Fe(II)EDTA 络合吸收耦合H2-MBfR 处理NO 效能
2024-03-28刘婉婷钱飞跃赵俊杰徐正慧王建芳缪润珠苏州科技大学环境科学与工程学院江苏苏州25009城市生活污水资源化利用技术国家地方联合工程实验室江苏高校水处理技术与材料协同创新中心江苏苏州25009苏州科技大学天平学院江苏苏州25009
刘婉婷,钱飞跃,2,赵俊杰,徐正慧,王建芳,2,*,缪润珠(.苏州科技大学环境科学与工程学院,江苏 苏州 25009;2.城市生活污水资源化利用技术国家地方联合工程实验室,江苏高校水处理技术与材料协同创新中心,江苏 苏州 25009;.苏州科技大学天平学院,江苏 苏州 25009)
大气中的氮氧化物(NOx)主要来源于各种运输活动及工业过程,尤其是燃煤发电厂或锅炉的工业烟气[1].NO 是NOx的主要成分,在烟气中约占90%[2].近些年来,选择性催化还原(SCR),选择性非催化还原(SNCR),吸附和吸收等方法被陆续用于NOx的去除[3-5],然而这些控制策略存在高成本、低效率等弊端.NOx生物去除技术运营成本低、能源低以及产生的二次污染少.化学吸收-生物还原(CABR)使用亚铁乙二胺四乙酸酯(Fe(II)EDTA)作为溶剂快速吸收NO,利用反硝化细菌将NOx还原为无害的N2的同时,实现Fe(II)EDTA 的再生[6].由于工业烟气通常含有3%~8%的O2,会导致Fe(II)EDTA很容易被氧化成Fe(III)EDTA,进而使NO 无法与之发生络合反应,严重影响处理效率.该技术的关键过程是Fe(II)EDTA的再生,涉及Fe(III)EDTA 和Fe(II)EDTA-NO 向Fe(II)EDTA 的生物转化.
CABR 虽然性能稳定且操作简便,但仍存在气液传质效率低,生物膜堵塞等问题[7].此外,Fe(II)EDTA 再生缓慢会导致NO 去除率降低[8],且Fe(III)EDTA 和Fe(II)EDTA-NO 还原分别需要反硝化和铁还原微生物.氢基质膜生物膜反应器(H2-MBfR)利用清洁的H2作为电子供体,采用无泡曝气的方式,提高了气体传质效率及H2利用率[9].H2-MBfR 的无泡曝气技术和络合剂Fe(II)EDTA 的使用,可有效改善NO 气液传质效率低的问题[10].一体化络合吸收- H2-MBfR 的快速启动及维持反应器内Fe(II)EDTA 浓度是增强处理效果的关键因素[11],关于以H2为电子供体的混合功能微生物培养研究较少.目前研究人员利用膜改性方法改变中空纤维膜丝的表面特性[12-13]缩短启动时间;添加还原性金属[14-15];利用生物膜电极反应器[16-17];混合吸收剂(Fe(II)Cit/Fe(II)EDTA)[18]等加速Fe(II)EDTA 再生.然而,这些方法的局限性也相对明显,例如加剧膜污染,能耗和成本偏高,可持续性较差.本研究构建Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统,通过启动H2-MBfR,驯化具有同步反硝化及铁还原性能混合菌种,探究在H2-MBfR 体系中,H2作为唯一电子供体的反硝化和铁还原效能以及NO的去除效率,研究进水Fe(II)EDTA 浓度、pH 值对一体化系统中NO 去除效率的影响,分析微生物群落结构特征.
1 材料与方法
1.1 实验装置
本实验使用双管式MBfR,分别命名为A 管和B管,A 管通入H2(氢气发生器QL-300 7350,山东赛克赛斯),B 管通入NO(河南源正),通过蠕动泵(Longer,China)促进反应器内液体循环.反应器总体积为80mL,有效容积60mL,反应器内径1.2cm,高度36cm.实验用膜丝(830Y-0.20,日本杜邦帝人)单组含有32根PVDF 中空纤维膜,膜内径250µm,外径350µm,膜丝表面积为13.5cm2,具体装置如图1 所示.
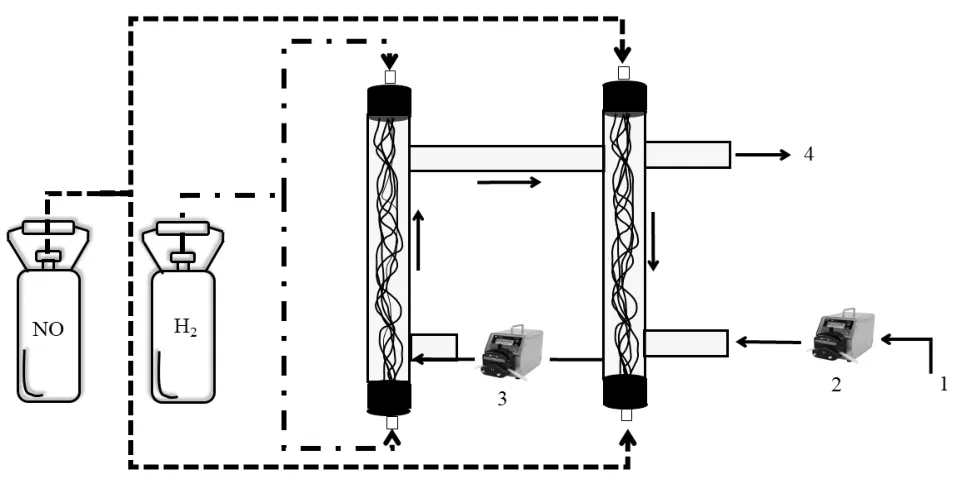
图1 H2-MBfR 反应器装置图Fig.1 Diagram of H2-MBfR reactor installation
1.2 实验水质
采用实验室人工模拟废水,由NaNO3,KH2PO4,K2HPO4及微量元素组成,NaHCO3作为唯一无机碳源,投加量20mg/L.Fe(III)EDTA 由FeCl3·6H2O,乙二胺四乙酸四钠按比例配置而成.
为维持微生物生长,在模拟废水中投加微量元素:1.98mg/L MnCl2·4H2O,0.5mg/L CuSO4·5H2O,0.44mg/L Na2MoO4·2H2O,0.38mg/L NiCl2·6H2O,0.028mg/L H3BO3,0.2mg/L ZnSO4·7H2O,0.002mg/L CaCl2·2H2O,0.1mg/L MgCl2.
1.3 实验试剂
NaNO3(≥99.0%);NiCl2·6H2O(≥98.0%);ZnSO4·7 H2O(≥99.5%)及CaCl2·2H2O(≥74.0%)均购买自上海凌峰化学试剂有限公司.KH2PO4(≥99.5%);K2HPO4(≥99.0%);FeCl3·6H2O(≥99.0%);乙二胺四乙酸四钠(≥99.0%);MnCl2·4H2O(≥99.0%);CuSO4·5H2O(≥99.0%);Na2MoO4·2H2O(≥99.0%);MgCl2(≥99.0%)均购买自天津市科密欧化学试剂有限公司.NaHCO3(≥99.5%);H3BO3(≥99.0%)均购买自无锡市晶科化工有限公司.所有药品的纯度均为AR.NO 浓度为400×10-6(3.01%,河南源正).
1.4 分析项目及检测方法
1.4.1 水样测试指标及方法 本实验通过无菌注射器取样,水样经过0.45µm 水系PES 滤膜过滤.硝态氮采用紫外分光光度法HJ/T 346-2007 测定;亚硝态氮采用N-(1-奈基)-乙二胺分光光度法测定;pH 值采用pH 计(BestLab 962244)测定;NO 采用烟气分析仪(testo 340)测定;Fe(II)EDTA 采用1,10-菲罗啉分光光度法测定[19];Fe(II)EDTA-NO 采用分光光度法进行测定[20];总铁浓度采用邻菲罗啉分光光度法测定[21].
1.4.2 生物膜样品的采集与16S rRNA 高通量测序分析 取同步反硝化铁还原阶段第27d 膜丝表面的生物膜样,命名为DQ;一体化系统第50d 膜丝表面的生物膜样,其中与溶液相接触的生物膜样,命名为DS,A 管供氢侧与中空纤维膜接触的生物膜样,命名为DH,B 管通NO 侧与中空纤维膜接触的生物膜样,命名为DN,共4 个生物膜样品.
4 组生物膜样品送至美吉公司进行16S rRNA高通量测序分析. 16S rRNA 基因在V3~V4 高变区测序,选用细菌通用引物 338F(5’-ACTCCTACGGGAGGCAGCAG-3’)及806R(5’-GGACTACHVGGGTWTCTAAT-3’).在优化序列的基础上按照97%相似性分析,得到每个OTU 对应的物种分类信息,并在各水平上统计每个样品的群落组成,获得样品中微生物门(phylum)和属(genus)等物种组成及丰 度.
1.5 数据处理
1.6 实验方法
1.6.1 反应器运行 本实验主要分为2 个阶段:菌种驯化及生物膜培养(S1)、构建Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统(S2).其中菌种驯化阶段(S1)分为两个部分:驯化具有氢自养反硝化能力的菌种及生物膜培养(S1-SR),驯化具有同步反硝化及铁还原能力的混合菌种(S1-SF).
反应器接种来自苏州某污水厂厌氧污泥,接种污泥浓度为5000mg/L,S1 阶段在A 管和B 管的PVDF 膜内通入H2,氢分压维持在0.06MPa.S2 阶段A 管通入H2,氢分压为0.06MPa,B 管通入400×10-6的NO,分压维持在0.06MPa.本实验控制进水pH 值为6.8~7.0,HRT 设置为8h.内循环泵流速维持在72mL/min 以保障A 管和B 管基质与生物膜良好接触.保证反应器内氢通量JH2 为0.096g H2/(m2·d),NO通量 JNO为0.117g NO/(m2·d).
S1-SR 阶段:为了更好地挂膜,初期采用间歇式运行反应器,HRT 为48h.运行2d 后,HRT 缩短至24h,间歇运行3d,PVDF 膜上附着一层生物膜,进一步缩短HRT 至8h,运行3d,反应器转为连续运行,并逐渐提高进水硝酸盐的浓度.直到 NO3--N 提高至5mmol/L,硝酸盐去除率稳定,则认为已完成氢自养反硝化微生物驯化及效能提升.之后实验维持HRT为8h 不变.
S1-SF 阶段:在H2-MBfR 具有稳定反硝化性能的基础上,在反应器内加入不同浓度的Fe(III)EDTA,培养具有同步反硝化及铁还原能力的生物膜,探究同步反硝化与铁还原的性能.
S2 阶段:在具有良好的同步铁还原和反硝化性能的H2-MBfR 的基础上,调整进水浓度及配比.通过系统还原Fe(III)EDTA 产生的Fe(II)EDTA 络合NO,逐渐降低硝酸盐基质浓度,构建Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统,探究络合吸收、反硝化与铁还原以及整体系统对NO 的去除能力.
1.6.2 批次实验 在具有良好性能的Fe(II)EDTA络合吸收NO 耦合H2-MBfR 还原一体化系统的基础上,控制pH 值在6.8~7.0,探究初始Fe(II)EDTA 浓度(1,3,5,10mmol/L)对系统的影响.保持初始Fe(II)EDTA 浓度为5mmol/L,HRT 为8h,通气压力0.06MPa,探究pH 值(6,7,8,9)对系统的影响.

表1 实验条件Table 1 Experimental conditions
2 结果与讨论
2.1 氢自养反硝化菌种驯化(S1-SR 阶段)
在图1 所示反应器内,双管都通入H2,进行氢自养反硝化菌种的驯化及生物膜的培养.考虑到后期一体化系统的构建,在保证充足电子供体的前提下,提升硝酸盐浓度至5mmol/L.满足在单管供氢的工况下,进水硝酸盐能够被完全消耗,确保氢自养反硝化菌种的活性.
由图2 可以看出,氢自养反硝化菌逐渐适应进水基质,随着进水硝酸盐浓度从1mmol/L 提升到5mmol/L,硝态氮去除率整体呈现先快速提升,后维持平稳的趋势.在硝酸盐浓度达到5mmol/L 时(R5 阶段),硝态氮去除率维持在98.93%以上.H2-MBfR 能够在短时间内启动且具有较高的脱氮效率[22].值得注意的是当硝酸盐浓度由1mmol/L 提升到2mmol/L时,硝态氮去除率出现下降趋势,在第10d 的时候出现最低去除率,为84.06%,这主要是因为在驯化初期,反硝化菌性能不稳定.而当进水硝酸盐浓度超过2mmol/L 后,硝态氮去除率波动较小,表明微生物逐步适应环境及进水基质,表现出稳定的氢自养反硝化性能,此阶段反应器完成挂膜.

图2 H2-MBfR 反硝化启动及效能变化Fig.2 Denitrification initiation and efficiency changes in H2-MBfR
在第32d 于反应器内进行原位批次实验,进水硝酸盐浓度为2mmol/L.由图3 可以看出,随着时间的增加,硝态氮去除率逐渐增加.60min 后,反应速率的变化较小.在180min 时,硝态氮去除率达到100%,说明此时反应系统内的硝酸盐已经被微生物全部消耗,进一步验证了此时反应器内驯化完全的微生物具有高效的反硝化性能,能够满足后续实验需求.硝酸盐去除速率随着时间的增加呈现先增加后减小的趋势,在 30min 时获得最大去除速率为0.38mmol/(L·min).说明反应器内驯化完全的微生物,其反硝化效能达到稳定.
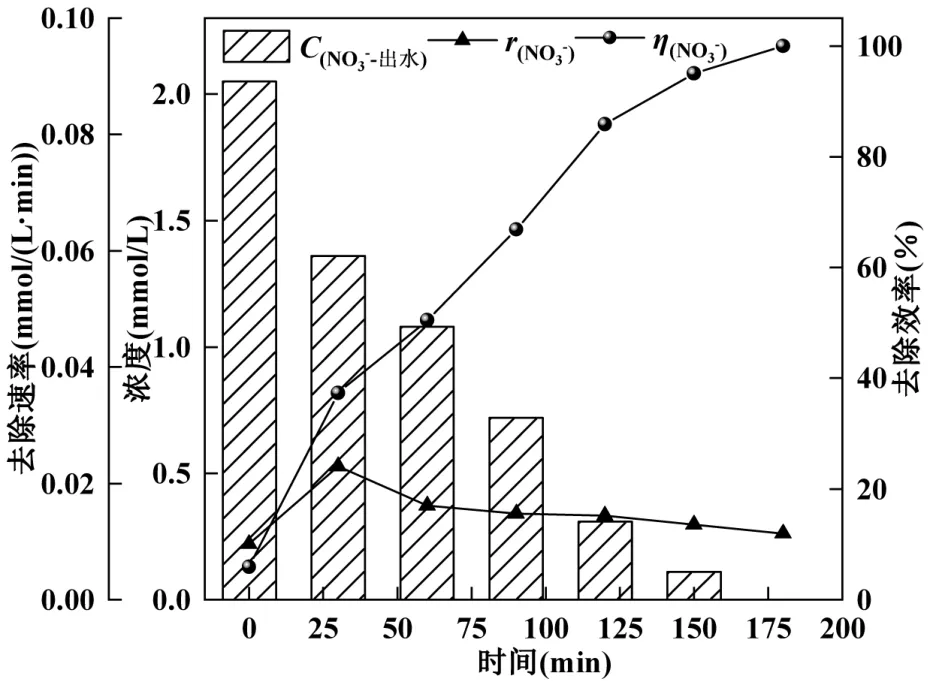
图3 第32d H2-MBfR 反硝化批次实验Fig.3 Day 32d enitrification batch experiments in H2-MBfR
2.2 驯化具有同步反硝化及铁还原能力的混合菌种(S1-SF 阶段)
改变进水工况,驯化具有同步反硝化及铁还原能力的混合菌种,如图4 所示.F1 阶段(1~12d)设置进水Fe(III)EDTA 为5mmol/L,NO3--N 为1mmol/L.运行过程中发现,混合体系中硝酸盐优先还原,当监测到出水中有Fe(II)EDTA 时,硝酸盐及亚硝酸盐浓度基本消耗至0mg/L.硝酸盐在反应器中作为第一电子受体,能够优先利用H2进行还原[23].
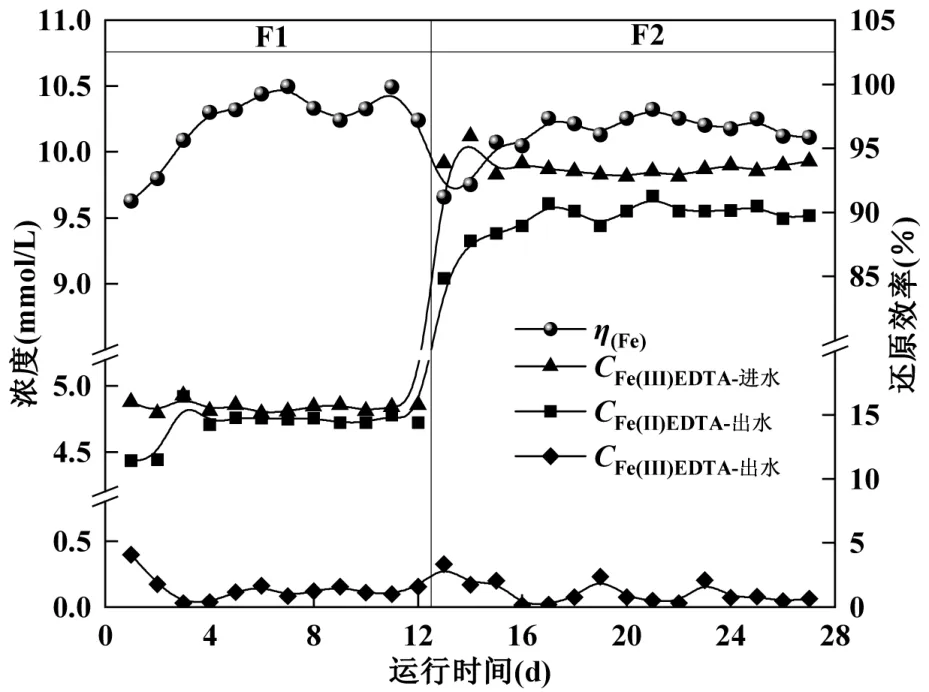
图4 H2-MBfR 同步反硝化及铁还原效能Fig.4 Simultaneous denitrification and iron reduction efficiency in H2-MBfR
F1 阶段,在反应第1d 出水Fe(III)EDTA 的浓度快速降至0.39mmol/L,还原率达到90.87%,表明部分反硝化菌具有一定的铁还原性能,因此能够在工况改变的初期保持较高的铁还原率.在2~12d之间,出水Fe(III)EDTA 浓度稳定在0.20mmol/L 以下,铁还原率最大值出现在第7d,为99.84%,此阶段微生物已经适应环境内Fe(III)EDTA 浓度.在第13d,将进水Fe(III)EDTA 浓度提高至10mmol/L,同时进水硝酸盐浓度提升至2mmol/L(F2 阶段).此时出水 Fe(III)EDTA 出现小幅度提升,出水中Fe(II)EDTA 浓度有所下降,铁还原率也降至91.19%,随后出水Fe(II)EDTA 开始逐步提升,最终达到稳定.在 21d 时,铁还原率出现最大值为98.06%,这也表明0.06MPa 的氢分压可以提供充足的电子供体.
以上结果表明,H2-MBfR 通过快速富集反硝化菌,实现同步反硝化和铁还原的功能.当实验进行28d 后,铁还原效率维持高水平,因此认为同步反硝化和铁还原混合菌种驯化及生物膜挂膜完成.
2.3 Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原体系对NO 的去除效能(S2 阶段)
直接在体系内通入NO,监测一体化体系对NO的去除率,逐步利用Fe(II)EDTA-NO 替代硝酸盐.由图5 可知,在A1 阶段(1~10d)连续通入400×10-6的NO,为保证反硝化微生物在耦合初期还能维持活性,进水Fe(III)EDTA 为10mmol/L,进水NO3--N 为2mmol/L.运行的第1d,出水Fe(II)EDTA 浓度维持在9.04mmol/L 以上,NO 去除率为90.87%.随着运行时间的增加,出水Fe(II)EDTA 浓度及NO 去除率逐渐增加.在第10d,出水Fe(II)EDTA 浓度及NO 去除率分别为9.61mmol/L 和99.50%.说明Fe(II)EDTA 络合吸收耦合H2-MBfR 还原体系能够做到对NO 的高效且稳定的去除.
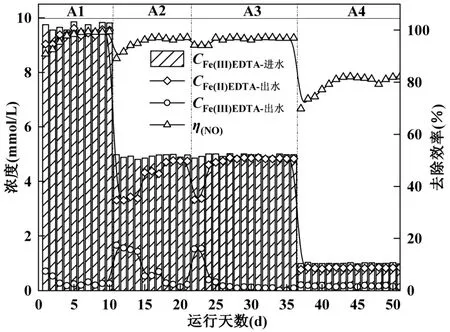
图5 Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统脱氮效能Fig.5 The removal efficiency of NO by Fe(II) EDTA complexing absorption coupled with H2-MBfR reduction integrated system
A2 阶段(11~21d)降低进水 Fe(III)EDTA 至5mmol/L,进水NO3--N 降低至1mmol/L,考察进水Fe(III)EDTA 浓度对NO 去除效率的影响.改变运行工况初期,出水Fe(II)EDTA 浓度及NO 去除率发生明显波动,出水Fe(III)EDTA 浓度也有所提升.此时Fe(II)EDTA 生成量最小值为3.32mmol/L,NO 去除率最小值为89.11%.考虑是由于进水Fe(III)EDTA浓度下降,导致生成的Fe(II)EDTA 浓度降低,发生短暂波动.在此工况下运行15d 后,微生物逐步适应Fe(III)EDTA 浓度降低,出水Fe(II)EDTA 浓度明显回升,同时NO 去除率也开始稳步提升.出水Fe(II)EDTA 最大值为 4.76mmol/L,NO 去除率最高为97.24%.
A3 阶段(22~36d),不再投加硝酸盐,进水Fe(III)EDTA 浓度维持5mmol/L 不变.出水Fe(II)EDTA略有下降,下降至3.32mmol/L,NO去除率下降至94.20%.此时生成的Fe(II)EDTA 浓度与工况改变前的浓度存在差距,但与第11d 的NO 去除率明显下降不同,第22d 的NO 去除率只降低了2.98%.考虑是由于A2 阶段不仅减少了硝酸盐投加量,同时进水Fe(III)EDTA 浓度的降低导致了络合剂的Fe(II)EDTA 生成量减少.体系内Fe(II)EDTA 浓度的降低直接导致了NO 去除率的明显减低.A3 阶段保持进水Fe(III)EDTA 浓度不变且不再投加硝酸盐.Li等研究证明,反硝化菌能够有效还原Fe(II)EDTANO,因此在前期减少硝酸盐投加量,促使反硝化菌逐步适应,提高对Fe(II)EDTA-NO 的还原效率[24].而在之后的批次实验中发现,6h 内Fe(II)EDTA-NO 浓度快速下降,证实了前期论述,此时反硝化菌能够高效还原Fe(II)EDTA-NO,加快Fe(II)EDTA 的循环再生,达到NO 的高效去除.由此可以推断,此时改变系统的运行工况,不易影响 NO 去除率.维持出水Fe(II)EDTA 稳定后,NO 去除率也随之稳定,说明在电子供体供应充足的条件下,本系统能够实现稳定高效去除NO.
A4 阶段(37~51d),进一步降低进水Fe(III)EDTA浓度至1mmol/L,考察低进水Fe(III)EDTA 浓度下的影响.第37d,出水Fe(II)EDTA 浓度为0.77mmol/L,NO 去除率明显下降至69.85%.随着反应时间的增加,NO 去除率最大值为82.21%.出气端NO 浓度提升,说明NO 去除不完全,可能是体系内络合吸收剂Fe(II)EDTA 不足造成.进水Fe(III)EDTA 的减少,导致生成的Fe(II)EDTA 不足,能够吸收的NO 总量下降,最终导致NO 去除率的下降.Zhao 等研究表明,Fe(II)EDTA 浓度是吸收 NO 的关键,体系内高Fe(II)EDTA 浓度能够加快NO 的去除[17].反之,低Fe(II)EDTA 浓度会导致NO 去除量减少,验证了A4段的实验结果.
综上所述,保证体系内具有一定浓度的Fe(II)EDTA,是保证Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统稳定、高效去除NO 的关键因素之一.
2.4 各类体系中Fe(II)EDTA-NO 还原效率比较
如表2 所示,生物膜电极反应器对Fe(II)EDTANO 去除率仅在79.1%[17].而Xia 等虽然对反应器进行了优化,提升了Fe(II)EDTA-NO 的去除率,但远没有一体化体系更为稳定高效[25].利用还原剂[26-28]、金属铁粉[29]及锌粉[15]对Fe(II)EDTA-NO 进行还原再生,都存在去除效率较低的问题,而金属粉末易氧化失效,从而丧失循环利用的能力.本研究采用Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统,可以更为稳定高效的去除 NO,实现Fe(II)EDTA 快速循环再生.

表2 不同体系中Fe(II)EDTA-NO 还原对比Table 2 Comparison of Fe(II)EDTA-NO reduction in different systems
2.5 影响因素
2.5.1 进水Fe(II)EDTA 浓度对NO 去除率的影响由图6(a)可以看出,在60min内基本实现Fe(II)EDTA消耗完全.由图6(b)可知,体系中Fe(II)EDTA-NO 浓度持续增加.随着初始Fe(II)EDTA 浓度的增加,60min 时体系中Fe(II)EDTA-NO 浓度也随之提升,说明初始Fe(II)EDTA 浓度越高,能够吸收的NO 总量越大,有利于体系对NO的快速去除.由图6(c)可以看出,初始Fe(II)EDTA 浓度越高,体系内的NO 平均去除速率越高.在Fe(II)EDTA 浓度为10mmol/L 时,达到最大去除速率为 44.68mg/(m3·h). 初始Fe(II)EDTA 浓度5mmol/L 和10mmol/L 时,其NO 平均去除速率差别较小,认为在此NO 浓度及进气流速限制下,5mmol/L 的Fe(II)EDTA 即能达到良好的NO 去除效果,因此增加初始Fe(II)EDTA 浓度,NO 的平均去除速率未出现提升[31].
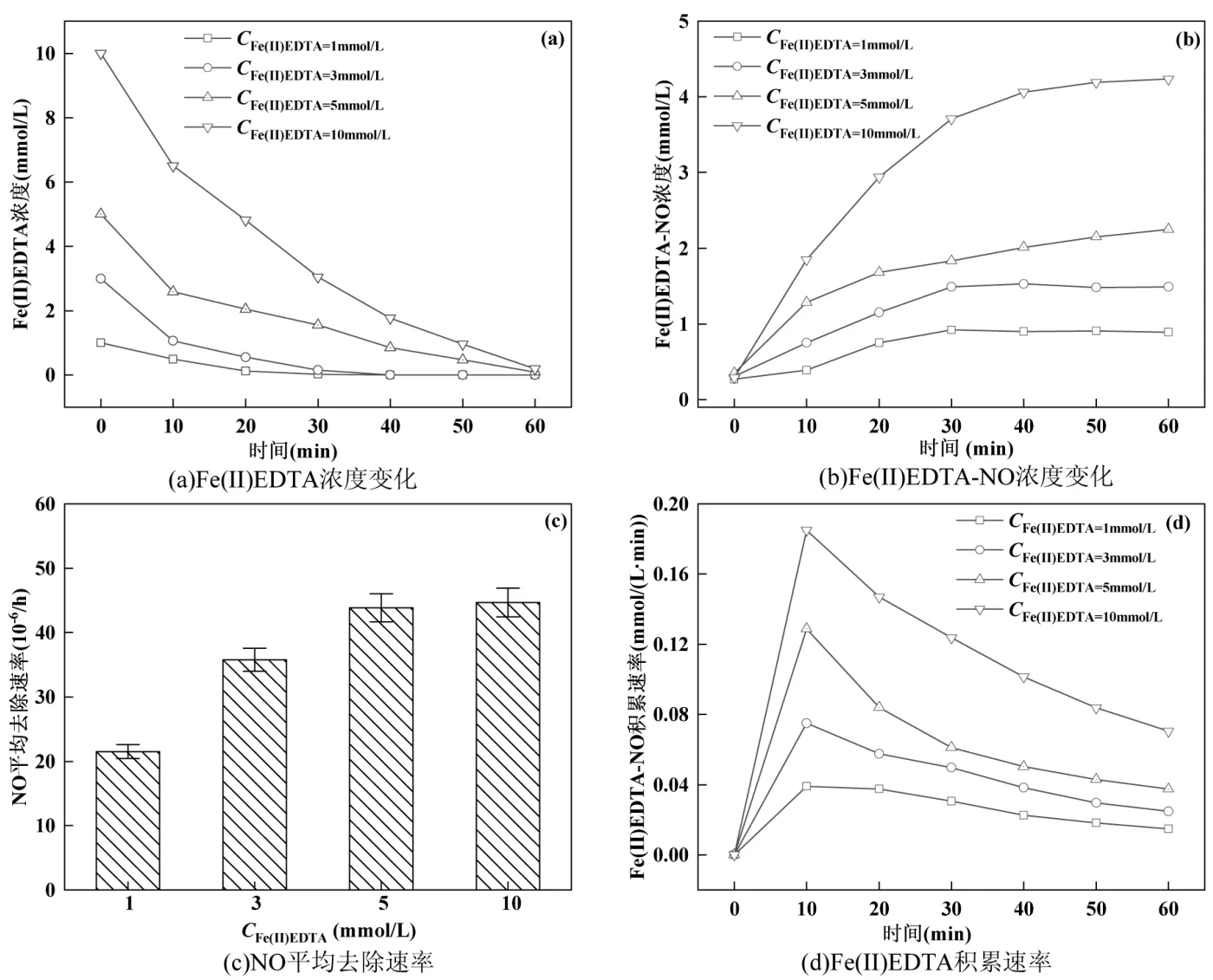
图6 初始Fe(II)EDTA 浓度对NO 去除的影响Fig.6 Effect of initial Fe(II)EDTA concentration on NO removal
在图6(d)中,Fe(II)EDTA-NO 积累速率出现先上升再下降的趋势,且Fe(II)EDTA 浓度越高,生成速率越高,Fe(II)EDTA-NO 在初期运行的第10min内快速积累,积累速率分别为0.039,0.075,0.129 和0.185mmol/(L·min),此时吸收速率高于生物还原速率.随着反应时间延长,Fe(II)EDTA-NO 浓度下降,表明高浓度的初始Fe(II)EDTA 有利于NO 的快速络合.
体系中 Fe(II)EDTA-NO 最终浓度与初始Fe(II)EDTA 浓度之间存在差异,认为是由于反应器内生成 Fe(II)EDTA-NO 的同时,也发生着部分Fe(II)EDTA-NO 的还原.测定的出水Fe(II)EDTANO 浓度为体系内的积累量,即生成量与消耗量的差值,因此会与初始Fe(II)EDTA 浓度存在差异.
2.5.2 进水pH 值对NO 去除效率的影响 由图7(a)可以看出,pH 值越高,相同反应时间体系内剩余Fe(II)EDTA 浓度越高,说明 pH 值能够影响Fe(II)EDTA 对NO 的吸收能力.图7(b)可以看出,pH值越高,体系中Fe(II)EDTA-NO 浓度越小.当pH 值为 6 时,Fe(II)EDTA-NO 的即时浓度最高,达4.24mmol/L.随着pH 值的提升,体系中Fe(II)EDTANO 浓度也随之降低.pH 值为9 时,Fe(II)EDTA-NO浓度为 0.89mmol/L,表明 pH 值过高不利于Fe(II)EDTA 对NO 进行络合,符合前期分析.由图7(c)可以看出,随着pH 值的升高,NO 的平均去除速率逐渐下降.当pH 值为6 时,NO 平均去除速率明显高于其他三组,为43.86mg/(m3·h).当pH值提高至9时,NO平均去除速率明显下降,仅为pH 值为6 时的50%左右,为23.66mg/(m3·h).
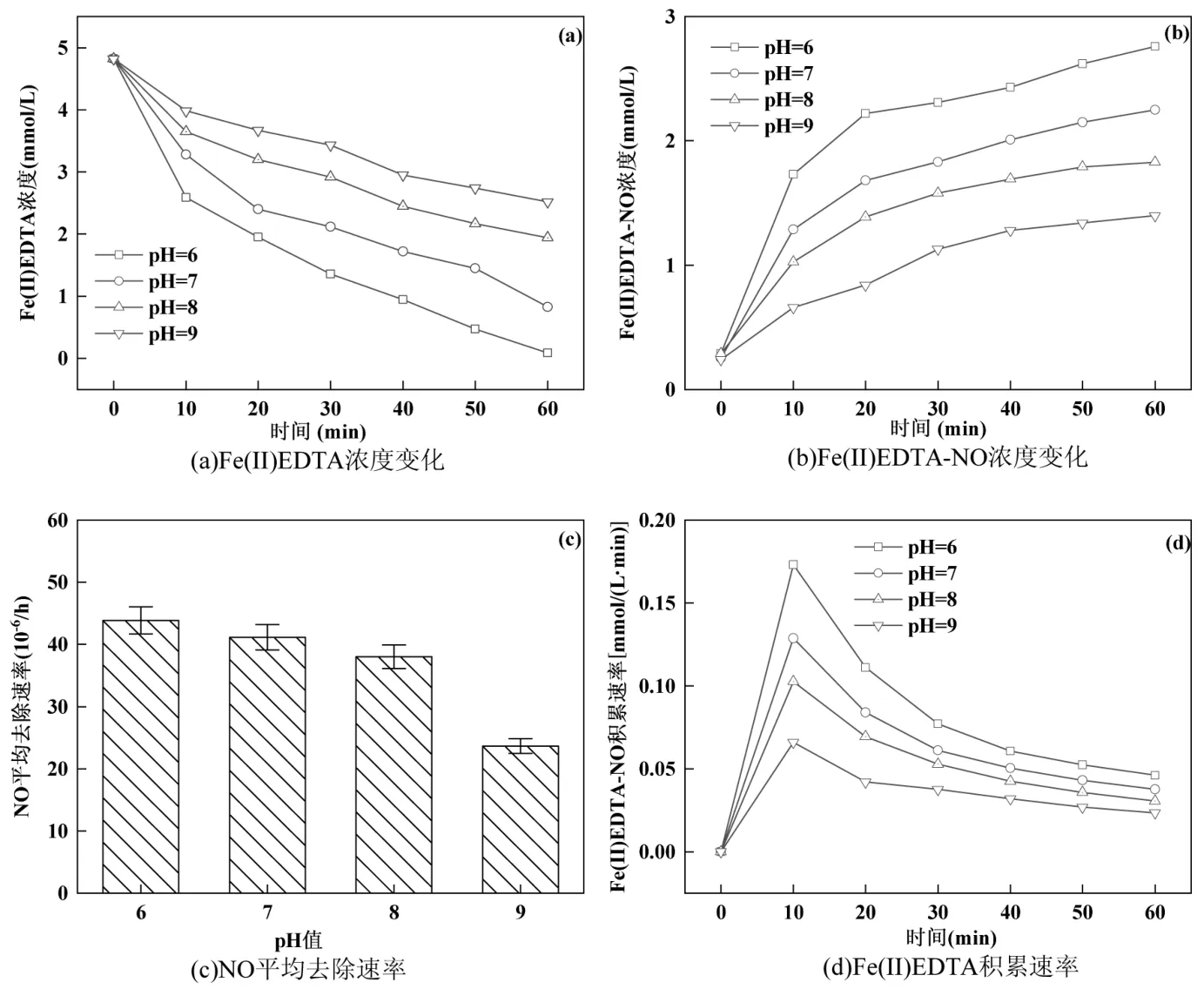
图7 进水pH 值对NO 去除的影响Fig.7 Effect of influent pH on NO removal
根据实验结果分析可知,Fe(II)EDTA 充足情况下,与NO 的络合能力同样影响着一体化反应体系对NO 的去除.Fe(II)在偏酸性的环境下更容易与EDTA 形成螯合物[32].殷祥男等研究发现 pH>8时,Fe(II)EDTA 对NO 的络合能力对比pH 值在6~8时出现下降,NO 的吸收速率较低[33].pH 值过高时Fe(II)EDTA 的稳定性降低,游离的Fe2+会与溶液中的OH-生成Fe(OH)2沉淀,从而失去对NO 的络合吸收能力[34].当系统内环境呈碱性时,会造成EDTA 的降解[35],降低参与反应的Fe(II)EDTA 浓度,从而影响对NO 的去除能力.且H2溶于水,以H+作为电子供体被利用,因此更适应偏中性的环境[36].
研究表明,pH>8.4 时,反硝化关键酶的活性会受到影响[33],进水pH 值过高会导致微生物还原能力受到抑制.由图7(d)可知,随着pH 值的提升,体系中Fe(II)EDTA-NO 积累速率逐渐降低,进水pH 值为9时,最高Fe(II)EDTA-NO积累速率远低于进水pH值为6 时的积累速率.
pH 值控制为6 时,系统快速生成Fe(II)EDTANO 的同时,也被微生物快速还原,实现Fe(II)EDTA的高速循环再生.在此条件下,Fe(II)EDTA 与Fe(II)EDTA-NO 维持稳定的平衡.过高的pH 值会严重影响微生物活性及络合吸收剂Fe(II)EDTA 的吸收效率,从而影响NO 的去除.将pH 值调控在6 左右,有利于一体化系统高效去除NO.
2.6 微生物群落分析
2.6.1 微生物群落多样性分析 由表3 可知,4 个样品中的微生物测序样本覆盖率较高,均大于0.99,所测微生物多样性数据具有说服力.Simpson 指数值排序为:DH>DN>DS>DQ,说明不同位置微生物群落多样性存在细微差距,与H2接触近的组分物种丰度较高,但没有明显差异.Chao 指数能够有效估计物种数量,从Chao 指数分析,DS、DH、DN 无明显差异,但远高于同步反硝化及铁还原阶段.这是由于通入NO后,体系内反硝化及铁还原的反应更为复杂,需要更多的微生物参与到还原过程中,具有硝酸盐还原功能的Dechlorosoma 及具有铁还原功能的Geothrix菌属仅在DS、DH、DN 样品属水平中得以发现.梁岑研究发现,Fe(II)EDTA-NO 浓度的升高会导致总的微生物物种多样性会出现上升趋势[37].DS、DH、DN 阶段的Shannon 指数对比同步反硝化铁还原阶段下降,这是由于NO具有毒性,靠近NO进气端的微生物受到影响, 后期发现 Lutispora 、Chitinophagaceae、Anaerovorax 等参与氮循环,具有硝酸盐降解功能的部分菌属被淘汰.同时NO 与体系中Fe(II)EDTA 结合,使得反硝化及铁还原代谢途径进一步增强,反硝化菌及铁还原菌属在生存方面的优势增大,部分不适应环境的微生物,例如Rhodocyclaceae、Aquimonas 即被淘汰,优势菌种占比上升.Sobs 指数,即生态优势度指数,数值大小排序为DS>DN>DH>DQ,说明微生物群落内物种数量分布不均匀,优势菌种占比大,符合预期.
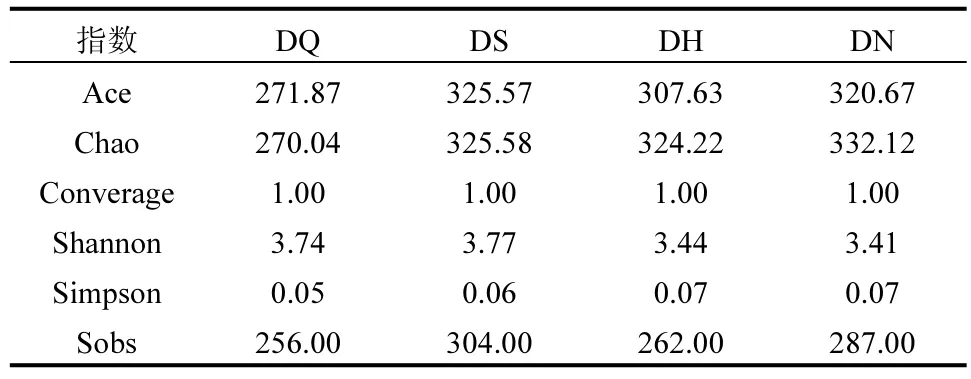
表3 微生物群落α-多样性分析Table 3 Microbial community α-diversity analysis
2.6.2 门水平微生物群落结构分析 由图8 可知,变形菌门Proteobacteria、拟杆菌门Bateroidota、厚壁菌门Frimicutes 为同步反硝化及铁还原驯化阶段的优势菌种,分别占比44.1%,17.11%,33.76%.变形菌门Proteobacteria 中包含亚硝化细菌、硝化细菌和反硝化类的细菌,利用进水中的氮元素进行反应活动,促进微生物生长,在调节微生物群落结构方面有重要影响,是体系中的优势菌种,相对丰度较高[38].拟杆菌门Bateroidota 异养菌,参与氮磷的去除,分解进水中的分子聚合物重新组成溶解性有机物[39].厚壁菌门Firmicutes 是典型铁还原富集物中的常见菌属,即铁还原富集菌群的优势菌门[40].以上几类菌种的出现,结合此阶段的硝酸盐及亚硝酸盐被完全消耗,铁还原率稳定在98.60%以上,可以从微生物水平表明同步反硝化及铁还原阶段菌群驯化完成,具备反硝化能力和铁还原能力的微生物具有高丰度,确保了进水硝酸盐及Fe(III)EDTA 能够被快速消耗还原.

图8 同步反硝化及铁还原阶段门水平微生物Fig.8 Microorganisms at the phylum level in the simultaneous denitrification and iron reduction stage
由图9 可知,在一体化阶段,优势菌种同前一阶段相同,但占比出现明显变化.DS 及DN 阶段变形菌门Proteobacteria 及拟杆菌门Bacteroidetes 相对丰度明显增加.DH 段变形菌门Proteobacteria、拟杆菌门Bateroidota、厚壁菌门 Frimicutes 分别占比50.35%,16.43%,14.98%,说明持续通入NO 可以促进反硝化菌种的富集,此阶段最低NO 去除率也可达81.20%,由此证明了Fe(II)EDTA-NO 在一体化阶段中占比较高时,促进了Proteobacteria 和Bacteroidetes菌种大量繁殖积累.而DN段由于靠近NO侧,厚壁菌门Frimicutes 相对丰度有所降低,说明具有铁还原功能的菌属对NO 敏感,不适宜在NO 浓度较高的环境中生存.对比之下,DH 段由于靠近供氢端,厚壁菌门Frimicutes 相对丰度对比其他两组最高,根据前期分析可知, Fe(III)EDTA 与硝酸盐共存时,微生物优先还原硝酸盐,然后进行铁还原.而反硝化过程消耗了大量的电子供体,因此只有在电子供体充足的条件下,铁还原功能菌属才能大量繁殖[41].DH 阶段厚壁菌门 Frimicutes 相对丰度较高,有利于强化Fe(III)EDTA 还原过程[42],此时出水Fe(III)EDTA 还原率在78.66%以上.
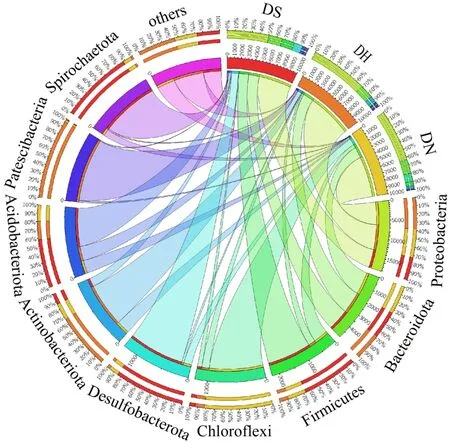
图9 一体化阶段门水平微生物Fig.9 Microorganisms at the phylum level in the integration stage
而与同步反硝化及铁还原阶段相比,变形菌门Proteobacteria 及拟杆菌门Bateroidota,相对丰度有所上升,而厚壁菌门Frimicutes 物种相对丰度下降.而根据前期微生物群落多样性分析可知,一体化阶段三组样本的微生物数量,都远高于同步反硝化及铁还原阶段,说明一体化阶段更有利于微生物繁殖富集,典型菌种的大量出现也确保了系统对NO 的高效去除,这一微生物特征与NO 去除效能相匹配.
2.6.3 一体化系统进化树分析 由图10 可以看出,DS、DH、DN 阶段具有相同的菌种,但其丰度有较大差异.nirK 型反硝化菌Rhizobiales 属于变形菌门Proteobacteria,在DS、DH、DN 阶段相对丰度分别为 26.48%,4.81%,9.63%.Hydrogenophaga 属于Proteobacteria,是典型的厌氧氢自养微生物,利用H2作为电子供体参与硝酸盐的代谢.在DH 阶段相对丰度远超DS 和DN 阶段,对生物膜的形成起到重要作用[43],一体化阶段稳定高效的NO 去除率也充分证明了在充足H2供应下,对电子供体敏感的微生物更易富集,获得更好的处理效果.Desulfitobacterium作为稳定的优势菌种,属于Frimicutes,是一种严格厌氧菌,在土壤、污泥中广泛存在,不仅可以利用电子供体还原硝酸盐,同时具有铁还原能力[44].此时出水Fe(II)EDTA 浓度与进水Fe(III)EDTA 浓度差值最大仅为0.19mmol/L,说明Desulfitobacterium 作为属水平的铁还原功能关键菌属,其较高的相对丰度确保了在一体化阶段对Fe(III)EDTA 的还原能力,保障了体系的稳定性.Desulfitobacterium 菌属的出现也验证了系统的厌氧状态.
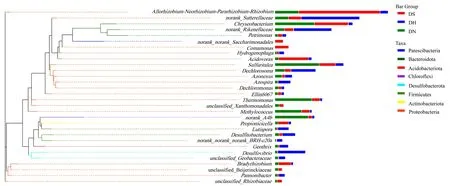
图10 一体化阶段属水平进化树Fig.10 Phylogenetic tree at the genus level in the integration stage
在一体化阶段,兼性厌氧菌Dechloromanas 为优势菌种, 是典型的硝酸盐还原菌.DH 段Dechloromanas 丰度明显大于DS、DN 及同步反硝化铁还原阶段,说明菌种易富集在靠近供氢端的位置,能够获得充足的电子供体参与硝酸盐代谢活动.Geothrix 是少数可以利用Fe(III)氧化物进行呼吸的淡水可培养细菌之一[45].DH 段Geothrix 相对丰度明显大于其余两组,说明Geothrix 对H2的需求较大,在H2浓度相对较高的膜表面,Geothrix 容易富集,符合前期推论.上述典型菌属的出现,从微生物水平说明了Fe(II)EDTA 的稳定生成与循环,解释了一体化阶段的高效NO 去除率.
通过对一体化阶段3 组样本的进化树分析可以发现,除上述分析的菌属,Bacteroidetes、Sulfuritalea、Sutterellaceae 和Thermomonas 等皆属于变形菌门Proteobacteria,且相对丰度较高.根据Sun 研究可知,以上皆为典型的反硝化优势均属[46],说明变形菌门Proteobacteria 在一体化系统中较为重要,承担重要功能,其在微生物水平上的高相对丰度确保了一体化阶段的反硝化功能.在此阶段,最高NO 去除率为97.24%,符合前期分析.
3 结论
3.1 H2-MBfR 采用先培养稳定的反硝化生物膜,再驯化铁还原菌的方法,可快速成功构建Fe(II)EDTA络合吸收NO 耦合H2-MBfR 还原体系系统,NO 最高去除率为99.50%.
3.2 提高初始Fe(II)EDTA 浓度,有利于提升NO 吸收量,体系内NO 平均去除速率随着Fe(II)EDTA 浓度的提升而提升.在Fe(II)EDTA 浓度为10mmol/L时,最大去除速率为44.68mg/(m3·h).控制进水pH 值为6,NO 平均去除速率可达43.86mg/(m3·h),有利于一体化系统络合吸收和生物还原的良好平衡.
3.3 同步反硝化铁还原阶段与一体化阶段优势菌门均为Proteobacteria、Bacteroidetes 和Firmicutes,丰度占比有差异,分别为44.10%,17.11%,33.76%和50.35%,16.43%,14.98%,两阶段都具有稳定的反硝化及铁还原效能.
3.4 Proteobacteria 是Fe(II)EDTA 络合吸收NO 耦合H2-MBfR 还原一体化系统中主导菌门,在各生物膜样品中丰度均达到50%以上.Hydrogenophaga 是典型的氢自养微生物,具备氢自养反硝化能力,属于变形菌门Proteobacteria.
