饲粮中添加甜菜碱和表没食子儿茶素没食子酸酯对育肥猪生长性能、肉品质、血清生化和抗氧化指标的影响
2024-04-02陈雨诗刘禹熙杨童寓丹王新霞
陈雨诗 刘禹熙 杨童寓丹 王新霞
(浙江大学动物科学学院,浙江大学动物分子营养教育部重点实验室,农业农村部(华东)动物营养与饲料重点实验室,浙江省饲料与动物营养重点实验室,杭州 310058)
我国是猪肉生产大国,也是猪肉消费大国,猪肉在我国肉类消费品种中占有重要地位。随着民众生活和消费水平的提高,肉品质越来越受到重视,提升猪肉品质已经成为当下生猪产业的关键目标。“杜×长×大”三元杂交猪(以下称杜长大猪)是我国目前养殖的主要品种,具有生长速度快、饲料利用率高的特性,然而其肉品质与我国优良地方品种猪相比,还是有肌内脂肪含量低、风味差等缺陷。因此,如何通过营养调控手段提高杜长大猪的肉品质和风味是养殖产业亟需解决的关键问题。
甜菜碱是甘氨酸的三甲基衍生物,在机体营养代谢中起着十分重要的作用[1]。研究发现,甜菜碱具有促进生长发育、改善胴体品质及调节脂代谢的功效,因此被广泛应用到畜禽生产中[2]。甜菜碱在提升肌内脂肪含量和改善肉品质方面具有良好的效果,饲粮中添加1 g/kg甜菜碱能显著提高猪肉肌内脂肪含量[3]。Martin等[4]研究发现,饲粮中长期添加甜菜碱能在不影响肌肉理化特性的情况下增加猪肉肌内脂肪沉积。本课题组前期研究发现,在“杜×长×大”三元杂交仔猪的饲粮中添加1 000 mg/kg的甜菜碱能提高仔猪的生长性能,降低背膘厚,同时还能显著提高肌内脂肪含量[5];在生长育肥猪的饲粮中添加1 000和1 500 mg/kg的甜菜碱也能降低背膘厚,并提高肌内脂肪含量[6]。本课题组对于甜菜碱调控肌内脂肪沉积的机制进行了深入研究,发现甜菜碱能通过上调肝脏甜菜碱高半胱氨酸甲基转移酶(BHMT)表达促进肝脏产生还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH),并提高机体循环及骨骼肌NADPH含量,进而上调肌内脂肪细胞中肥胖相关蛋白(fat mass and obesity-associated protein,FTO)表达,从而降低了肌内脂肪细胞的mRNA上的N6-甲基腺苷(m6A)水平,促进肌内脂肪细胞克隆增殖,进而促进肌内脂肪细胞分化聚酯,最终提高了肌内脂肪含量[7]。
表没食子儿茶素没食子酸酯(EGCG)是绿茶中主要的活性物质[8],已有研究表明EGCG可以有效降低动物脂肪沉积[9]。本课题组前期研究发现,EGCG可以通过抑制FTO表达,提高细胞克隆增殖关键基因细胞周期蛋白A2(cyclin A2,CCNA2)和周期蛋白依赖性激酶2(cyclin-dependent kinase 2,CDK2)的m6A修饰水平,从而招募了YTH结构域家族蛋白2(YT521-B homology domains 2,YTHDF2)的靶向结合,抑制其蛋白表达,导致细胞周期延迟,阻抑了前体脂肪细胞到脂肪细胞的分化聚酯过程[10]。本课题组将EGCG应用于金华猪养殖生产,结果表明基础饲粮中添加0.3%的EGCG可以有效减少金华猪的平均背膘厚和血清甘油三酯(TG)含量,同时还显著提高了金华猪眼肌面积[11]。
基于上述研究结果,本试验假设饲粮中同时添加甜菜碱和EGCG能在提高杜长大猪肌内脂肪含量的同时进一步降低背部脂肪沉积,并提高肉品质和生长性能。因此,本试验旨在研究饲粮中添加甜菜碱和EGCG对杜长大猪生长性能、肉品质、血清生化指标及抗氧化能力的影响,重点分析甜菜碱和EGCG联合应用对育肥猪背膘厚、肌内脂肪沉积的影响,为甜菜碱和EGCG在生猪养殖中的应用提供理论基础和科学依据。
1 材料与方法
1.1 试验设计和饲养管理
选取81头平均体重为90.03 kg、健康的育肥期杜长大猪,随机分为3组,每组3个重复,每个重复9头猪。对照组饲喂基础饲粮,试验Ⅰ组在基础饲粮中添加2 500 mg/kg的甜菜碱,试验Ⅱ组在基础饲粮中添加2 500 mg/kg的甜菜碱和1 000 mg/kg的EGCG(有效含量为98%)。
饲养试验在浙江科强生态养殖有限公司进行,预试期5 d,正试期40 d,整个过程中保证试验猪自由饮水和采食。试验饲粮组成及营养水平见表1。
表1 试验饲粮组成及营养水平(风干基础)
1.2 样本采集
试验结束时,每个重复随机选择2头(每组6头)健康且体重相近的试验猪,禁食12 h后屠宰。采用颈静脉采血法采集血液样本,4 000 r/min离心10 min得到血清,用于血清生化指标分析;采集背最长肌,液氮速冻后保存于-80 ℃冰箱,用于氨基酸、脂肪酸含量和分子生物学分析。
1.3 指标检测
1.3.1 饲粮营养成分
依照《饲料中粗蛋白测定方法》(GB/T 6432—1994)、《饲料中粗脂肪的测定》(GB/T 6433—2006)、《饲料中粗灰分的测定》(GB/T 6438—2007)、《饲料中钙的测定》(GB/T 6436—2018)和《饲料中总磷的测定》(GB/T 6437—2018)测定粗蛋白质、粗脂肪、粗灰分、钙和总磷含量,然后依据《饲料中氨基酸的测定》(GB/T 18246—2000)用高效液相色谱(HPLC)法测定氨基酸含量。饲粮中消化能计算方法为饲料总能与粪能的差值[12]。
1.3.2 生长性能
试验猪于试验开始和试验结束时在晨饲前逐头称重。根据试验期间每日记录的每次给料量、剩余料量和损耗料量,以重复为单位计算平均日采食量,同时根据试验猪个体重计算平均日增重和料重比。
1.3.3 胴体及肉质指标
试验结束时,按照《瘦肉型猪胴体性状测定技术规范》(NY/T 825—2004)、《猪肉品质测定技术规范》(NY/T 821—2019)分别测定胴体性状和肉品质。
1.3.4 背最长肌氨基酸和脂肪酸组成
参照《食料中氨基酸的测定》(GB/T 18246—2000),利用日立氨基酸自动分析仪测定背最长肌中的脂肪酸组成;参照《食品中脂肪酸的测定》(GB 5009.168—2016),利用气相色谱仪测定背最长肌中的脂肪酸组成。
1.3.5 血清生化和抗氧化指标
血清生化指标:采用南京建成生物研究所的试剂盒测定血清甘油三酯、总胆固醇(TC)、低密度脂蛋白胆固醇(LDL-C)和高密度脂蛋白胆固醇(HDL-C)含量;血清抗氧化指标:采用南京建成生物研究所的试剂盒测定血清谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量及总抗氧化能力(T-AOC)。
1.3.6 背最长肌中脂肪沉积、肌纤维类型相关基因mRNA表达
采用Trizol试剂盒(大连宝生物工程有限公司)提取背最长肌的总RNA,参照ABScript Ⅲ RT Master Mix for qPCR(武汉爱博泰克生物科技有限公司)逆转录试剂盒说明书对总RNA样品进行逆转录合成cDNA。采用荧光定量PCR(qPCR)技术检测背最长肌中脂肪沉积、肌纤维类型相关基因mRNA相对表达水平。10 μL反应体系:2×SYBR Green qPCR Master Mix 5 μL,cDNA 4 μL,上、下游引物各0.5 μL。反应条件:95 ℃预变性3 min,95 ℃变性5 s,60 ℃退火35 s,共40个循环。根据目的基因和内参基因β-肌动蛋白(β-actin)的阈值循环(Ct),采用2-ΔΔCt法计算样品中各目的基因mRNA相对表达水平。引物序列见表2。
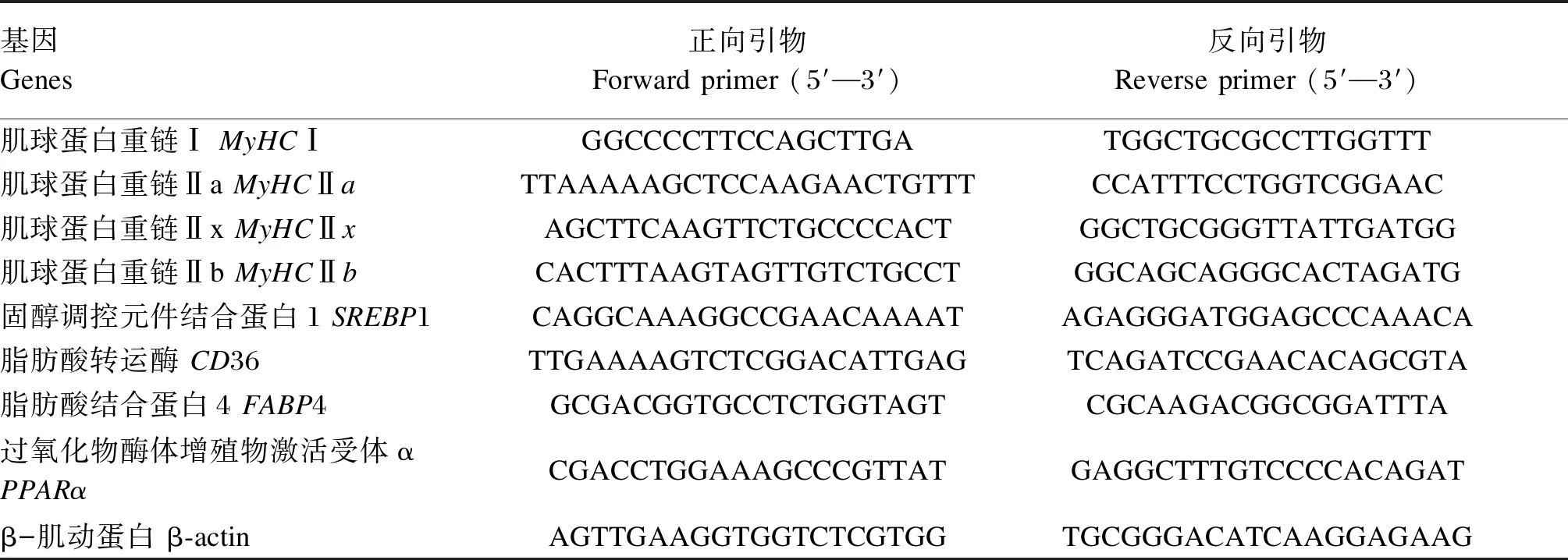
表2 引物序列
1.4 数据统计分析
所有试验数据均以平均值和标准误(SE)表示,采用GrapPhad 9.0软件进行单因素方差分析(one-way ANOVA),并采用Duncan氏多重比较检验法进行组间比较,以P<0.05作为差异显著性判断标准。
2 结 果
2.1 饲粮中添加甜菜碱和EGCG对杜长大猪生长性能的影响
由表3可知,3组试验猪的终末体重、平均日增重和料重比均无显著差异(P>0.05)。与对照组相比,试验Ⅰ、Ⅱ组的平均日采食量均显著降低(P<0.05),试验Ⅰ组降低了7.23%,试验Ⅱ组降低了11.14%;试验Ⅰ、Ⅱ组之间平均日采食量无显著差异(P>0.05)。

表3 饲粮中添加甜菜碱和EGCG对杜长大猪生长性能的影响
2.2 饲粮中添加甜菜碱和EGCG对杜长大猪肉品质的影响
2.2.1 饲粮中添加甜菜碱和EGCG对杜长大猪胴体和肉质指标的影响
由表4可知,3组试验猪的宰前体重、胴体重、胴体长、屠宰率和眼肌面积均无显著差异(P>0.05)。与对照组相比,试验Ⅰ、Ⅱ组的背膘厚均显著降低(P<0.05),试验Ⅰ组降低了13.94%,试验Ⅱ组降低了15.31%;试验Ⅰ、Ⅱ组之间背膘厚无显著差异(P>0.05)。
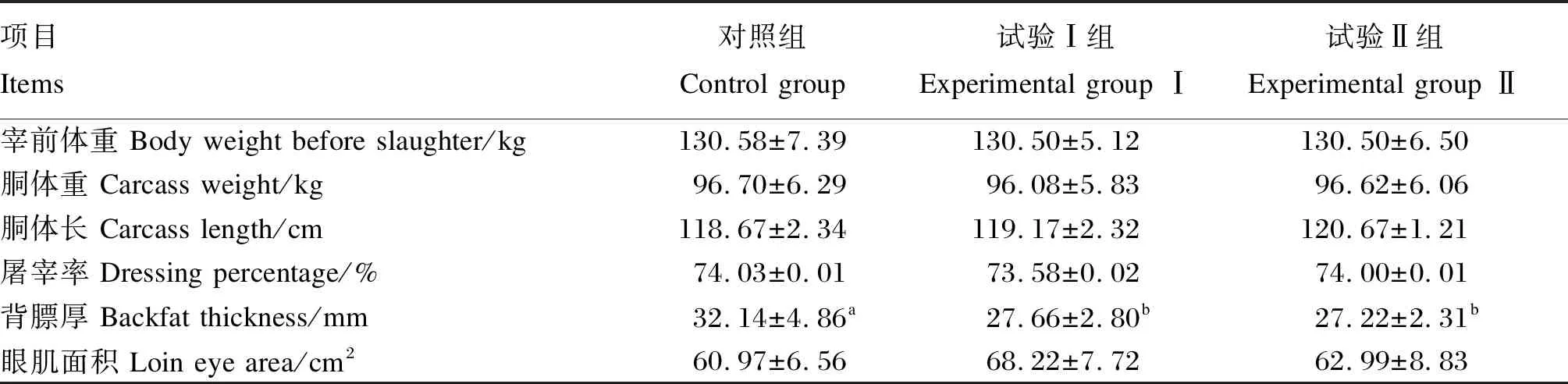
表4 饲粮中添加甜菜碱和EGCG对杜长大猪胴体指标的影响
由表5可知,3组试验猪的背最长肌pH、大理石纹评分、肉色评分和肉色亮度、红度、黄度值及肌苷酸含量均无显著差异(P>0.05)。与对照组相比,试验Ⅰ、Ⅱ组的背最长肌24 h滴水损失均显著降低(P<0.05),试验Ⅰ组降低了32.04%,试验Ⅱ组降低了27.07%;试验Ⅰ、Ⅱ组之间背最长肌24 h滴水损失无显著差异(P>0.05)。与对照组相比,试验Ⅰ组的背最长肌肌内脂肪含量显著升高(P<0.05),提高了31.91%;试验Ⅱ组的背最长肌肌内脂肪含量无显著差异(P>0.05)。
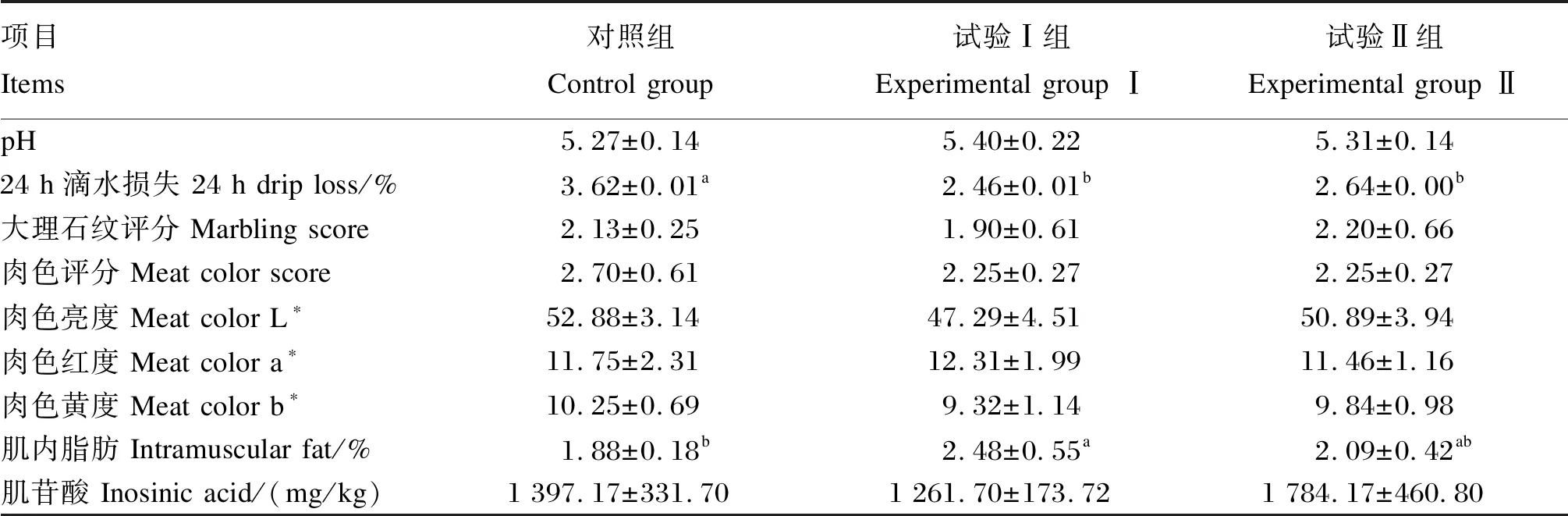
表5 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌肉质指标的影响
2.2.2 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌氨基酸和脂肪酸组成的影响
由表6可知,3组试验猪的背最长肌氨基酸含量均无显著差异(P>0.05)。

表6 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌氨基酸组成的影响
由表7可知,试验共检测出15种脂肪酸,其中包括5种饱和脂肪酸和10种不饱和脂肪酸,其中单不饱和脂肪酸5种,多不饱和脂肪酸5种。与对照组相比,试验Ⅰ、Ⅱ组的背最长肌二十二碳酸含量均显著降低(P<0.05),试验Ⅰ组降低了46.15%,试验Ⅱ组降低了51.92%;试验Ⅰ、Ⅱ组之间背最长肌二十二碳酸含量无显著差异(P>0.05)。3组试验猪的背最长肌其他脂肪酸含量均无显著差异(P>0.05)。

表7 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌脂肪酸组成的影响
2.2.3 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌中肌纤维类型和脂肪沉积相关基因表达的影响
如图1所示,与对照组相比,试验Ⅰ组的背最长肌肌球蛋白重链Ⅱb(MyHCⅡb)的mRNA相对表达水平显著降低(P<0.05),背最长肌肌球蛋白重链Ⅱx(MyHCⅡx)的mRNA相对表达水平显著升高(P<0.05);试验Ⅱ组的背最长肌肌球蛋白重链Ⅱa(MyHCⅡa)和MyHCⅡx的mRNA相对表达水平显著升高(P<0.05)。
如图2所示,与对照组相比,试验Ⅰ组的背最长肌固醇调控元件结合蛋白1(SERBP1)的mRNA相对表达水平显著提高(P<0.05),试验Ⅱ组的背最长肌过氧化物酶体增殖物激活受体α(PPARα)的mRNA相对表达水平显著提高(P<0.05)。
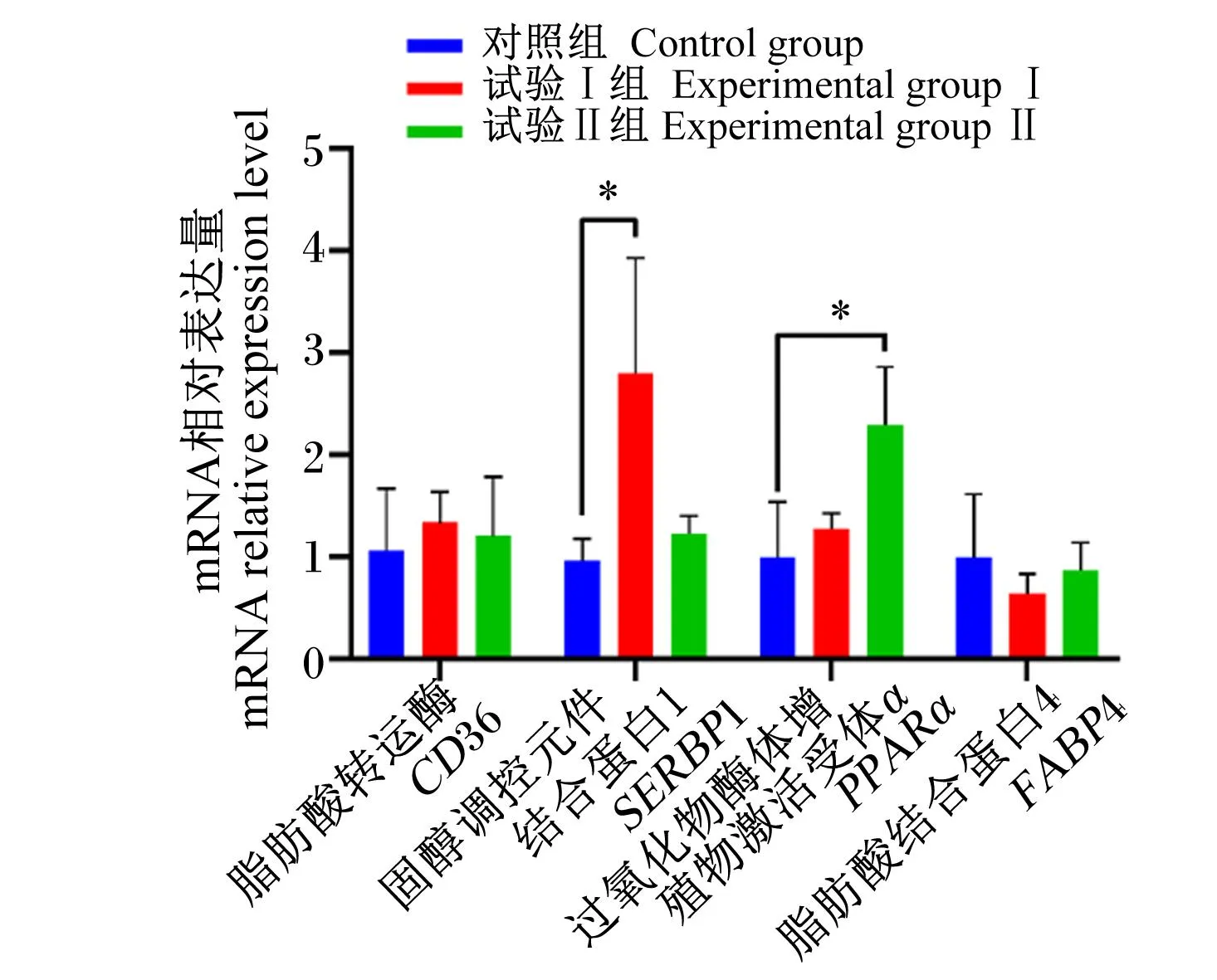
图2 饲粮中添加甜菜碱和EGCG对杜长大猪背最长肌脂肪沉积相关基因表达的影响
2.3 饲粮中添加甜菜碱和EGCG对杜长大猪血清生化和抗氧化指标的影响
由表8可知,血清生化指标方面,与对照组相比,试验Ⅰ、Ⅱ组的血清总胆固醇含量显著降低(P<0.05),试验Ⅰ组降低了8.6%,试验Ⅱ组降低了11.89%;试验Ⅰ、Ⅱ组之间血清总胆固醇含量无显著差异(P>0.05)。与对照组相比,试验Ⅰ组的血清甘油三酯含量显著降低(P<0.05),降低了8%。3组试验猪的血清高密度脂蛋白胆固醇和低密度脂蛋白胆固醇含量均无显著差异(P>0.05)。血清抗氧化指标方面,与对照组相比,试验Ⅰ组的血清谷胱甘肽过氧化物酶活性显著提高(P<0.05),提高了8.52%;试验Ⅱ组的血清丙二醛含量显著降低(P<0.05),降低了6.42%;试验Ⅱ组的血清谷胱甘肽过氧化物酶活性和总抗氧化能力显著提高(P<0.05),分别提高了13.3%和3.4%。

表8 饲粮中添加甜菜碱和EGCG对杜长大猪血清生化和抗氧化指标的影响
3 讨 论
本试验的重点是分析甜菜碱和EGCG的联合应用对育肥期杜长大猪背膘厚和肌内脂肪沉积的影响。甜菜碱作为一种无毒无害、性质稳定的天然化合物,具有提高猪生长性能、降低胴体脂肪含量和改善肉品质等多种功能,是一种“新型营养再分配剂”[13]。而EGCG作为茶多酚在降低动物脂肪沉积方面也非常有效[9]。本试验中,饲粮中添加甜菜碱或甜菜碱和EGCG均能显著降低杜长大猪的背膘厚,虽然试验Ⅰ、Ⅱ组之间背膘厚无显著差异,但甜菜碱和EGCG联合添加的降背膘厚效果优于甜菜碱单独添加。肉色、pH、滴水损失和肌内脂肪含量等指标一般用于评估猪肉品质。本试验中,饲粮中添加甜菜碱或甜菜碱和EGCG均能显著降低杜长大猪的24 h滴水损失,且饲粮中添加甜菜碱还能显著增加肌内脂肪含量,但甜菜碱和ECGG联合添加对肌内脂肪含量无显著影响。以上结果表明,饲粮中添加甜菜碱和EGCG能通过降低背膘和滴水损失改善猪肉品质,且单独甜菜碱还能提高肌内脂肪含量,进一步提升猪肉品质。同时提示甜菜碱和EGCG的联合添加时,EGCG会削弱甜菜碱对肌内脂肪沉积的促进作用,可能是因为EGCG具有良好的降低脂肪沉积作用。
进一步的分子生物学分析结果显示,饲粮中添加甜菜碱显著上调了杜长大猪背最长肌SREBP1的表达,而甜菜碱和EGCG的联合添加显著上调了背最长肌PPARα的表达。SREBP1和PPARα是调节肌内脂肪合成的关键转录因子[14]。甜菜碱作为一种甲基供体,能通过DNA甲基化对基因表达进行表观遗传调控[15]。在鸡原代肝细胞和肝脏的研究中发现,甜菜碱处理能降低SREBP1启动子的甲基化水平并促进SREBP1启动子与糖皮质激素受体(GR)结合增加,进而促进脂质合成[16]。此外,甜菜碱也能通过降低PPARα启动子的甲基化促进其基因表达,从而增加机体的脂肪酸摄取和利用[17]。由此推测,在猪的背最长肌中,甜菜碱可能降低SREBP1和PPARα启动子的甲基化,并促进两者基因表达,进而增加肌肉内脂肪酸摄取和脂质合成来提高肌内脂肪含量。本课题组的前期研究还发现,甜菜碱通过上调肝脏BHMT表达提高机体循环及骨骼肌NADPH含量,进而上调FTO表达,从而降低肌内脂肪细胞的mRNA m6A水平,促进肌内脂肪细胞克隆增殖和分化聚酯,最终提高肌内脂肪含量[7]。以上研究均支持本试验的研究结果,即甜菜碱能促进肌内脂肪沉积。
此外,肌纤维类型是影响猪肉品质的重要因素,在母猪的饲粮中添加甜菜碱,能增加母猪背最长肌的眼肌面积和MyHCⅡx肌纤维的含量[18]。王丽娜等[19]研究发现,在杜长大猪的饲粮中添加0.025%、0.050%的甜菜碱能显著下调MyHCⅠ的表达,同时上调MyHCⅡa、MyHCⅡx和MyHCⅡb的表达。本试验结果显示,饲粮中添加甜菜碱能提高育肥期杜长大猪背最长肌中氧化型肌纤维MyHCⅡa的mRNA相对表达水平,同时显著降低糖酵解型肌纤维MyHCⅡb的mRNA相对表达水平;而饲粮中添加甜菜碱和EGCG能显著提高背最长肌中氧化型肌纤维MyHCⅡa和中间型肌纤维MyHCⅡa的mRNA相对表达水平,甜菜碱和EGCG联合添加对背最长肌MyHCⅡa的mRNA相对表达水平的提高效果优于甜菜碱单独添加。在小鼠成肌细胞系C2C12的研究中发现,甜菜碱能通过胰岛素样生长因子-1(IGF-1)信号通路激活IGF-1受体,从而增加肌球蛋白重链(MyHC)蛋白合成,促进肌纤维的增殖和分化[20]。甜菜碱处理还能通过增加单磷酸腺苷活化蛋白激酶(AMPK)的磷酸化和过氧化物酶体增殖物激活受体γ共激活因子-1α(PGC-1α)的蛋白表达来促进MyHC蛋白合成[21];而EGCG能通过降低AMPK活性和PGC-1α表达减少肌球蛋白重链Ⅰ(MyHCⅠ)形成和线粒体生物合成[22]。在本试验中,甜菜碱和EGCG的联合添加没有影响背最长肌MyHCⅠ的mRNA相对表达水平,可能是因为在本试验的处理条件下,甜菜碱发挥的作用更多,后续试验中应组合不同水平的甜菜碱和EGCG,可能发挥更好的改善肌纤维组成作用。以上结果表明,饲粮中添加甜菜碱或添加甜菜碱和EGCG均能改善育肥期杜长大猪肌纤维组成,且甜菜碱和EGCG联合添加对肌纤维组成的改善效果优于甜菜碱单独添加。
育肥猪生长缓慢,主要是脂肪沉积慢,测定育肥猪血清脂质相关指标可分析育肥猪机体的脂质代谢和脂肪沉积能力。本课题组前期研究发现,在育肥期金华猪的饲粮中添加0.3%的EGCG能显著降低血清甘油三酯含量[11]。杜海涛[23]研究发现,在大体重育肥猪饲粮中添加0.3%的甜菜碱对血清脂质指标无显著影响。本试验结果显示,饲粮中添加甜菜碱显著降低了育肥期杜长大猪血清甘油三酯和总胆固醇含量,而饲粮中添加甜菜碱和EGCG只显著降低了血清总胆固醇含量,表明饲粮种添加甜菜碱能有效提升育肥猪的脂质代谢能力,但甜菜碱和EGCG的联合添加并未表现更好的改善效果,与EGCG的前期研究结果不完全相符,可能与本试验中甜菜碱或EGCG的添加水平有关,其中的具体原因可能还需要进一步分析。在本研究中,饲粮中添加甜菜碱能有效降低血清中的脂质含量,并且提升肌肉中的脂质含量,表明甜菜碱可能具有影响胆固醇分配的作用。已有的研究结果也显示,在育肥猪饲粮中添加甜菜碱可显著增加肌肉中游离脂肪酸(FFA)含量,并降低血清总胆固醇和高密度脂蛋白胆固醇含量,提高肌肉总胆固醇含量[24]。进一步的机制研究发现,甜菜碱可能通过上调脂肪酸转运蛋白的基因表达促进肌肉脂肪酸的摄取;此外,甜菜碱激活了AMPK并上调了与脂肪酸氧化相关的基因,包括PPARα和肉碱棕榈酰转移酶1(CPT1)。因此,甜菜碱可能通过改善肌肉中的脂肪酸代谢来增加肌内脂肪含量,并降低血清中的脂质含量。
本试验还分析了甜菜碱和EGCG的联合添加对育肥猪生长性能和血清抗氧化指标的影响。甜菜碱和EGCG均能有效提高畜禽生长性能。在断奶仔猪上的研究发现,饲粮中添加600 mg/kg甜菜碱能显著提高断奶仔猪的平均日采食量、平均日增重[25]。郭建凤等[26]在“杜洛克×烟台合成系”商品猪上的研究发现,每100 kg基础饲粮中添加50、100 g甜菜碱均能有效促进商品猪的生长。本课题组前期研究发现,饲粮中添加0.3%的EGCG能显著降低金华猪的平均日采食量和料重比[11]。在本试验中,饲粮中添加甜菜碱或甜菜碱和EGCG均能显著降低育肥期杜长大猪的平均日采食量,说明甜菜碱能改善育肥期杜长大猪的饲料转化率,但甜菜碱和EGCG的联合添加并不会进一步改善杜长大猪的生长性能。研究发现,饲粮中添加甜菜碱能降低猪的热量消耗并增加回肠干物质和总肠道粗蛋白质消化率[27]。在本试验中,甜菜碱可能就是通过提高杜长大猪的肠道营养消化率、减少热量消耗从而提高杜长大猪的饲料转化率。
在生猪养殖过程中,养殖环境的改变等多种因素可诱导猪机体内产生大量自由基,进而引起氧化损伤,导致增重减缓、采食量下降等不良反应。现有研究表明,EGCG具有特殊的分子结构,可与自由基进行反应进而清除自由基[28],且甜菜碱也能通过减少丙二醛含量有效减少氧自由基[29]。本试验中,饲粮中添加甜菜碱能显著提高育肥期杜长大猪血清谷胱甘肽过氧化物酶活性;而饲粮中添加甜菜碱和EGCG能显著降低血清丙二醛含量,提高血清谷胱甘肽过氧化物酶活性和总抗氧能力。这表明单独添加甜菜碱能改善杜长大猪的抗氧化能力,且甜菜碱和EGCG的联合添加能进一步提高杜长大猪的抗氧化能力,提示我们在改善育肥猪氧化应激的研究中,可以采取多种营养素联合添加的形式以达到更好的效果。
4 结 论
饲粮中添加甜菜碱或甜菜碱和EGCG能提高育肥期杜长大猪的饲料利用率,通过降低背膘厚和滴水损失改善猪肉品质,并提升杜长大猪的脂质代谢和抗氧化能力;此外,饲粮中添加甜菜碱还能提高杜长大猪的肌内脂肪含量。
