滞育时间对豆天蛾幼虫营养成分及代谢酶活性的影响
2024-03-20李宗男李情怡廖怀建
钱 蕾, 秦 毅, 张 佳, 李宗男, 邓 盼, 周 刚, 李情怡, 廖怀建,
(1.江苏省农业科学院休闲农业研究所,江苏 南京 210014; 2.扬州大学植物保护学院,江苏 扬州 225009; 3.淮安市农业科学院,徐淮地区淮阴农业科学研究所,江苏 淮安 223001; 4.江苏科技大学生物技术学院,江苏 镇江 212100)
豆天蛾(Clanisbilineatatsingtauica)属鳞翅目(Lepidoptera)天蛾科(Sphingidae)[1],是一种具有极高营养价值、药用价值和经济价值的食用昆虫[2-4]。豆天蛾幼虫富含蛋白质、氨基酸、不饱和脂肪酸、维生素等营养物质,食用豆天蛾幼虫有助于促进人体大脑发育、减缓细胞衰退、保持机体内分泌平衡等[5-7]。目前,人工养殖豆天蛾幼虫已经成为中国农村的特色农业产业,特别是在江苏省连云港市灌云县、淮安市涟水县等地,豆天蛾幼虫年产量达30 000 t,年产值近4.5×109元[8]。豆天蛾江苏种群1年发生1代,每年9月豆天蛾以老熟幼虫入土越冬,进入滞育状态,直至第二年6月中旬化蛹、羽化[8-9]。滞育是昆虫响应不良环境的一种生存适应策略。进入滞育状态的昆虫一般通过调控体内营养物质(主要包括脂类、氨基酸和碳水化合物等)的含量和种类应对低温、食物匮乏等逆境[10-11]。有研究结果表明,昆虫体内营养物质含量随滞育时间的延长而变化,如滞育期开心果壳螟(Arimaniacomaroffi)体内脂肪含量以及烟蚜茧蜂(Aphidiusgifuensis)体内甘油和海藻糖的含量均随着滞育时间的延长呈倒U型变化,而烟蚜茧蜂的糖原含量在滞育期呈线性下降[12-13]。除了贮存营养物质外,昆虫还通过降低代谢保障滞育期间的能量需求和滞育解除后生长发育所需的能量[14-16],如苹果实蝇(Rhagoletispomonella)在滞育期间会抑制呼吸速率和代谢速率,减少营养物质消耗[17],而滞育期豆天蛾幼虫的营养代谢机制尚不清楚。
目前,已有关于豆天蛾滞育的研究主要集中在温度、光周期条件对滞育幼虫生物学特性的影响或者滞育幼虫体内瞬时的营养成分变化,关于整个滞育周期内豆天蛾幼虫内源营养物质及代谢酶动态变化的研究较少。因此,本研究拟以滞育期豆天蛾幼虫为研究对象,对滞育过程中老熟幼虫体内营养物质的含量进行检测,分析肠道代谢酶活性的变化趋势,以期为阐明豆天蛾滞育期间对营养物质的调控机理奠定理论基础,为豆天蛾幼虫绿色、高效繁育提供科技指导。
1 材料与方法
1.1 供试昆虫
2022年9月于江苏省农垦农业发展股份有限公司云台分公司豆丹养殖基地(119.29°E,34.59°N)采集了同一批大小一致的豆天蛾老熟幼虫,带回室内养虫室进行观察,作为试验虫源。条件设置为:温度(15±1) ℃,相对湿度70%±5%,光周期14 h(光照时间)∶10 h(黑暗时间)。
1.2 试验设计
待豆天蛾老熟幼虫入土做虫室,并且保持静止状态时,即认定其进入滞育期。采集即将有入土行为的老熟幼虫作为对照(滞育前);分别取滞育时间为30 d、60 d、90 d、120 d、150 d的豆天蛾幼虫为处理组。分别采集对照和处理组的豆天蛾幼虫,用去离子水洗净后放入50.0 ml离心管中,放入液氮中速冻,冷冻保存于-80 ℃超低温冰箱(DW-86L388J,购自青岛海尔生物医疗股份有限公司),用于豆天蛾幼虫体内蛋白质、氨基酸、脂肪酸含量的测定,3头幼虫为1个样本,每组重复5次。解剖豆天蛾幼虫取其中肠组织,放入1.5 ml离心管中,液氮速冻,冷冻保存于-80 ℃超低温冰箱,用于测定幼虫肠道代谢酶(包括海藻糖酶、碱性磷酸酶、己糖激酶、丙酮酸激酶和山梨醇脱氢酶)活性,10头幼虫的肠组织为1个样本,每组重复5次。
1.3 测定项目与方法
1.3.1 豆天蛾幼虫体内营养物质含量的测定 采用凯氏定氮法测定豆天蛾幼虫体内蛋白质的含量。称取0.2 g样品于消化管中,加入0.4 g硫酸铜、6.0 g硫酸钾及20 ml硫酸于消化炉中进行消化,当消化炉温度达到420 ℃之后,继续消化1 h,消化管中的液体呈蓝绿色并澄清透明后,取出冷却,然后加入20 ml水,放冷,于自动凯氏定氮仪(K9840,购自济南海能仪器股份有限公司)中蒸馏7 min,于接收瓶中加指示剂混合溶液1~2滴和10 ml硼酸溶液(20 g/L),接收蒸馏液到200 ml,然后用盐酸标准溶液(0.100 mol/L)滴定,终点为浅灰红色,同时做试剂空白。根据公式计算豆天蛾幼虫体内蛋白质的含量,换算系数为6.25。
通过氨基酸自动分析仪(LA8080;购自日立公司)测定豆天蛾幼虫体内17种氨基酸[天冬氨酸(Aspartic acid,Asp)、苏氨酸(Threonine,Thr)、丝氨酸(Serine,Ser)、谷氨酸(Glutamic acid,Glu)、甘氨酸(Glycine,Gly)、丙氨酸(Alanine,Ala)、半胱氨酸(Cysteine,Cys)、缬氨酸(Valine,Val)、蛋氨酸(Methionine,Met)、异亮氨酸(Isoleucine,Ile)、亮氨酸(Leucine,Leu)、酪氨酸(Tyrosine,Tyr)、苯丙氨酸(Phenylalanine,Phe)、赖氨酸(Lysine,Lys)、组氨酸(Histidine,His)、精氨酸(Arginine,Arg)、脯氨酸(Proline,Pro)]的含量。采用高效液相色谱法(HPLC)测定幼虫体内色氨酸(Tryptophan,Trp)的含量。色谱柱是Agilent C18(4.6 mm×150.0 mm×5.0 μm),柱温设为35 ℃,进样量为10.0 μl,流速为1.2 ml/min,波长为280 nm。
采用气相色谱-质谱联用仪测定豆天蛾幼虫体内脂肪酸的含量。色谱柱为HP-88 Agilent(100.00 m×0.25 mm×0.20 μm),进样温度设为290.0 ℃,载气He,流速1.0 ml/min,不分流。质谱条件设为:离子源温度280.0 ℃,传输线温度280.0 ℃,溶剂延迟5 min,扫描范围30~400 U,离子源为EI源,电子能量70 eV。升温程序:初始温度100.0 ℃保持13 min,以10.0 ℃/min的速率升温至180.0 ℃,保持6 min,以1.5 ℃/min的速率升温至192.0 ℃,保持6 min,再以3.5 ℃/min的速率升温至240.0 ℃保持4 min。
1.3.2 豆天蛾幼虫肠道代谢酶活性的测定 按照青岛科创质量检测有限公司试剂盒的说明书,测定不同滞育时间豆天蛾幼虫肠道代谢酶(包括海藻糖酶、碱性磷酸酶、己糖激酶、丙酮酸激酶和山梨醇脱氢酶)的活性变化。
1.4 数据计算与分析
按公式(1)计算蛋白质含量:
蛋白质含量(%)=[(试样消耗盐酸量-空白消耗盐酸量)×0.100×0.010]/(样品量×消化液体积/100)×6.250×100×100%
试验数据均采用Microsoft Excel 2016进行整理,利用IBM SPSS Statistics 25.0软件进行统计分析,用GraphPad Prism8.3.0软件作图。应用单因素方差分析明确不同滞育时间对豆天蛾幼虫体内蛋白质、氨基酸、脂肪酸含量以及肠道代谢酶活性的影响。采用Tukey检验法分析不同滞育时间处理之间的差异显著性(P<0.05)。
2 结果与分析
2.1 滞育时间对豆天蛾幼虫体内蛋白质含量的影响
图1显示,与对照相比,滞育期豆天蛾幼虫体内蛋白质含量均显著提高(P<0.05);随着滞育时间的延长,滞育150 d(滞育后期)的豆天蛾幼虫体内蛋白质含量高达21.03%,与滞育30 d、60 d、90 d、120 d的豆天蛾幼虫体内蛋白质含量相比,分别提高了4.16个百分点、3.40个百分点、2.60个百分点、3.96个百分点。

图中不同小写字母表示不同滞育时间的豆天蛾幼虫体内蛋白质含量差异显著(P<0.05)。
2.2 滞育时间对豆天蛾幼虫体内氨基酸含量的影响
本研究测定了滞育期豆天蛾幼虫体内18种氨基酸含量的变化,结果(表1)表明,滞育后期(150 d)豆天蛾幼虫体内18种氨基酸含量均显著高于对照(P<0.05)。滞育期豆天蛾幼虫体内苏氨酸(Thr)、异亮氨酸(Ile)、亮氨酸(Leu)、苯丙氨酸(Phe)和赖氨酸(Lys)这5种必需氨基酸的含量均显著高于对照(P<0.05);滞育90 d、120 d、150 d豆天蛾幼虫体内色氨酸(Trp)含量分别比对照高0.06个百分点、0.06个百分点、0.07个百分点(P<0.05)。豆天蛾第一限制氨基酸[苏氨酸(Thr)]含量在滞育后期(150 d)达到最高值,为0.60%,比滞育30 d、60 d、90 d、120 d分别高0.12个百分点、0.14个百分点、0.04个百分点、0.09个百分点。

表1 滞育时间对豆天蛾幼虫体内氨基酸含量的影响
2.3 滞育时间对豆天蛾幼虫体内脂肪酸含量的影响
滞育期的豆天蛾幼虫体内共检测到6种脂肪酸,分别是棕榈酸、棕榈油酸、硬脂酸、油酸、亚油酸和亚麻酸,其中亚油酸和亚麻酸是人体必需脂肪酸。表2显示,与对照相比,滞育60 d豆天蛾幼虫体内亚油酸含量显著降低0.030个百分点(P<0.05),滞育30 d、90 d、120 d、150 d亚油酸含量变化不显著。滞育30 d、90 d、120 d、150 d豆天蛾幼虫体内亚麻酸含量分别高于对照0.140个百分点、0.060个百分点、0.060个百分点、0.080个百分点(P<0.05)。
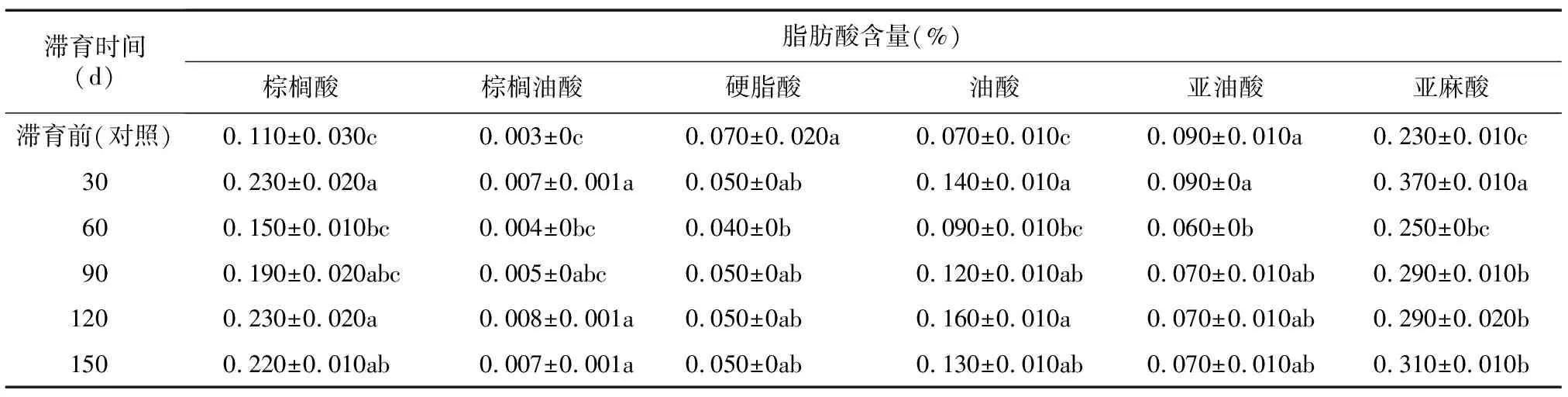
表2 滞育时间对豆天蛾幼虫体内脂肪酸含量的影响
2.4 滞育时间对豆天蛾幼虫肠道代谢酶活性的影响
2.4.1 海藻糖酶活性 图2显示,随着滞育时间的延长,豆天蛾幼虫肠道海藻糖酶活性表现出先缓慢升高后降低再升高的趋势。滞育90 d豆天蛾幼虫肠道海藻糖酶活性高达35.46 U/g,显著高于对照(P<0.05);滞育120 d豆天蛾幼虫肠道海藻糖酶活性最低,为28.74 U/g,显著低于对照(P<0.05)。与对照相比,滞育60 d、150 d豆天蛾幼虫肠道海藻糖酶活性显著升高,分别提高10.29%、9.39%(P<0.05)。
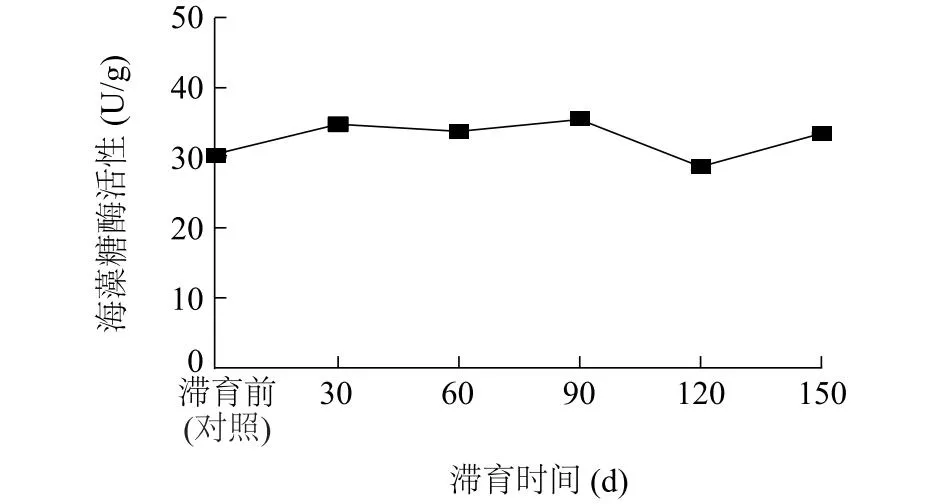
图2 滞育时间对豆天蛾幼虫肠道海藻糖酶活性的影响
2.4.2 碱性磷酸酶活性 图3显示,滞育120 d豆天蛾幼虫肠道碱性磷酸酶活性高达40.54 U/g,与对照相比,显著提高7.47%(P<0.05),且分别比滞育30 d、60 d、90 d、150 d豆天蛾幼虫肠道碱性磷酸酶活性显著提高14.26%、8.34%、14.22%、11.14%(P<0.05)。与对照相比,滞育30 d、90 d豆天蛾幼虫肠道碱性磷酸酶活性显著降低5.95%、5.91%(P<0.05)。
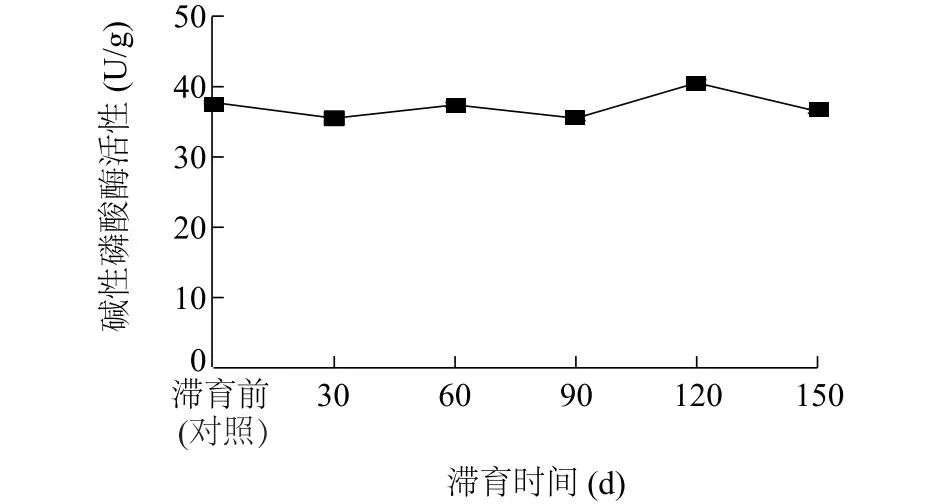
图3 滞育时间对豆天蛾幼虫肠道碱性磷酸酶活性的影响
2.4.3 己糖激酶活性 图4显示,随着滞育时间的延长,豆天蛾幼虫肠道己糖激酶活性表现出先降低后升高再降低的趋势。滞育30 d豆天蛾幼虫肠道己糖激酶活性最低,为15.07 U/g,比对照低4.76%(P<0.05),且分别比滞育60 d、90 d、120 d、150 d低8.33%、12.52%、18.80%、13.54%(P<0.05)。滞育120 d豆天蛾幼虫肠道己糖激酶活性高达18.56 U/g,比对照高17.30%(P<0.05)。
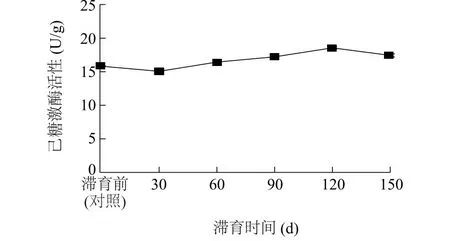
图4 滞育时间对豆天蛾幼虫肠道己糖激酶活性的影响
2.4.4 丙酮酸激酶活性 图5显示,随着滞育时间的延长,滞育90 d豆天蛾幼虫肠道丙酮酸激酶活性达到最高,为8.90 U/g,与对照相比,显著提高11.16%(P<0.05),且比滞育30 d、60 d、120 d显著提高8.01%、23.44%、12.47%(P<0.05)。滞育60 d豆天蛾幼虫肠道丙酮酸激酶活性最低,为7.21 U/g,与对照相比,显著降低9.95%(P<0.05)。

图5 滞育时间对豆天蛾幼虫肠道丙酮酸激酶活性的影响
2.4.5 山梨醇脱氢酶活性 图6显示,随着滞育时间的延长,滞育120 d豆天蛾幼虫肠道山梨醇脱氢酶活性达到最高,为4.21 U/g,与对照相比,显著提高18.37%,且比滞育30 d、60 d、90 d、150 d提高22.50%、12.27%、23.58%、22.86%(P<0.05)。滞育60 d山梨醇脱氢酶活性为3.75 U/g,与对照、滞育30 d、90 d、150 d相比,显著提高5.34%、9.01%、9.97%、9.33%(P<0.05)。

图6 滞育时间对豆天蛾幼虫肠道山梨醇脱氢酶活性的影响
3 讨 论
滞育期昆虫体内的营养物质含量随着滞育时间的延长而波动,以应对不同滞育阶段的能量需求,维持滞育期间的代谢[18-19]。滞育期间昆虫血淋巴中的氨基酸既可以作为代谢的中间产物,用于合成代谢与分解代谢,又可调节昆虫血淋巴渗透压,提高昆虫抗寒性[20-22]。本研究发现,与对照相比,滞育期的豆天蛾幼虫体内蛋白质含量显著升高,滞育后期(150 d)18种氨基酸的含量均显著高于对照。推测这是因为昆虫通过积累滞育关联蛋白质来储存氨基酸,这些蛋白质在滞育前开始积累,并在滞育期间持续合成[10,18]。西南玉米螟(Diatraeagrandiosella)、苹果蠹蛾(Cydiapomonella)等昆虫滞育期间也表现为增加体内游离氨基酸含量和积累特定的蛋白质[10]。然而前人也有不同的发现,中红侧沟茧蜂(Microplitismediator)滞育过程中蛋白质的含量变化平缓,无显著差异[23],可能是因为滞育期能量需求与合成代谢的不同造成昆虫体内生理响应程度的差异[17]。笔者前期研究短期滞育的豆天蛾时也发现,滞育28 d豆天蛾幼虫体内粗蛋白含量变化不显著[24],这可能是因为滞育前期营养物质还在积累中,蛋白质变化还未达到显著水平。
脂肪酸不仅是滞育昆虫储存的营养物质,还是其低温下的抗冻保护剂,可以保障生物膜的正常功能[25-28]。郭明明等[29]研究滞育期豆天蛾幼虫营养物质瞬时变化规律时发现,滞育期豆天蛾幼虫必需脂肪酸亚麻酸的含量明显低于未滞育幼虫与滞育解除后的幼虫。而本研究分析整个滞育期豆天蛾幼虫脂肪酸含量动态变化时发现,幼虫体内亚麻酸的含量在滞育初期急剧增加,并在滞育30 d达到最高。推测可能是豆天蛾幼虫响应滞育初期的营养代谢反应更强烈,当滞育状态逐渐稳定时,幼虫体内代谢活动也越稳定。笔者前期研究也发现,滞育28 d豆天蛾幼虫体内亚麻酸含量明显升高,与本研究的结果一致[24]。
酶活性的稳定对生物代谢有至关重要的作用,酶活性越稳定,生物代谢就越稳定,生物适应能力就越强[30-32]。本研究分析了滞育时间对豆天蛾幼虫肠道代谢酶活性的影响,发现滞育中期(90 d)豆天蛾幼虫肠道海藻糖酶活性达到最高,但在滞育120 d、150 d波动较大。有研究发现,桃小食心虫、梨小食心虫、家蚕体内海藻糖酶活性随滞育时间的延长而逐渐降低,从而使海藻糖含量维持在较高水平,提高虫体抗逆能力[15,20]。豆天蛾幼虫肠道碱性磷酸酶活性在滞育120 d达到最高,并在滞育后期迅速降低,可能是因为碱性磷酸酶对昆虫滞育时间有一定的调控作用[33]。己糖激酶是糖酵解途径的关键酶。滞育前期(30 d)豆天蛾幼虫肠道己糖激酶的活性降低,说明此时豆天蛾幼虫肠道糖酵解较少;滞育30~120 d己糖激酶活性逐渐升高,说明糖原酵解正在加强,豆天蛾幼虫在滞育期间不断积累糖原,为其生命活动提供能量,这一结果与张顺益等[33]、王鹏等[34]的研究结果一致。豆天蛾幼虫肠道山梨醇脱氢酶活性在滞育120 d达到最高,并在滞育150 d迅速降低,说明豆天蛾幼虫在滞育后期代谢活动逐渐减弱,降低体内营养物质消耗,从而顺利度过严寒季节[35]。
本研究结果表明,豆天蛾幼虫通过调控体内关键代谢酶的活性,使其在滞育期间的营养物质和能量代谢得到调节,提高虫体抵御外界不良环境条件的能力,从而顺利越冬。另外,肠道微生物能够通过调节宿主关键酶的活性进而影响昆虫营养物质的合成[36]。未来可结合微生物学与代谢组学对豆天蛾幼虫滞育期间关键酶、基因的作用进行联合分析,研究肠道细菌通过代谢酶对宿主营养合成与代谢的调控机制。本研究结果可以为下一步研究温度、光周期调控豆天蛾幼虫滞育机制奠定理论基础,为豆天蛾产业打破幼虫冬季滞育提供技术支撑,从而提高豆天蛾的年产量;还可以为豆天蛾营养物质的开发与利用提供技术指导,有利于提高产品的附加值,最终为豆天蛾产业结构的优化升级提供科学依据。
