWOX3b、StWOX3c/SlWOX3b、StWOX4/SlWOX4、StWOX5/SlWOX5、StWOX9/SlWOX9、StWOX11/SlWOX11 和StWOX13/SlWOX13;马铃薯StWOXs 和水稻OsWOXs 之间无同源基因对(图1‑B)。

图1 马铃薯WOX 成员共线性分析Fig.1 Collinear analysis of potato WOX members
2.3 系统发育进化树分析
为明确马铃薯WOX 基因家族的进化关系,对马铃薯、拟南芥、番茄、烟草、水稻5 个物种共58 个蛋白序列进行多重比对并构建系统发育进化树。结果(图2)显示,58 个蛋白序列分为3 个类群:WUS 进化分支、中间进化分支和古老进化分支,每个类群分别含有33、15 和10 个WOX 成员。11 个马铃薯WOX 成员分布在3 个不同的分支中,其中,WUS 进化分支中成员最多,包含StWUS、StWOX1、StWOX2、StWOX3a、StWOX3b、StWOX3c、StWOX4、StWOX5 共8 个 蛋 白, 与 其分在同一类群的还有8 个拟南芥WOX 成员(At‑WUS、AtWOX1/2/3/4/5/6/7)、6 个 水 稻WOX 成 员(OsWOX1/2/3/4/5/9)、4 个烟草WOX 成员(NtabWUS、NtabWOX4a/4b/5)、7 个 番 茄WOX 成 员(SlWUS、SlWOX1/2/3a/3b/4/5);中间进化分支中含有2 个马铃薯WOX 成员(StWOX9/11),与其分在同一类群的有4 个拟南芥WOX 成员(AtWOX8/9/11/12)、6个 水 稻WOX 成 员(OsWOX6/7/10/11/12/13)、1 个烟草WOX 成员(NtabWOX11)、2 个番茄WOX 成员(SlWOX9/11);古老进化分支中仅含1 个马铃薯WOX 家族成员(StWOX13),与其分在同一类群的有 3 个拟南芥WOX 成员(AtWOX10/13/14)、1 个水稻WOX 成员(OsWOX8)、4 个烟草WOX 成员(NtabWOX13a/13b/13c/13d)、1 个 番 茄WOX 成 员(SlWOX13)。通过比对分析发现,马铃薯WOX 基因家族成员与同为茄科的番茄同源关系最近,且所有亚家族中均含有马铃薯、拟南芥、番茄中的相应成员,预示着WOX 基因家族成员分化的时间可能在马铃薯、拟南芥、番茄物种分化时间之前。

图2 马铃薯、拟南芥、番茄、烟草、水稻WOX 蛋白的系统发育分析Fig.2 Phylogenetic analysis of WOX proteins among S.tuberosum, A.thaliana, S.lycopersicum, N.tabacum, and O.sativa
2.4 StWOXs基因结构与蛋白motif分析
为了解StWOXs 基因结构特征,对该家族成员基因的内含子/外显子分布进行了分析。由图3 可知,StWOXs 基因结构相对简单,外显子数目2-6个,内含子数目1-5 个。其中5 个基因(StWOX2、StWOX3a、StWOX3b、StWOX3c、StWOX5) 外 显子 最 少, 仅 有2 个。 另 外,4 个 基 因(StWUS、StWOX2、StWOX9、StWOX11)没有UTR 结构。利用MEME 在线软件,分析确定了马铃薯StWOX 蛋白的15 个保守基序,命名为motif1-motif15。由图4可知,motif1 和motif2 是除StWOX9 蛋白外其他10个StWOX 蛋白所共有的。不同进化分支中,StWOX蛋白的motif 组成差异较大。其中,motif3 仅存在于WUS 分支,motif5 和motif6 为中间进化分支成员所特有。

图3 StWOXs 基因结构示意图Fig.3 Structure of StWOX genes
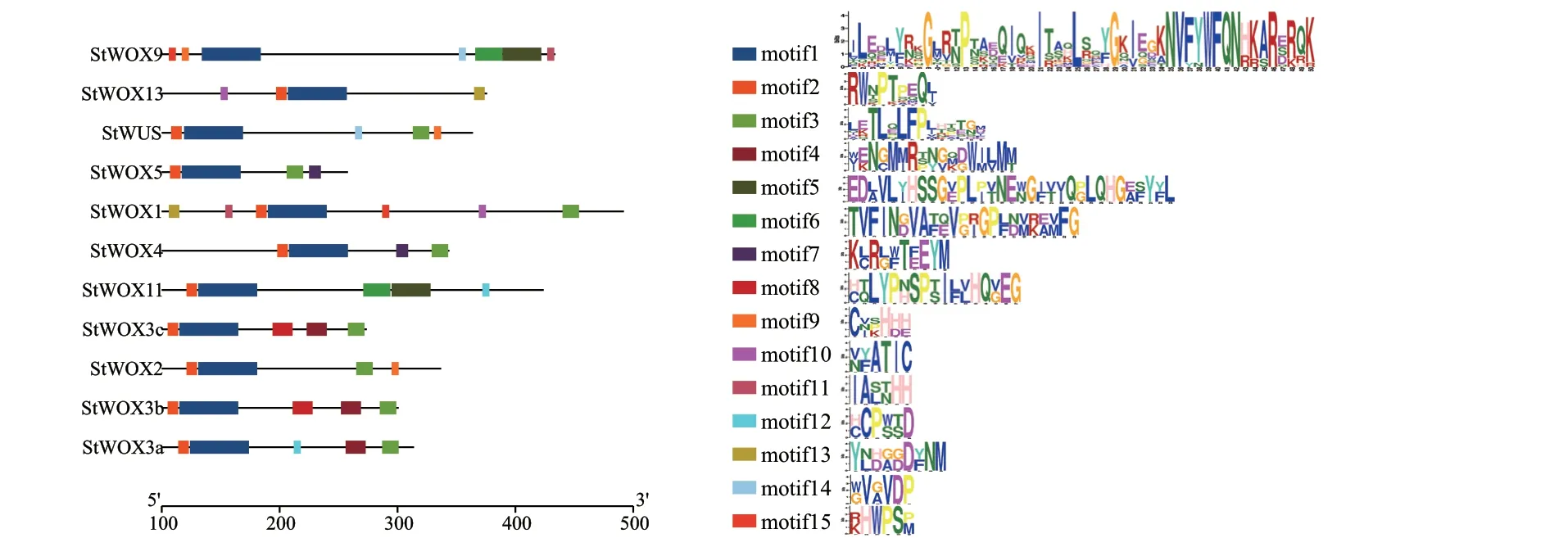
图4 StWOX 蛋白motif 分布及其序列标识Fig.4 Distribution and sequence logos of motifs in StWOX proteins
2.5 StWOXs启动子区转录因子结合位点分析
为研究转录因子对StWOXs 表达的调控,提取StWOXs序列上游2 000 bp 区域作为启动子调节序列,预测启动子序列中的TFBS 及其数量(图5),Dof、C2H2、MYB、ERF、MYB_related、MIKC_MADS、BBR‑BPC、GATA、bZIP、M‑type_MADS、WRKY、bHLH 是排名前12 的转录因子家族,它们与光响应、植物激素响应、逆境胁迫响应以及生长发育相关。Dof、C2H2、MYB、ERF、MYB_related、bZIP 在所有马铃薯WOX 基因家族成员中均预测出现,Dof 转录因子家族的结合位点数量最多(102 个,含“+、‑”双链),bHLH 的结合位点数量最少(24 个,含“+、‑”双链)。不同进化分支间、不同成员间TFBS 数量和类别存在差异,StWOX5 启动子区域的TFBS 最多(103个),其中ERF 和GATA 数量均大于20;StWOX3a启动子区域的TFBS 数量最少(17 个)。

图5 StWOXs 启动子区域转录因子结合位点预测及数量统计Fig.5 Prediction and numerical statistics of TFBS in the promoter region of StWOXs
2.6 StWOXs表达模式分析
11 个StWOX 在马铃薯根、茎、叶、花、果实、愈伤组织等不同组织中表达差异显著(图6‑A),StWUS 在叶柄组织中高表达;StWOX1、StWOX3a、StWOX3c 在茎中表达量最高;StWOX2、StWOX9 在果实中的表达量较高;StWOX3b 主要在花中表达;StWOX4、StWOX5、StWOX11、StWOX13 均在愈伤组织中得到大量积累,推测在植物离体再生过程中可能发挥一定作用。在干旱胁迫下(图6‑B),StWUS、StWOX1、StWOX3c、StWOX4、StWOX5、StWOX13表达量上调;在盐胁迫下,StWOX3a、StWOX11上 调 表 达;在 高 温 处 理 下(图6‑C),StWOX4、StWOX5、StWOX13 表达上调,初步说明StWOXs 在马铃薯响应逆境胁迫中发挥作用。

图6 StWOXs 在不同组织部位(A)、干旱和盐胁迫(B)、热胁迫(C)中的表达模式Fig.6 Relative expression patterns of StWOXs in different tissues(A), under drought and salt stresses(B), and under heat stress(C)
2.7 愈伤组织诱导分化及StWOX5在分化过程中的RT‑qPCR验证
为验证StWOX5 在马铃薯离体再生过程中的功能作用,将StWOX5 作为候选基因,分析其在不同马铃薯品种(系)愈伤组织分化过程中的表达模式。结果显示,接种10 d 后,4 个品种(系)愈伤组织诱导率呈显著差异(P<0.05),其中84 号出愈率最高,其次为Z291 号,DY、85 号最低。在接种20 d后出愈率无明显差异,除85 号外,其余出愈率均达到100%,且愈伤组织颜色较绿,结构松散,呈纺锤状生长(图7‑A-B)。分化培养30 d 后,4 个品种(系)愈伤组织分化效果呈现显著差异(P<0.05),其中84号分化率最高,达55.1%,85 号愈伤组织无绿色芽点出现,分化率最低(图7‑C-D)。不同品种(系)愈伤组织分化阶段StWOX5 的RT‑qPCR 结果表明(图7‑E),在分化20 和30 d 后,84 号StWOX5 的相对表达量均最高,而85 号最低,这与愈伤组织分化率趋势一致,说明分化过程中目标基因的相对表达量与愈伤组织分化率呈正相关,初步验证了StWOX5参与马铃薯离体再生过程。

图7 马铃薯不同品种(系)愈伤组织诱导分化及StWOX5 的RT-qPCR 验证Fig.7 Callus induction and differentiation of different potato varieties(lines), and RT-qPCR verification of StWOX5
3 讨论
WOX 是植物中特有的转录因子家族,在植物生长发育、干细胞分化、植株离体再生等过程中发挥重要作用[26]。目前,WOX 基因家族已在拟南芥、玉米、番茄、芥菜等各类植物中得到鉴定[4,27-28],但马铃薯WOX 基因家族相关研究尚未见报道。
本研究从马铃薯基因组中鉴定到11 个WOX 基因家族成员,它们随机分布在5 条染色体上,对比发现马铃薯WOX成员比同为茄科植物的番茄多1个,即StWOX3c[29]。番茄和马铃薯中均包含WOX3a、WOX3b,分布在各自的第11 染色体上,而马铃薯StWOX3c 分布在第6 染色体上,且与StWOX3b 具有共线性,可能是因为在物种进化过程中发生了序列复制事件,推测StWOX3c 由StWOX3b 复制而来。分析物种间共线性发现StWOX 基因与拟南芥AtWOX基因、番茄SlWOX 基因之间存在多个同源基因对,表明StWOX 基因可能和拟南芥、番茄WOX 基因的同源基因来源于相同祖先,在调控植物生长发育和响应逆境胁迫过程中发挥类似作用。对11 个马铃薯WOX 基因成员的基本信息进行分析,各成员亚细胞定位均在细胞核内,氨基酸数量、等电点、相对分子质量等理化性质存在一定程度差异,该结果与李晓旭等[29]对番茄WOX 全基因组研究的结果相似,可能预示着马铃薯WOX 基因家族成员参与或调控细胞核内的基因表达和遗传信息的传递,且不同成员在植物生长发育过程中发挥的作用不同。前期研究结果表明,WOX 基因家族成员可分为3 个分支[1],马铃薯WOX 转录因子家族成员同样分成WUS、中间和古老共3 个进化分支,且每个亚家族中都至少有1 个成员与番茄、拟南芥WOX 相应亚家族中的成员存在直系同源关系,反映出WOX 基因家族成员分化的时间可能在马铃薯、拟南芥、番茄物种分化时间之前。结构分析表明,不同进化分支间,基因外显子和内含子数量、蛋白motif 组成、启动子区转录因子结合位点类型和数量存在不同程度的差异。对启动子区的转录因子结合位点预测发现,数量最多的转录因子家族为Dof、C2H2、MYB、ERF 等,均具有调控植物生长和发育、应答环境胁迫等功能[30-33]。例如,Dof 广泛参与种子萌发、维管的发育和叶片极性、花和花粉发育、光周期反应,参与植物逆境响应等过程[34],C2H2参与调节植物器官生长、控制根的分生组织和叶片的极性等植物正常生长发育的各个进程,同时也参与植物生物和非生物胁迫下逆境应答[31],这预示马铃薯WOX 基因家族成员也在植物生长发育、器官形态建成、响应逆境胁迫等方面参与调控,具体调控机制有待研究。
植物特异性转录因子WOX 基因通常具有组织表达特异性和响应逆境胁迫的能力,绿豆中VrWOX5 在种子中表达量较高、VrWOX6 则在叶片中大量表达[35];烟草NtWOX9 在花器官中特异性表达、NtWOX5b 则在根中表达量较高[36];茶树CsWOX1/2/3/20/28/29 在干旱胁迫下表达量较高、CsWOX21 在盐胁迫下表达量上调[20]。与上述结果相似,StWOXs 同样表现出组织表达特异性和响应非生物胁迫的能力。StWOX4/5/11/13 在愈伤组织中特异表达;StWUS、StWOX1/3c/4/5/13 在甘露醇处理后上调表达;StWOX3a/11 在盐胁迫下上调表达;StWOX4/5/13 在热胁迫下表达量升高,暗示相应基因可能参与马铃薯离体再生过程和非生物胁迫响应。
WOX 基因的表达水平能够影响植物组织培养过程中外植体愈伤组织的再生能力[37]。筛选在愈伤组织中高表达的基因StWOX5 进行验证发现,在离体再生能力不同的4 个马铃薯品种(系)中,StWOX5的相对表达量与愈伤组织分化率呈正相关,进一步证明StWOX5 在马铃薯离体再生过程中发挥重要作用,类似地,在苦荞中也发现FtWOX 的表达模式与苦荞出愈率呈显著正相关关系[38]。同时已有研究表明过表达小麦TaWOX5 能够显著提高小麦的转化效率[13],推测过表达StWOX5 可能会提高马铃薯外植体产生愈伤组织的能力,这将对提高马铃薯遗传转化效率、促进基因编辑等技术在马铃薯遗传育种中的应用意义重大。
4 结论
共鉴定得到11 个马铃薯WOX 基因家族成员,马铃薯WOX 基因家族成员被分成3 个进化分支,即WUS 分支、中间分支和古老分支,不同分支中StWOXs 的基因结构、蛋白motif 组成、启动子区转录因子结合位点类型和数量存在差异。StWOXs 具有组织表达特异性和响应非生物胁迫的能力。在具有不同再生能力的马铃薯品种(系)中,StWOX5 的相对表达量与愈伤组织分化率呈正相关,证明StWOX5在马铃薯离体再生过程中的作用。
