燕麦全基因组SSR 位点鉴定及多态性标记开发
2024-03-12陈凯凌武涛徐逸群高佳张美俊李欣贾举庆
陈凯凌 武涛,2 徐逸群,2 高佳,2 张美俊,2 李欣 贾举庆,2
(1.山西农业大学农学院,太谷 030801;2.山西农业大学杂粮种质资源创新与分子育种国家实验室(筹),太原 030031)
燕麦(Avena sativa L.)为异源六倍体(AACCDD,2n=6X=42),包含A、C、D 三个亚基因组,每组包含7 条染色体,是一种重要的特色杂粮。中国作为裸燕麦(莜麦)的起源地,在内蒙古、山西等地广泛种植[1]。燕麦是一种可全谷物食用的健康谷物,由于其含有独特的燕麦蛋白、燕麦多肽、β-葡聚糖等营养物质,对于人体健康与疾病防治有重要作用而受到广泛关注[2]。
由于燕麦基因组较大、基因组结构复杂并缺少高质量的参考基因组,相较于同为禾本科的小麦、水稻、玉米而言,燕麦功能基因发掘与分子进化的相关研究起步较晚[3]。在2022 年,相继公布了2 个六倍体燕麦参考基因组[4-5],为燕麦基因组学研究提供了强有力的工具。
开发SSR 标记的方法主要有根据全基因组测序结果开发[6]、转录组测序结果开发[7]、SSR 富集文库构建开发[8]及利用已报道的SSR 标记交叉扩增[9]。基于转录组开发SSR 标记所包含的物种基因信息较少,构建SSR 富集文库开发成本较高,近缘物种交叉扩增有效率较低。基于参考基因组开发SSR 标记简单、经济并且包含基因信息丰富。
在燕麦中,基于基因组开发的SSR 分子标记数量有限,主要集中在燕麦抗冠锈病[9-10]、高矮[11]、β-葡聚糖[12]含量相关SSR 标记的开发与基因定位,其次是基于EST 序列开发的SSR 标记[13],关于燕麦皮裸性状的多态性SSR 标记鲜有报道。
本研究利用公布的六倍体裸燕麦“三分三”全基因组进行SSR 位点检索并对其进行特征分析,同时开发出52 对多态性SSR 引物,并进行克隆测序验证其真实性。在F2遗传群体材料中进行小规模扩增,为后续燕麦皮裸基因克隆、遗传图谱构建、群体多样性分析等提供了科学的参考依据。
1 材料与方法
1.1 材料
选取2 个六倍体裸燕麦白燕2 号(父本)与皮燕麦Banner(母本)作为杂交亲本。双亲种质资源由山西农业大学生物技术实验室提供。将白燕2 号和Banner 通过人工去雄杂交获得F1单株种子,F1种子自交、结实、收获、鉴定,获得F2种子。于2022 年3 月,将F2及双亲种子播种在山西农业大学农作站实验基地,期间进行科学的田间水肥管理。在F2群体中分别在2 个极端表型(皮燕麦和裸燕麦)中随机选取皮裸性状差异明显的植株各10 株,共计20 个单株为试验材料。
1.2 方法
1.2.1 燕麦全基因组序列下载及SSR 位点检索 在NCBI 网站下载裸燕麦“三分三”参考基因组序列,登录号为PRJNA727473。利用TBtools(v1.09876-63)[14]中SSR 检索工具对全基因组序列SSR 位点进行搜索。搜索标准为单碱基至少重复10 次、双碱基至少重复6 次,三碱基、四碱基、五碱基与六碱基至少重复5 次。同一SSR 最长不超过2 kb。将搜索数据利用Excel 与SPSS 软件进行统计分析。
1.2.2 燕麦SSR 引物设计和PCR 验证 选取SSR 位点上下游500 bp 序列,将和控制燕麦籽粒皮裸性N1连锁的分子标记序列[15]与“三分三”基因组比对,判断N1 在染色体上的物理区段,从该区段中挑选长度大于40 bp 的SSR 位点100 个,在基因组上截取SSR 位点上下游500 bp 序列,将这些序列分别比对“三分三”和“Sang”基因组,以去除非特异性位点及在两个测序的燕麦品种中无多态性的SSR 位点。剩余的序列利用Primer3 在线设计SSR 引物,引物设计参数设置为长度18-22 bp,退火温度为55-59℃,扩增产物长度为250-400 bp;应避免引物间二聚体、发夹结构等结构出现。并将设计引物在“三分三”和“Sang”基因组中进行电子PCR 分析,去除有非特异扩增与在2 个燕麦基因组中扩增片段相同的引物,提高其特异性与多态性。将设计完成的引物序列交由北京华大科技有限公司合成。
1.2.3 皮裸性状调查 对杂交双亲及F2群体中选取的燕麦植株进行皮裸性状调查。
1.2.4 DNA 提取 选取试验材料苗期的幼嫩叶片,按照北京天根生化科技有限公司提供的DNA 提取试剂盒提取DNA。用1%琼脂糖凝胶电泳检测其完整性,并将DNA 稀释为50 ng/μL,-20℃保存备用。
1.2.5 燕麦SSR 引物的开发与多态性检测 将亲本DNA 进行初步PCR 扩增,筛选出具有目的条带的SSR 引物,进一步在20 个F2群体燕麦单株和双亲DNA 中进行PCR 扩增,筛选出多态性明显、条带清楚的稳定SSR 引物。SSR-PCR 反应体系:1.0 μL DNA(50 ng/μL)、左右引物各0.5 μL(0.5 μmol/L)、1.0 μL 10×PCR buffer(含Mg2+)、0.8 μL dNTP(0.23 nmol/L)、0.12 μL Taq DNA 聚合酶(5 U)和ddH2O 6.08 μL。反应程序:94℃ 4 min;94℃ 45 s,56℃(依据引物最佳退火温度而定)45 s,72℃ 20 s,30 个循环;72℃ 10 min,4℃保存。PCR 扩增产物用8%非变性聚丙烯酰胺凝胶电泳检测,染色、显影,并拍照保存。
1.2.6 SSR 引物的验证 经非变性聚丙烯酰胺凝胶检测后,回收扩增的目的产物,加入100 μL 无菌水,用无菌玻璃棒将其研磨充分,55℃溶解30 min,离心;取1 μL 上清液作为模板DNA 进行PCR 扩增,扩增条件同1.2.5。将扩增产物经2%琼脂糖凝胶电泳分离,用DNA 回收试剂盒将目的基因进行切胶回收,并连接至T 载体,转入JM109 感受态细胞,挑选阳性克隆菌液送至北京华大科技有限公司进行测序。利用Jalview 软件对测序序列进行多序列比对。
2 结果
2.1 燕麦全基因组SSR位点数量分布特征
在NCBI 网站下载六倍体裸燕麦“三分三”及“Sang”全基因组序列,“三分三”的基因组总长为11 Gb。燕麦基因组共分为3 个亚基因组,每个亚基因组各有7 条染色体,共计21 条染色体,每条染色体长度为293.84-701.82 Mb。利用TBtools 软件在21条染色体上共检索到828 138 个SSR 位点(表1),在1A(46 585)、3C(49 132)和4C(46 713)位点最为丰富,在6D(22 043)分布最少。在全基因组中,平均每12.90 kb 会出现一个SSR 位点,在1C 出现SSR 位点距离最长;全基因组平均SSR 密度为78.16个/Mb,在2A 上SSR 分布密度最高。表明燕麦全基因组SSR 位点数量丰富、分布均匀。
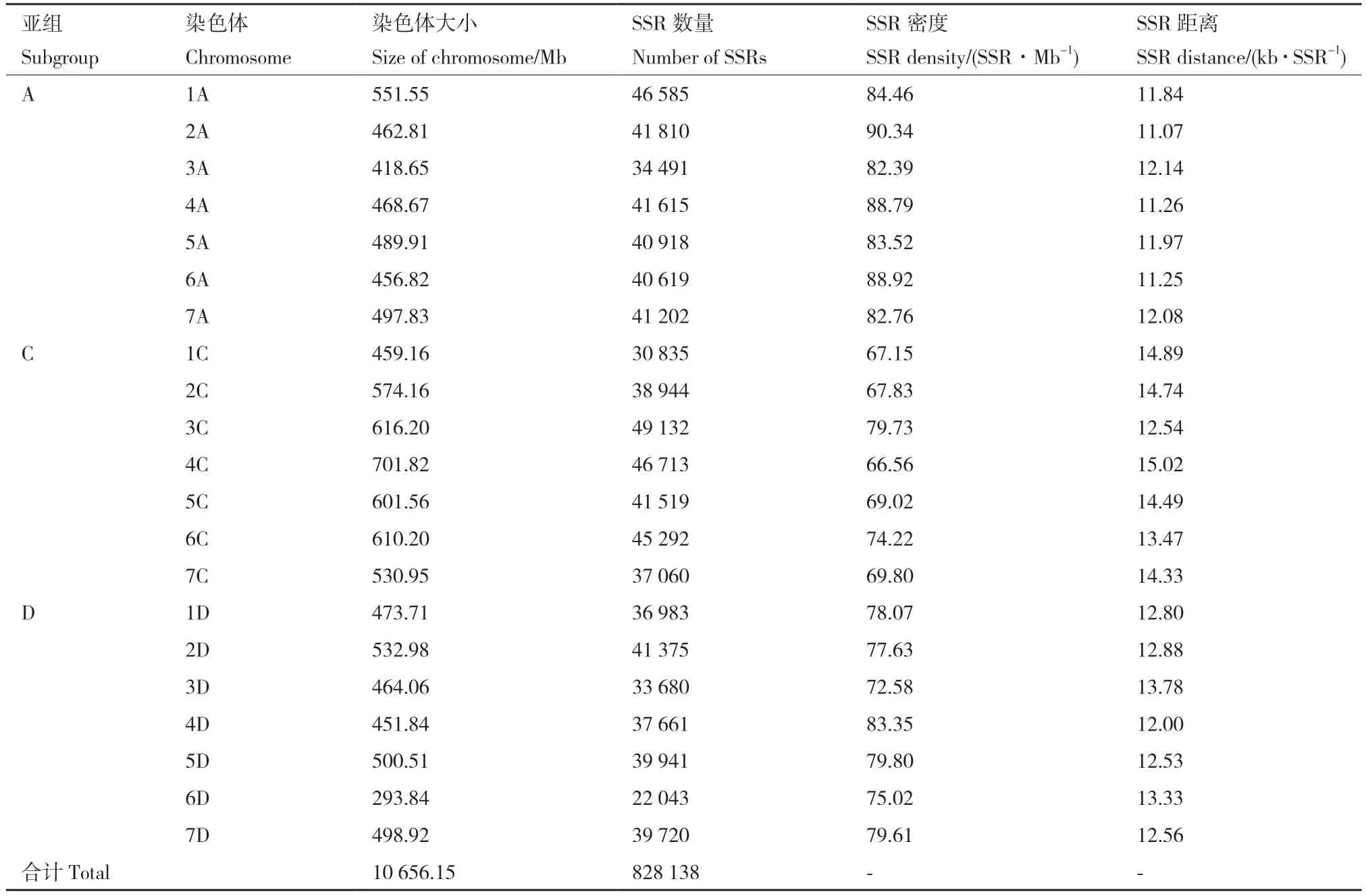
表1 SSR 数量分布特征Table 1 SSR quantitative distribution characteristics
燕麦染色体大小和SSR 密度、SSR 距离、SSR数量之间均符合正态分布,结果显示,染色体大小与SSR 数量之间呈正相关关系,相关系数为0.81。同时,染色体大小与SSR 密度、SSR 距离之间无相关关系(表2),在A、C、D 三个亚基因组间C 亚基因组染色体最长、SSR 数目最多,但是SSR 密度最低;A 亚基因组染色体长度与SSR 数目适中,但SSR 密度最高(表3)。

表2 相关性分析Table 2 Correlation analysis

表3 亚基因组间SSR 分布特征Table 3 SSR distribution characteristics among subgroups
2.2 燕麦全基因组SSR位点基元重复次数与类型分布特征
在燕麦全基因组中共检测到6 种核苷酸重复类型(表4),其中,单核苷酸(29.85%)、双核苷酸(33.69%)和三核苷酸(33.79%)重复类型占主导优势;四核苷酸(1.81%)、五核苷酸(0.28%)、六核苷酸(0.59%)占比较少。在全部SSR 位点中,重复次数在5-567 次之间均有检索到,最小重复次数为5 次,最多重复次数为一个双核苷酸重复类型的SSR 重复567 次。在重复次数的数量上分析,大于10 次的重复次数最多(310 749),在不大于10 次的重复次数中5 次(177 356)和6 次(185 716)重复次数数量最多,其余重复次数数目均较少。在核苷酸重复类型和重复次数两个复合条件下分析,随着核苷酸重复类型的基元序列变长和核苷酸重复次数增加,SSR 位点数量逐渐减少,其中,三核苷酸重复、重复5 次数目最多(164 814),五核苷酸重复、重复大于10 次数目最少(96)。
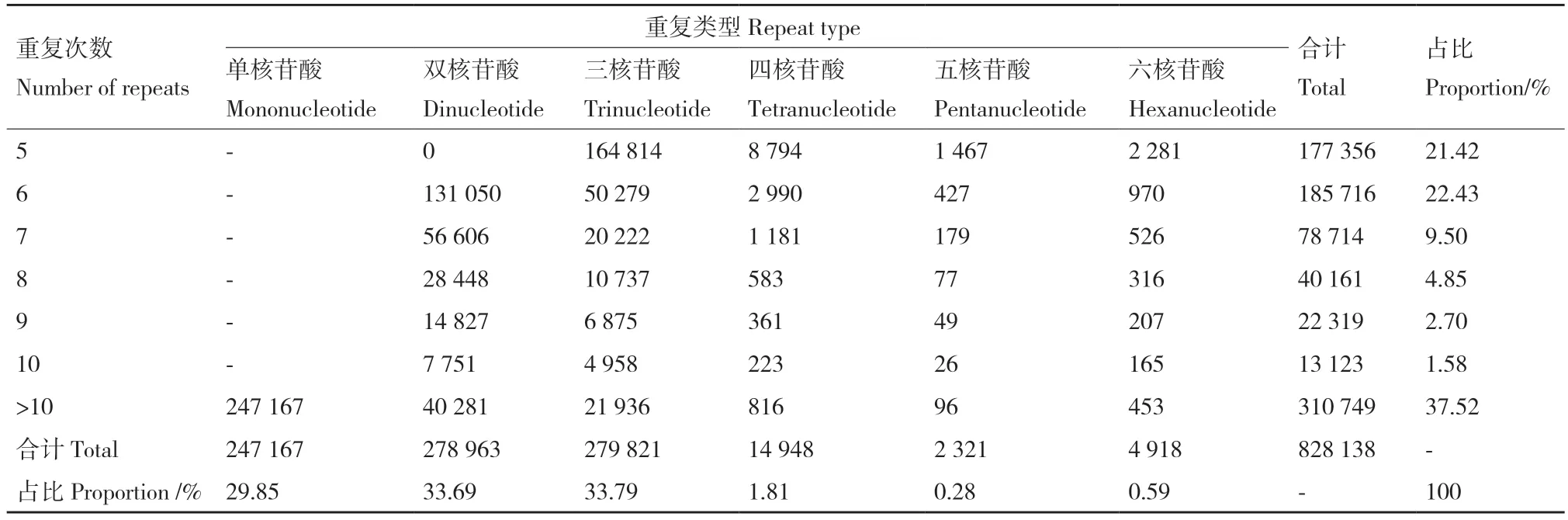
表4 燕麦全基因组SSR 重复类型、重复次数和占比Table 4 SSR repeat type,repeat number and proportion of whole genome of oat
2.3 燕麦全基因组SSR位点基元类型分布特征
在燕麦全基因组中共检索到1 548 种不同的重复类型(表5),不同基元重复类型在6 种不同核苷酸类型中均有出现。在单核苷酸重复中,2 种基元类型A/T(58.80%)与C/G(41.20%)占比大致相当;在双核苷酸重复中4 种基元类型占比差异较大,AG/CT(49.15%)为优势基元,CG/CG(1.67%)占比最小;在三核苷酸重复中AAC/GTT(24.49%)、ATC/ATG(20.50%)为优势基元;在四、五、六核苷酸重复中的优势基元分别为AAAT/ATTT(15.29%)、TAAGA/TCTTA(39.21%)、CCTGGG/CCCAGG(41.91%)。随着核苷酸重复类型基元序列变长,基元类型显著增加,但各基元数量明显减少。在检索的所有优势基元中,含有A、T 碱基的SSR数量明显高于含有C、G 碱基的SSR 数量,说明在燕麦全基因组SSR 位点中含有大量A、T 碱基。

表5 燕麦全基因组各SSR 重复类型中优势基元类型及分布Table 5 Types and distribution of dominant motifs in each SSR repeat type of oat genome
2.4 燕麦全基因组SSR位点长度分布特征
决定一个SSR 位点能否使用取决于其多态性的有无,序列长度越长的SSR 具有多态性的可能越大。对燕麦全基因组SSR 位点长度分布进行分析,能够检索到最长的SSR 序列为1 587 bp,最短的为11 bp,SSR 平均长度为17.55 bp。其中,SSR 序列长度为11(123 990)、12(177 110)和15 bp(176 477)的SSR 位点数目占主导优势(图1)。长度在12 bp以下(低度多态性)的SSR 位点数目为123 990 个,长度在12-20 bp(中度多态性)的SSR 位点数目为582 176 个,长度大于20 bp(高度多态性)的SSR位点数目为121 972 个,表明在燕麦全基因组中SSR位点呈现中度多态性。
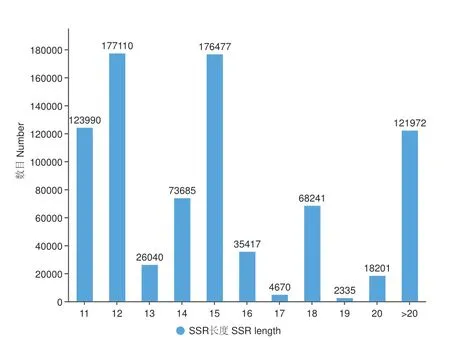
图1 燕麦基因组SSR 长度分布Fig.1 SSR length distribution of oat genome
2.5 燕麦4D染色体末端SSR标记开发、验证及多态性检测
将设计的100 对引物在2 个亲本DNA 中进行PCR 扩增,使用非变性聚丙烯酰胺凝胶电泳检测,除10 对引物无扩增片段,其余引物扩增产物均符合预期长度,有效扩增率为90%;在有效扩增的引物中52 对在2 个亲本间具有多态性(表6),扩增多态率为58%。将多态性SSR 引物进一步在F2群体的20 个单株中进行扩增,均可以看到多态性条带(图2),52 对引物在亲本中的扩增片段进行克隆测序,并将结果进行比对“三分三”基因组发现,PCR 扩增位点与SSR 引物设计位点一致,且PCR 扩增片段的多态性确由SSR 造成(图3)。说明本研究基于燕麦基因组开发的SSR 分子标记,无论是PCR 扩增的特异性还是产物的多态性,都可用于后续的燕麦N1及其他基因的定位研究。

图2 引物A-1(A)和A-2(B)在试验材料中扩增结果Fig.2 Amplification results of primers A-1(A)and A-2(B)in experimental materials

图3 引物A-1(A)和A-2(B)在2 个亲本中的扩增序列比对结果Fig.3 Amplification sequence alignment of primers A-1(A)and A-2(B)in two parents

表6 开发的燕麦多态性SSR 引物信息Table 6 Information of SSR primers developed for oats polymorphism
3 讨论
由于第三代测序技术的出现,使得小麦[16]、燕麦[4-5]等六倍体作物的参考基因组组装得以实现。本研究利用最新公布的燕麦参考基因组,对全基因组进行SSR 位点鉴定与特征分析,并利用鉴定的SSR 位点开发52 对多态性引物并克隆测序验证其真实性,证明基于参考基因组开发SSR 标记有效可行,相较于转录组测序、物种间交叉扩增和构建富集文库的开发方法[6-9]简单高效,为后续大规模开发燕麦SSR 标记提供科学参考依据。
通过对燕麦全基因组SSR 位点鉴定发现,在数量上远高于一般农作物如玉米[17]、苦荞[18]、谷子[19],以及蔬果类如花生[20]、黄瓜[21]、苹果[22]、甜菜[23]等基因组。此现象的主要原因是由于燕麦为六倍体作物,染色体条数多,基因组庞大。但与同为六倍体的小麦中国春基因组[24]相比,两者SSR 数量相差较大,主要在于检索SSR 标准不同,小麦中未检索单核苷酸的重复类型。已有研究表明,随着基因组的扩大,SSR 位点数量不会无限扩增,其分布密度会相对减少[25],在燕麦中也符合这一规律。燕麦基因组相比于四倍体马铃薯[26]的SSR 密度减少较多;同时发现在燕麦基因组的不同亚组间也符合这一规律,表明在不同物种间和同一物种的不同亚组间,DNA 在同一条染色体复制过程中发生的滑动错配的概率会随着染色体增长而降低。
在燕麦SSR 的优势基元中,与薏苡[27]、黄麻[28]、烟草[29]、绿豆[30]等其他物种相比,不同物种SSR优势基元不尽相同,但比较发现不同物种优势基元中均含有丰富的A、T 碱基。这种现象可能是由于A、T 碱基之间含有2 个氢键相较于C、G 碱基之间3 个氢键更容易被打开所导致。
在燕麦基因组中检索得到的所有SSR 序列长度占全基因组序列比例远小于动物的SSR 含量,相较于植物如拟南芥和玉米,其SSR 含量也较少[31]。表明在燕麦基因组中SSR 位点数量虽然丰富,但是其含量并不丰富。已有研究发现,SSR 位点在基因组中的扩张与物种进化密切相关[31],燕麦较低的SSR含量表明燕麦相较于鱼类和拟南芥等物种进化相对缓慢。燕麦SSR 含量与同为麦类作物的节节麦[32](0.10%)相比含量稍高,节节麦是六倍体小麦(AABBDD)D 套染色体的二倍体祖先。燕麦由于染色体结构复杂,导致染色体间重组更为频繁,从而导致更易出现SSR 位点,同为六倍体的小麦染色体间的重组导致物种基因组出现马赛克结构[16],推测燕麦SSR 含量可能与燕麦基因组的马赛克结构相关。
本研究开发的52 对多态性SSR 标记,能够用于后续燕麦相关基因定位克隆、DNA 指纹图谱和遗传图谱构建、种群群体分析等研究。但是开发SSR标记随机性仍旧较大,比较依赖参考基因组的准确性,通过非变形聚丙烯酰胺凝胶判别条带人为误差较大,而且耗时耗力,后续可以开发KASP 标记进行基因分型或者使用毛细管电泳的方法检测 SSR 序列的多态性。
4 结论
通过对燕麦“三分三”参考基因组进行SSR 位点检索发现,该基因组SSR 位点多样、数量丰富。根据检索结果设计引物,在构建的F2代群体小规模进行扩增,开发了52 对具有多态性的引物,并克隆测序验证其真实性。表明在燕麦“三分三”参考基因组中开发SSR 标记可行有效,开发的多态性SSR标记可以用于后续燕麦皮裸基因定位。
