高寒区氮添加和间作种植互作对燕麦和豌豆根系构型影响的研究
2024-03-22鲍根生李媛冯晓云张鹏孟思宇
鲍根生,李媛,冯晓云,张鹏,孟思宇
(1. 青海省畜牧兽医科学院,青海 西宁 810016;2. 青海省青藏高原优良牧草种质资源利用重点实验室,青海 西宁 810016;3. 青海大学畜牧兽医科学院,青海 西宁 810003)
干旱和寒冷极端逆境、草地载畜量过高和人类不合理利用等因素的综合作用,造成高寒地区天然草地生产力较低,成为限制高寒区草地畜牧业发展的重要瓶颈[1-2]。然而,平衡草食家畜与草地低生产力间的矛盾成为解决这一瓶颈的关键途径[3-4]。前期研究发现,人工草地生产力和牧草营养价值远高于天然草地[5-6];同时,碳固持、养分循环利用及水土保持方面人工草地也凸显优势[7-8]。禾本科和豆科植物混播(同行混合间作)是高寒地区人工草地增产提质的经典种植模式[9],该种植方式主要利用混播物种对空间和营养资源的互补利用特征,即深根系禾本科植物对土壤氮素的强消耗特性能强化短根系豆科植物的固氮能力[10]。禾本科和豆科植物间作种植可分为同行混合、隔行和带状间作3 种方式[11-12];其中,隔行和带状间作广泛应用于农业生态系统,能显著提高间作作物的产量和品质[13-15]。相比之下,高寒区人工草地常采用禾豆同行混合间作种植方式,混合间作系统的产量和品质较单播均有显著增加[16]。众多研究证明,隔行间作种植的禾本科和豆科种间兼容性和间作系统生产力更高[17]。然而,以往研究仅关注禾豆单播与同行混合间作或禾豆单播与隔行间作种植对系统生产力提升的对比研究[14,17-18],而系统生产力增加往往与禾豆间作植物根系空间分布差异及资源互补利用有关。
根系作为植物吸收、运输土壤养分和水分的关键载体,成为植物与土壤相互作用的桥梁[19]。研究表明,间作作物对资源利用的生态位差异是禾豆间作系统产量增加的关键途径,而这一过程主要与间作作物根系对土壤养分和水分获取能力的空间差异性有关[20-21]。例如,禾本科植物主根纤细较长且细根发达能生长至深层土壤进而吸收利用较多的养分,而豆科植物主根粗壮较短且细根较少仅能吸收较浅土层的养分[22]。可见,间作作物根系形态特征很大程度上会影响间作作物对土壤养分和水分的利用效率[23]。根系构型主要通过根系的几何形态、分形特征和拓扑结构等参数直观反映植物根系在土壤中的空间布局、拓展能力和资源捕获能力[24-25]。例如:朱亚琼等[17]研究发现不同混播方式对禾豆混播草地植物根系构型特征具有明显的可塑性,异行混播通过增加细根数量和拓展根系表面积途径降低无芒雀麦(Bromus inermis)和红豆草(Onobrychis viciaefolia)种间竞争强度,进而实现异行混播体系生产力增加。同时,朱亚琼等[26]研究发现混播豆科牧草根系形态参数比单播豆科牧草显著增加。可见,禾豆间作种植可通过优化间作群体的根系构型和高效利用土壤营养资源等途径降低间作作物的种间竞争强度,进而提高间作体系系统生产力[11,21,26]。
氮作为影响陆地生态系统植物生产力的重要限制因子,氮添加能短期内提高土壤肥力和保障作物丰产[27]。禾豆间作体系利用豆科固氮和禾本科耗氮的种间氮素利用特性,已成为全球广泛采用提高作物产量和降低氮肥投入的农业生产体系[10]。研究发现,禾豆间作体系生产力提高主要与间作植物根系对土壤氮素的协同吸收利用过程有关,而这一过程与间作植物根系构型变化、生理特性及根际微生物组成密切相关[21]。Ramirez-Garcia 等[28]和Corre-Hellou 等[29]研究发现,氮添加能显著改变间作禾本科和豆科植物的根系形态参数及土壤的空间分布结构。Zhang 等[30]的研究也发现氮添加对禾豆间作体系根系直径、主根和侧根发育有明显调控作用。然而,有关氮添加和间作种植互作对禾豆根系空间分布与构型特性调控作用尚不明晰。基于此,本研究以高寒区人工草地建植常用牧草-燕麦(Avena sativa)和饲用豌豆(Pisum sativum)为研究对象,比较氮添加和不同间作种植方式(单播、同行混合和隔行间作)对燕麦和豌豆生物量(地上和地下)、根系形态及构型的影响,明晰不同间作种植方式和氮添加对禾豆间作体系系统生产力提升的根系调控作用,并为高品质高寒区禾豆间作人工草地科学建植提供技术支撑。
1 材料与方法
1.1 试验地概况
试验地位于青海省西宁市湟中区土门关乡加汝尔村(36°26′51″ N,101°41′28″ E,海拔2668 m),无灌溉条件,属典型浅山种植区。该地属高原大陆性气候,暖季短而冬季较长。年均温和年降水量分别为2.8 ℃和368 mm,平均无霜期120 d。土壤类型为栗钙土,有机质、全氮和全磷含量分别为34.39 g·kg-1、2.20 g·kg-1和2.44 g·kg-1,铵态氮、速效磷和速效钾含量分别为121.00 mg·kg-1、27.60 mg·kg-1和290 mg·kg-1,pH 为8.18。2021 年试验地前茬作物为燕麦。
1.2 试验设计
间作种植材料为燕麦和饲用豌豆,其中,燕麦为青海省畜牧兽医科学院培育的‘青永久872’新品系,该品种具有晚熟、抗逆性强、饲草和籽粒产量双高特性;豌豆品种为‘青建1 号’饲用豌豆,具有产草量高的特点。田间种植采用裂区试验设计,氮添加为主区,不同间作种植方式为裂区。其中,氮添加设置3个水平,分别为对照(0 kg·hm-2)、低氮(50 kg·hm-2)和高氮(200 kg·hm-2);间作种植方式分为燕麦单播、豌豆单播、燕麦与豌豆同行混合间作和燕麦与豌豆隔行间作。田间共设置4个15 m×20 m 区组,依据处理组合(3个氮水平×4 种种植方式),各区组内设置12个2 m×5 m 小区,将不同处理随机布置于区组内。相邻区组间设置宽2 m 保护行,小区间设置1 m保护行。
2022 年5 月5 日,开展田间种植试验。用自制划印器在小区内划出8个长5 m、宽25 cm 的开沟线,沿开沟线用开沟器人工挖出深10 cm 种植行。过磷酸钙(P2O5含量12%)作为底肥一次性施入,每行施入量为12 g。尿素(N 含量46%)按1∶1 比例分别在播种期(底肥)和拔节期(追肥)分两次施入;其中,低氮和高氮添加处理每行尿素施入量分别为6.25 和25.00 g。为避免高温对尿素造成的挥发性损失,田间表层土和尿素按10∶1 比例混合均匀后撒入种植行内,不施氮处理种植行内撒入同体积田间土。依据高寒区燕麦和饲用豌豆播种量的相关研究[31-32],燕麦和豌豆播种量分别为225 和112 kg·hm-2,即单播燕麦和豌豆的行播量分别为28.12 和13.90 g。隔行间作种植时,燕麦和豌豆隔行交替种植4 行,每行播种量与单播播种量相同。同行混合间作种植时,燕麦和豌豆播种量降至单播行播量的50%。间作混合播种时,燕麦和豌豆种子充分混匀后撒播至种植行内。播种后,人工覆土并镇压,燕麦拔节期和开花期种植小区内人工除杂两次。
1.3 生物量和根系形态参数测定
燕麦乳熟期(8 月16 日),各处理小区内随机选取0.5 m×0.5 m 样方进行取样。由于燕麦和豌豆根系主要分布于0~30 cm 土层[33],用铁锹沿样方边缘小心挖取深35 cm 土块。为防止土块内根系受损,土块置于注满水的水桶(高50 cm、直径80 cm)内浸泡24 h,后用自来水缓慢冲洗燕麦和豌豆根际土壤,挑选根系完整且长势一致的燕麦和豌豆整株各10 株带回实验室。将燕麦、豌豆按茎叶和根系分开;其中,5 株燕麦、豌豆茎叶和根系105 ℃杀青30 min,80 ℃烘箱中烘至恒重并测定地上(aboveground biomass, AB)和地下生物量(belowground biomass, BB)。采用双光源彩色扫描仪(中晶 ScanMaker i800 plus,杭州万深检测科技有限公司)对剩余5 株燕麦和豌豆根系进行扫描,待扫描根系需用软毛刷梳理,尽量保证其均匀分布。根系放入扫描盘后,用镊子再次调整根系,尽量使根系均匀分布于扫描盘内。扫描根系图片用LA-S 植物根系分析系统(杭州万深检测科技有限公司)对根形态指标进行分析,获取根表面积(root surface area, RSA)、根体积(root volume, RV)、根系平均直径(root average diameter, RAD)、连接数(connections, Co)、根尖数(root tips, RT)、分叉数(forks, F)、分形维数(fractional dimension, FD)、内部连接(internal connections, Ic)和外部连接(external connections, Ec)等参数。
1.4 根系拓扑结构测定
依据植物根系主根和侧根的土壤空间分布差异,根系拓扑结构可分为鱼尾形和叉状分支两种极端模式,而大多数植物根系结构介于这两种模式之间,拓扑指数可直观反映不同植物根系分支的差异[24-25]。拓扑指数(topological index, TI)= lgA/lgM,其中,M指根系所有外部连接数,外部连接是根系分支与分生组织间的部位;A指最长根系通道所有内部连接数,内部连接指根系两个分支点间部位。TI 接近1,表明根系趋向于鱼尾形分支;而TI 接近0.5,表明根系趋向于叉状分支。
1.5 数据统计分析
采用一般线性模型分析主区(氮添加)、裂区(间作种植方式)及二者交互效应对燕麦和豌豆生物量(地上和地下生物量)、根系形态参数(根表面积、根体积、根系平均直径、连接数、根尖数、分叉数、分形维数、内部连接数和外部连接数)和根系拓扑指数的影响;其中,间作种植方式和氮添加为固定因子,区组为随机因子。同时,选择Duncan 多重比较分析不同间作种植方式和氮添加水平对豌豆和燕麦生物量、根系形态参数和拓扑指数的影响。分析前需进行方差齐性检测以满足方差分析。另外,采用结构方程模型(structural equation modeling, SEM)综合分析氮添加和间作种植方式对燕麦和豌豆根系形态参数和根系生物量的影响。将根系形态参数作为关键因子,不同间作种植方式和氮添加水平作为起始变量,根系拓扑指数、分形维数和地下生物量设为目标变量。选取R 语言中PLS-PM 模块构建分段结构方程。所有数据用“平均值±标准误”表示,采用IBM SPSS 26.0 软件进行数据分析,Sigmaplot 15.0 进行绘图。
2 结果与分析
2.1 氮添加对间作种植方式燕麦和豌豆生物量的影响
氮添加和间作方式对燕麦地上、地下及豌豆地上生物量有显著影响,其中,氮添加和间作方式对燕麦生物量和豌豆地上生物量存在显著交互作用(表1 和图1,P<0.05)。随氮添加量增加,燕麦地上和地下生物量持续增加;高氮添加燕麦的地上和地下生物量显著高于对照和低氮添加处理(图1a,c;P<0.05)。低氮添加后,隔行间作燕麦生物量显著高于单播和同行混合间作;而不添加氮同行混合间作燕麦地上生物量显著高于单播和隔行间作燕麦(图1a,c;P<0.05)。随氮添加量增加,单播豌豆地上和地下生物量显著增加;低氮添加隔行间作豌豆的生物量显著高于对照和高氮添加处理(图1b,d;P<0.05)。高氮添加水平下,单播豌豆地上生物量显著高于同行混合和隔行间作豌豆,而氮添加处理豌豆地下生物量却显著低于单播豌豆(图1b,d;P<0.05)。

图1 氮添加对单播、同行混合和隔行间作种植燕麦和豌豆生物量的影响Fig. 1 Effect of nitrogen addition on above- and belowground biomass of monocropped, mixed and alternate-row intercropped planting patterns for oat and pea
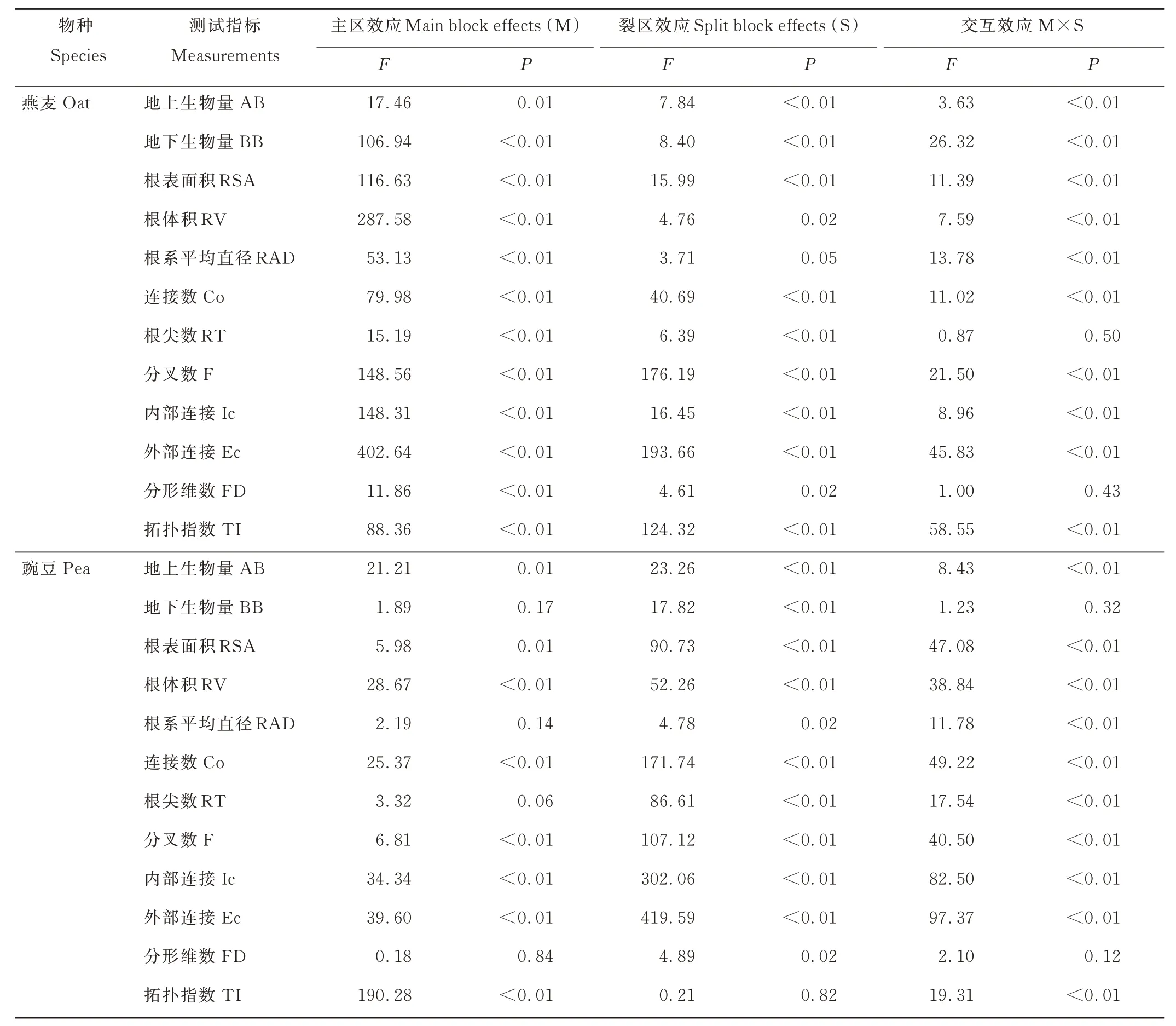
表1 主区(氮添加)和裂区(间作种植方式)对燕麦和豌豆生物量及根系形态影响的方差分析Table 1 ANOVA of above- and belowground biomass and root morphological parameters in monocropped, mixed and alternate-row intercropped planting patterns for oat and pea under different nitrogen levels
2.2 氮添加对间作种植方式燕麦和豌豆根系形态的影响
氮添加和间作种植方式对燕麦和豌豆根系形态存在显著影响(表1,P<0.05);其中,氮添加和间作种植方式除对燕麦根尖数与分形维数无显著交互作用外(表1,P>0.05),对其他根系形态参数均存在极显著的交互作用(表1,P<0.01)。低氮水平下,同行混合间作燕麦的根表面积、根体积、根尖数、分叉数和内部连接数显著高于单播燕麦,而隔行间作燕麦的外部连接数显著高于单播和同行混合间作燕麦(图2,P<0.05)。高氮水平下,隔行间作燕麦除根体积和根尖数外,其他根系形态参数显著高于单播燕麦(图2a,c,e,g,i,k;P<0.05)。随氮添加量增加,单播豌豆根表面积、根体积和根尖数呈先降低后增加的趋势,而分叉数和内部连接数却持续降低(图2 b,d,f,h,j;P<0.05)。同行混合间作豌豆的根体积、根尖数和内部连接数随氮添加量增加呈先增加后降低的趋势,而根表面积和外部连接数却随施氮量增加显著降低(图2 b,d,f,j,l;P<0.05)。隔行间作豌豆的根表面积、根体积和根尖数随施氮量增加呈先增加后降低的趋势(图2b,d,f),而分叉数、内部和外部连接数却显著增加(图2 h,j,l;P<0.05)。
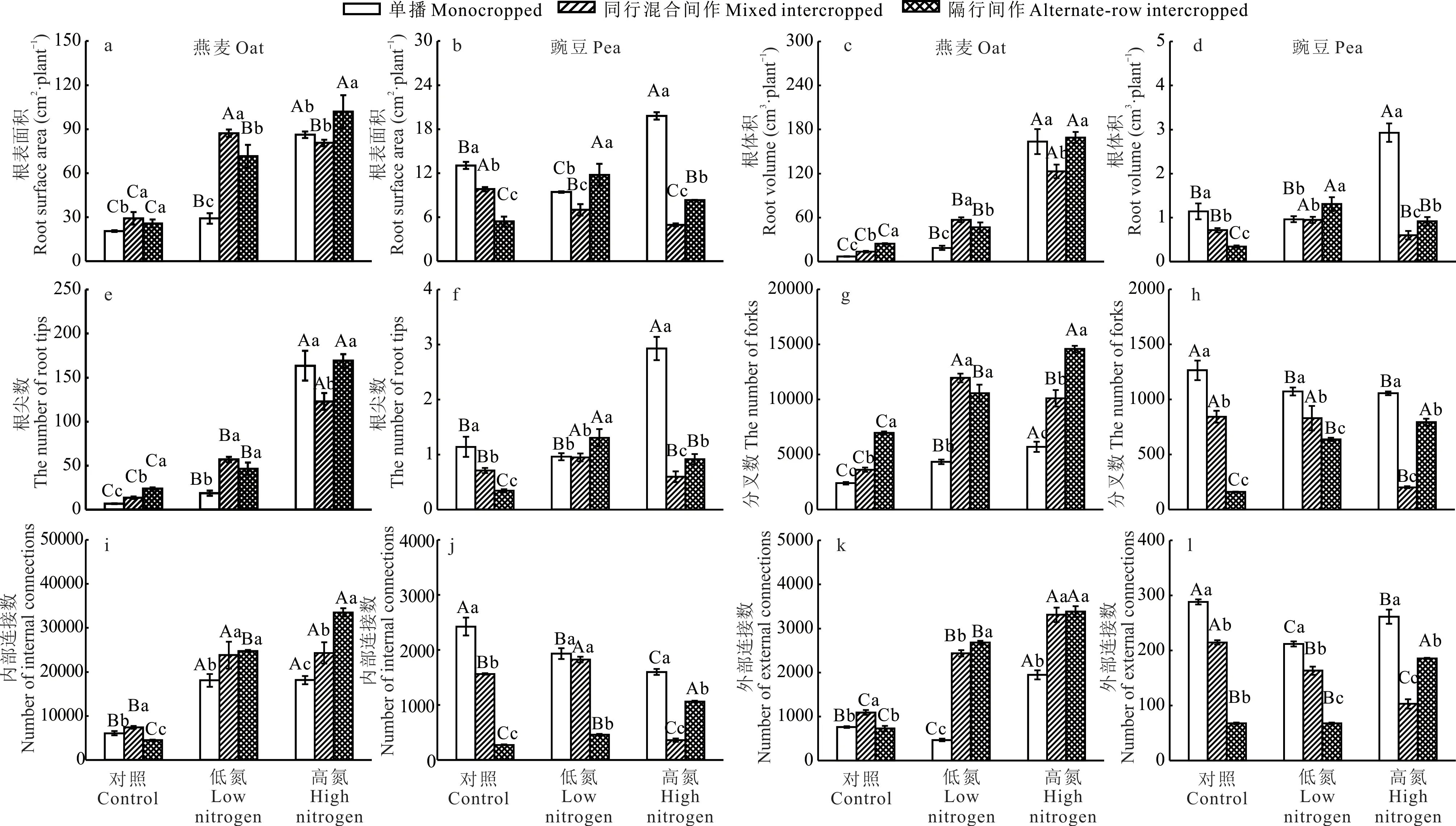
图2 氮添加对单播、同行混合和隔行间作种植燕麦和豌豆根系形态的影响Fig. 2 Effect of nitrogen addition on root morphology in monocropped, mixed and alternate-row intercropped planting patterns for oat and pea
2.3 氮添加对不同间作种植方式燕麦和豌豆根系分形维数和拓扑指数的影响
氮添加和间作种植方式对燕麦根系分形维数和拓扑指数存在显著影响;其中,氮添加和间作种植方式对根系拓扑指数有显著交互作用(表1,P<0.01)。高氮添加同行混合间作燕麦的拓扑指数显著高于未添加和低氮添加燕麦,而低氮添加单播燕麦的拓扑指数显著低于同行混合和隔行间作燕麦(图3a,P<0.05);未添加和低氮添加后,同行混合和隔行间作燕麦的拓扑指数显著高于单播燕麦(图3a,P<0.05)。随氮添加量增加,不同间作种植方式豌豆的拓扑指数不断增加(图3b);其中,未添加氮条件下,隔行间作豌豆的拓扑指数显著高于单播和同行混合间作豌豆;而低氮添加后,单播豌豆的拓扑指数显著高于同行混合和隔行间作豌豆(图3b,P<0.05)。不同间作种植燕麦分形维数随施氮量增加而增加,其中,高氮添加燕麦的分形维数显著高于未添加燕麦(图3c,P<0.05)。未添加氮条件下,单播豌豆的分形维数显著高于同行混合和隔行间作豌豆(图3d,P<0.05)。
2.4 氮添加和不同间作方式与根系形态构型和根系生物量的关联分析
SEM 路径综合分析显示,氮添加、间作种植方式与燕麦、豌豆根系构型和根系生物量间存在相关性(图4)。其中,间作种植方式与燕麦根表面积、连接数及分叉数存在显著正相关,路径系数分别为0.276、0.292 和0.331(图4a);同时,根体积、根系平均直径、连接数和节点数与燕麦分形维数间呈正相关关系,路径系数分别为0.597、0.848、1.455 和1.144(图4a),而根表面积、根尖数、分叉数和外部连接数与燕麦分形维数间呈负相关关系,路径系数分别为-0.776、-0.381、-0.961 和-1.568(图4a)。另外,间作种植方式与燕麦分形维数存在极显著正相关关系,路径系数为0.561(图4a,P<0.001)。氮添加水平与燕麦根表面积、根体积和外部连接数存在显著正相关,而与根系平均直径存在显著负相关关系(图4a,P<0.01)。同时,根体积和内部连接数与拓扑指数存在显著负相关关系,而外部连接数与拓扑指数存在显著正相关关系(图4a,P<0.01)。另外,氮添加与燕麦根系拓扑结构和地下生物量存在极显著正相关,路径系数为0.499 和2.640(图4a,P<0.001)。
间作种植方式与豌豆根体积、连接数、根尖数和内部连接数存在显著负相关关系,路径系数分别为-0.467、-0.612、-0.338 和-0.128(图4b,P<0.01),氮添加与连接数存在显著负相关关系,而与根体积存在显著正相关关系(图4b,P<0.01)。同时,内部连接数与拓扑指数存在极显著负相关关系,而与外部连接数存在极显著正相关关系(图4b,P<0.001)。另外,间作种植方式与豌豆根系拓扑指数存在极显著负相关关系,路径系数为-0.369(图4b,P<0.001),而氮添加与根系生物量存在显著负相关关系,路径系数为-0.619(图4b,P<0.05)。
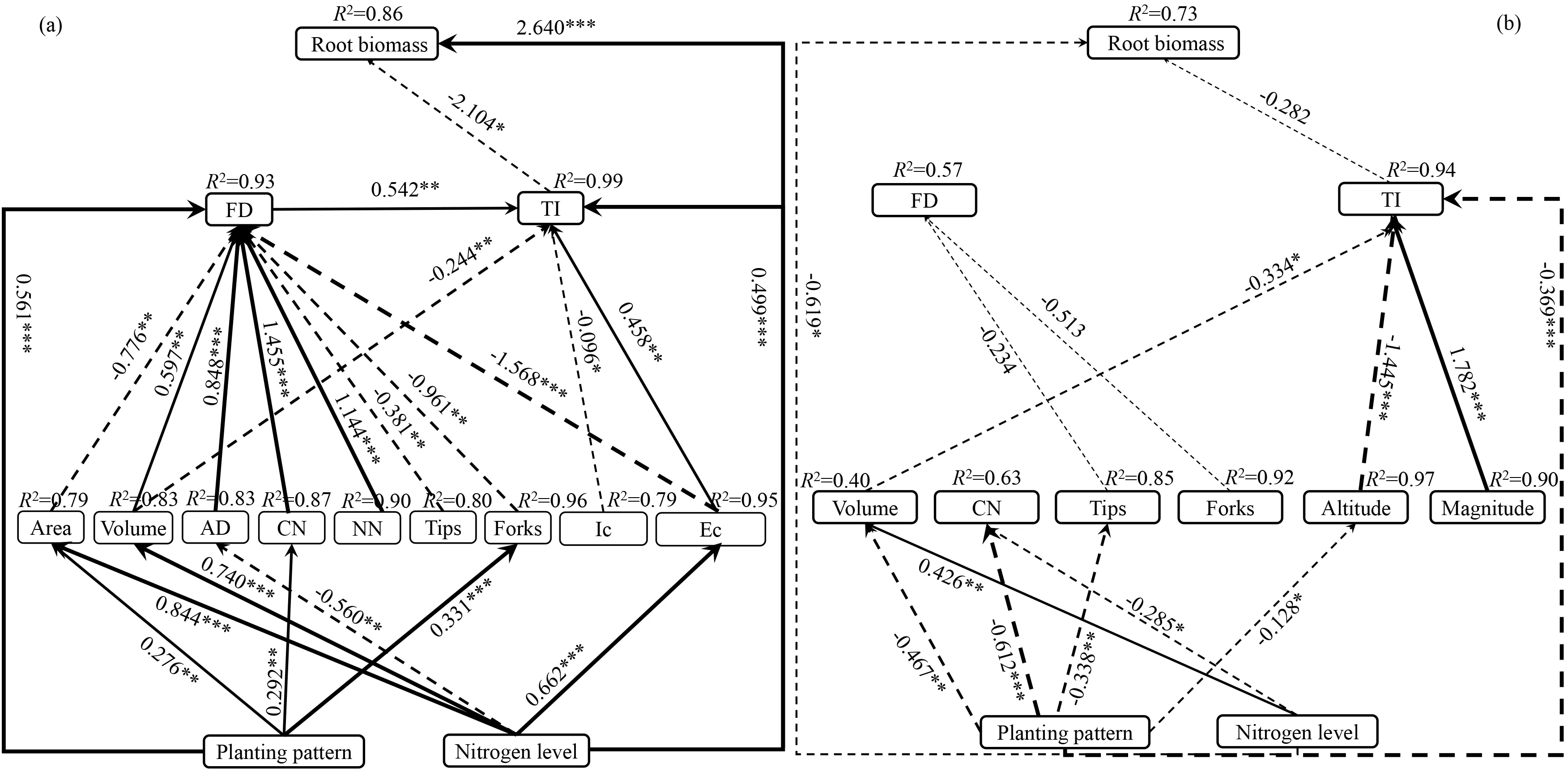
图4 氮添加和不同间作种植方式对燕麦、豌豆根系构型和地下生物量影响的结构方程模型Fig. 4 Structural equation model (SEM) based on effects of nitrogen addition and intercropped planting patterns on root architecture and root biomass for oat and pea
3 讨论
3.1 燕麦和豌豆根系构型对氮添加的响应差异
根系作为植物从土壤中获取养分和水分的重要器官,很大程度上决定茎叶生长和种子产量[34]。然而,土壤矿质养分和水分异质性分布也将影响植物根系发育和空间布局[35];同时,植物常通过改变根系形态、分布和调节生理特性等途径最大限度获取土壤的养分和水分,进而增强植物对干旱和贫瘠生境的适应能力[36]。氮作为植物生长发育所需的基本元素,成为同域植物间竞争的关键养分。根系构型差异在一定程度上决定植物对土壤氮素吸收利用的能力[34-35]。例如,燕麦作为一年生禾草具有根体积大和根系长等特点,这将有利于燕麦从垂直和水平维度上获取土壤氮素,使燕麦具备耐贫瘠特性[37]。相比之下,豌豆由于根际共生细菌的固氮作用能丰盈土壤氮素,导致根体积小、侧根发育迟缓和根系生物量低[38]。本研究中未添加氮条件下燕麦根系形态参数明显高于饲用豌豆的研究结论与其一致。
土壤原位氮添加通过改变土壤氮素异质分布,进而影响植物根系形态及分布。燕麦为喜硝态氮型植物,随尿素(硝态氮)添加量增加,燕麦根直径、根长和侧根密度等形态特征显著变化,而总根长和根直径变化能增加硝态氮吸收和促进燕麦侧根发育[37]。本研究发现氮原位添加后,燕麦根系形态参数总体优于未添加氮处理。然而,原位硝态氮添加后,豌豆根表面积、根体积和根尖数增加而分叉数却降低。说明豌豆在养分充足土壤中可通过增加根系与土壤接触的途径吸收土壤中氮素,同时,根尖作为固氮细菌活动最活跃区域,根尖数量降低表明高硝态氮添加可限制根际固氮细菌固氮能力,这与豆科植物生物固氮属于高耗能过程有关,这也是豆科植物适应不同质量土壤环境的权衡策略[39]。另外,氮添加对拟南芥(Arabidopsis thaliana)根系发育研究也证明原位添加硝态氮能显著降低拟南芥侧根数量而促进侧根生长[30]。由此可见,未添加氮素的燕麦和豌豆根系形态和空间分布格局存在显著差异,这主要与植物根系发育特性、土壤养分异质性分布及物种间土壤资源竞争强度密切相关[21,23-25,30,33]。
3.2 间作种植方式对燕麦和豌豆根系构型的调控作用
植物常通过改变根系形态和空间分布调整植物种间对土壤养分和水分的竞争或协作利用关系,植物种间根系表现出趋避或聚集现象[23]。单播植物根系在水平和垂直方向生长往往无差异,而与其他植物同行混合或隔行间作时,根系形态常发生显著变化[21]。玉米(Zea mays)与小麦(Triticum aestivum)隔行间作后,小麦根系长度是单播小麦的1.5~2.0 倍;同时,表层土壤玉米的根系密度也是单播玉米的1.5 倍[38]。玉米与蚕豆(Vicia faba)隔行间作后,蚕豆根系大部分分布于表层土壤,而玉米根系却倾向于在深层土壤生长且部分根系已生长至蚕豆根系下方[38]。本研究发现隔行间作种植条件下,燕麦根系形态参数明显优于单播燕麦,说明燕麦与豌豆隔行间作能促进燕麦根系生长和发育,进而捕获土壤中更多的养分供给到茎叶,这也为隔行间作燕麦生物量高于单播燕麦提供证据。同时,朱亚琼等[17]对无芒雀麦和红豆草同行、异行混播比较研究发现,异行混播无芒雀麦的根体积、长度和表面积显著高于同行混合无芒雀麦,说明隔行间作种植模式对无芒雀麦根系发育存在促进作用,本研究结论与其一致。造成这种现象的原因主要为:1)隔行间作燕麦为获取豌豆根际生物固氮固定的氮素将促进垂直方向主根和水平方向侧根发育,进而获取更多的豆科根际氮素[38];2)燕麦和豌豆同行混合间作播种后,土壤养分出现生态位重叠,加剧二者地下资源竞争强度,进而限制燕麦根系发育和生长[17,21]。然而,二者隔行间作种植时,燕麦和豌豆土壤养分的生态位相对独立,使种间竞争强度变低。生长后期,喜氮型燕麦为获取豌豆根际更多氮素将通过改变根系形态策略获取更多氮素加速生长[23,39]。
与间作种植燕麦根系形态特征高于单播燕麦结论相反,同行混合和隔行间作豌豆根系形态参数显著低于单播豌豆。造成这种现象的原因可能为:1)间作体系中豌豆对光、土壤养分和水分竞争力较弱,植株为维持地上茎叶正常生长将大量光合产物优先输送至茎叶,进而影响地下根系生长,导致豌豆侧根发育较慢[17,20]。2)豌豆根系生长抑制作用随生长后期高位植物燕麦对光能资源拦截能力增强而增强[14,40],最终导致间作种植豌豆地上和地下生物量显著低于单播豌豆。综上所述,间作种植模式对间作组成中燕麦和豌豆根系形态的调控作用存在明显差异,间作能有效促进燕麦主根和侧根分别在土壤垂直和水平方向生长,而抑制豌豆主根和侧根发育。
3.3 氮添加与间作种植互作对燕麦和豌豆根系构型的可塑性
外源氮素添加和间作种植对禾本科和豆科植物根系形态及空间分布存在明显交互作用[14,28,40]。其中,氮素添加通过人为干扰土壤有效氮素的空间分布进而调控间作禾本科和豆科植物根系发育和生长[38]。然而,禾本科和豆科植物根系形态对外源氮添加的响应也表现出差异[28-29]。本研究发现氮添加通过增加燕麦根体积、根表面积和外部连接点等参数增加根系与土壤接触面,同时,氮添加可降低燕麦根系平均直径并促进毛根生长,上述两种途径可增强燕麦根系拓扑结构,并最终导致燕麦地下生物量增加。间作种植也可通过增加燕麦根系与土壤接触面积和增加分叉数进而增加根系分形维数和拓扑指数,最终增加燕麦地下生物量。由此可见,外源添加氮素和间作种植方式主要通过增加根系与土壤接触面积和促进分支以及侧根发育途径改变根系拓扑结构,进而增加燕麦根系生物量[21]。然而,间作方式和外源氮素添加通过降低豌豆根系连接数、根尖数和内部连接数进而降低根系拓扑指数,造成豌豆侧根生长和发育迟缓,获取土壤养分能力降低,并最终导致豌豆根系生物量降低[23]。可见,燕麦与豌豆间作种植时,竞争力强的燕麦通过改变根系构型在土壤资源获取方面占据主导地位,氮添加能促进间作燕麦根系生长,间作种植和氮添加互作对燕麦根系存在增益效应[14,40]。相比之下,间作种植豌豆的根系生长受到抑制,而这种抑制作用随外源氮添加而加剧[39]。综上所述,间作种植和外源氮添加对间作组成植物(燕麦和豌豆)根系具有较强的可塑性,其中,间作体系主导植物燕麦根系生长表现出明显促进作用,而处于劣势的豌豆根系发育表现出抑制作用[14,17,21,38,40]。
4 结论
氮添加和间作种植方式对燕麦和豌豆地上和地下生物量存在显著影响,而这一影响可能开始于地下根系养分竞争。间作种植和氮添加对燕麦根系生长存在增益效应,主要通过增加燕麦根系的土壤接触面积和侧根生长发育进而摄取较多的土壤养分,导致燕麦在间作体系中处于竞争优势,生物量显著增加。与之相反,间作种植和氮添加对豌豆侧根发育存在明显抑制作用,造成间作豌豆生物量较单播豌豆低。同时,间作种植和氮添加对燕麦和豌豆根系拓扑结构影响也存在明显差异,表现出对燕麦根系拓扑结构的促进作用而对豌豆则表现出抑制效应。另外,隔行间作高位燕麦和低位豌豆生长空间差异也使光能资源高效利用,并最终导致系统生产力较高。
