三峡水库入库溪流中麦氏花翅蜉(蜉蝣目:四节蜉科)的生活史研究
2024-03-04敖偲成李浩然李先福蔡庆华
杨 坤,田 震,石 欣,敖偲成,4,李浩然,李先福,6,7,谭 路,蔡庆华,4
1.大理大学东喜玛拉雅研究院,云南 大理 671003
2.中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,湖北 武汉 430072
3.吉首大学,湖南 吉首 416000
4.中国科学院大学,北京 100049
5.九江学院资源环境学院,江西 九江 332005
6.中国三江并流区域生物多样性协同创新中心,云南 大理 671003
7.云南省高校洱海流域保护与可持续发展研究重点实验室,云南 大理 671003
生活史信息是水生昆虫生态学研究的基础[1],在解释种群自然规律、确定物质和能量动态、完整呈现实验结果等方面都具有非常重要的意义[2],能反映出河流的环境健康状况[3]和生态系统结构完整性[4]。同时,掌握水生昆虫生活史性状对不同温度生境的响应模式,对预测未来气候变化的潜在影响具有非常重要的意义[5]。因此,在当前全球淡水生物多样性急剧下降的背景下[6],研究水生昆虫的分布、生长发育过程对掌握水生生物现状和评估生态环境状况均具有重要作用,可为构建和完善水生态环境保护体系提供科学参考。

花翅蜉属(Baetiella)是UÉNO于1931年建立的蜉蝣目(Ephemeroptera)四节科(Baetidae)新属,主要分布于古北区和东洋区[17-19]。目前已发现并报道了16种花翅蜉属昆虫,其中中国境内报道了9种。该类群是河流生态研究中的常见类群[20-24],但当前仍缺乏其生活史信息。因此,明确花翅蜉属的生活史性状,对于解释该类群分布所反映出的生态问题具有重要意义[12]。
本研究基于在三峡水库香溪河库湾的一条入库溪流中采集到的足够数量的麦氏花翅蜉(Baetiellamacani)个体,研究该种群的生活史,以填补花翅蜉属的生活史信息空白。
1 研究区概况
采样点位于汇入三峡水库香溪河库湾的一条入库溪流(螃蟹溪)中,如图1所示。水系图由GDEMV3 30 m分辨率数字高程数据经QGIS分析得到,高程数据下载自地理空间数据云(https://www.gscloud.cn)。样点A、B、C分别位于溪流的上、中、下游,海拔分别约为580、450、270 m。其中,上游靠近山脊区域存在少量次生林,中、下游两岸均为果园或农田。研究区域属于北亚热带气候,研究期间(2017年1—12月)的日平均气温在4.8~33.4 ℃之间,降雨集中出现在4—10月初期[11]。调查期间,螃蟹溪通常处于宽度较窄、流速慢的状态[12],河道内的主要基质类型为鹅卵石,而位于激流生境的鹅卵石基质正是花翅蜉属种类偏好的栖息地类型,这为本研究获取足够数量的麦氏花翅蜉样品提供了有利条件。同时,本研究所调查样点覆盖了麦氏花翅蜉在该溪流中的全部分布区域,以防止种群的空间移动对生活史判断造成影响。此外,调查期间,在现场用便携式水质测量仪(美国YSI,Professional Plus)对样点A、B、C的水质理化指标进行了测量。测量结果显示,水温、电导率和溶解氧在各个样点均存在季节性变化,而总溶解性固体、盐度和浊度均较为稳定,pH始终保持弱碱性状态[11]。
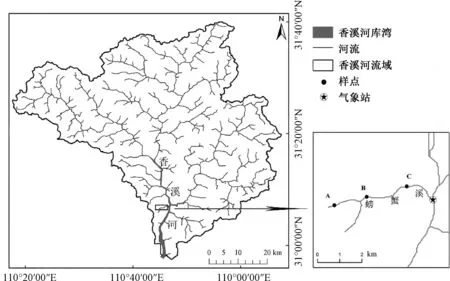
图1 研究区域及样点分布
2 数据来源与研究方法
2.1 野外调查和物种辨识
使用索伯(Suber)网(边长30 cm,孔径250 μm)对样点A、B、C进行为期一年(2017年1—12月)的大型底栖动物采集(A样点12月份数据缺失),在各样点逐月随机采集4~5个样品。其中,1月和4月由于河道较窄、大型底栖动物密度较高,仅采集了4个样品,而其他月份均采集了5个样品。现场用95%的酒精对采集到的样品进行保存。
根据麦氏花翅蜉新种描述文献[25]及相关分类文献[19],对所采集样品进行物种辨识。如图2所示,研究对象腹部Ⅲ到Ⅸ节具有成对的刺突,仅Ⅰ到Ⅵ节具有鳃片,Ⅲ节上的两个刺突的间距较小。

图2 麦氏花翅蜉老熟幼虫背面观
2.2 大小测量及生活史分析方法
根据由投影面积推算得到的个体半径,把麦氏花翅蜉幼虫分成10个组,从而构建用于确定其生活史的大小频率分布图,分组细节如表1所示。同时,图3呈现了各组的个体数量分布情况。利用OriginPro 8软件构建麦氏花翅蜉大小(投影面积)频率分布图,用翅牙变黑的高龄幼虫个体的出现时期指示成虫期,忽略雌雄个体大小差异的影响[12]。此外,由于蜉蝣成虫的寿命很短,用自成虫期开始到下一个世代的低龄幼虫首次出现所经历的时间段来代表卵期。各世代幼虫发育历期,即个体从孵化到羽化所经历的时间[29],可用从低龄幼虫首次出现到老熟幼虫羽化首次出现所经历的时间段来替代,或用从低龄幼虫末次出现到老熟幼虫羽化末次出现所经历的时间段来替代[30]。

表1 麦氏花翅蜉幼虫面积组的分组范围
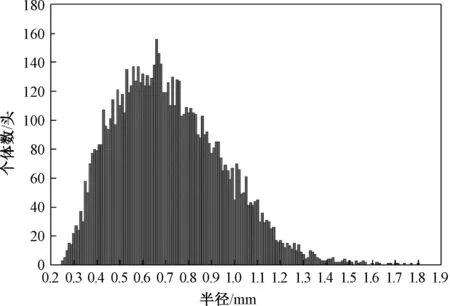
注:总个体数为7 974头。图3 麦氏花翅蜉幼虫的大小分布
2.3 有效积温计算方法
有效积温计算公式[12,31]如下:
TD=(Tmax+Tmin)/2-Tb
(1)
式中:TD为日有效积温,Tmax为每日最高温度,Tmin为每日最低温度,Tb为幼虫发育基础温度。麦氏花翅蜉幼虫的发育起点温度目前尚不清楚,但是通过观察发现,麦氏花翅蜉幼虫在整个冬天均未发生滞育现象,即使在接近0 ℃(1月)的极低水温下,麦氏花翅蜉幼虫也有较强的活动能力和明显的取食现象,因此,本研究假设麦氏花翅蜉幼虫发育的基础温度为0 ℃,并基于此计算了有效积温。调查期间样点水体的日最高温度和最低温度数据根据监测样点水温与气温的相关关系推算得到,具体可见LI等[12]的研究文献。
3 结果分析
螃蟹溪中的麦氏花翅蜉幼虫呈现出的逐月大小(投影面积)频率分布如图4所示,各样点用于分析生活史的具体个体数见表2。由图4可知,上游(A样点)、中游(B样点)、下游(C样点)的相同月份大小结构和成虫出现时间存在差异,主要体现在:上游样点的成虫首次出现时间是在4月,而中、下游样点均在2月;上游样点和中游样点的成虫羽化结束时间是在10月,而下游样点持续到了11月。

表2 各样点用于分析麦氏花翅蜉生活史的个体数量

注:各月份面积组的宽度表示该面积组的个体数占该月份总个体数的百分比,顶部的浅色方框表示成虫羽化期。图4 麦氏花翅蜉面积大小频率的逐月分布
综合样点A、B、C麦氏花翅蜉幼虫大小(投影面积)频率的逐月分布特征可知,麦氏花翅蜉幼虫全年在各样点河段均有分布。其中:低龄幼虫可区分出两个增补期,第一个出现在5—6月,第二个出现在7月到次年的2月;成虫也可区分出两个羽化期,首个出现在2—4月,第二个出现在6—10月。各世代辨识情况如下:
1)同时存在冬季世代和夏季世代。从幼虫大小结构变化和成虫羽化现象来看,4—5月存在明显的世代转变现象。从成虫羽化现象来看,11月前后存在明显的世代分离现象。因此,很容易得到麦氏花翅蜉同时存在冬季世代和夏季世代的结论。
2)存在两个夏季世代。从各样点的幼虫大小结构分析,5月份新增补的低龄幼虫个体在6月份就可羽化,即该世代幼虫的发育历期为1个月,其间需要的有效积温为757 ℃。基于该有效积温值,推测6月份增补的幼虫在7月份就能完成幼虫发育过程进入成虫期。因此,可确定5—6月增补的低龄幼虫个体的成虫羽化时间为6—7月,由此形成了一个完整的世代。同样,基于夏季世代幼虫发育需要的有效积温值(757 ℃),C样点在11月份出现的少量成虫的低龄幼虫增补时间为9月份,即9月份增补的低龄幼虫能在当年羽化,幼虫的发育历期为2个月,推导过程见图5。因此,可确定7—9月增补的低龄幼虫个体的成虫羽化时间为8—11月,由此构成了另一个完整的世代。
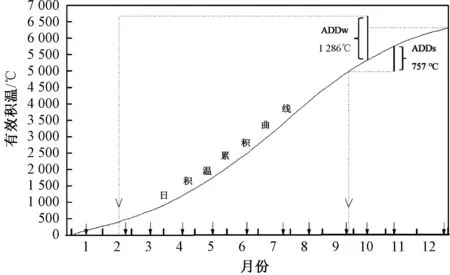
注:横轴箭头表示具体的采样时间,ADDs表示夏季世代幼虫发育的有效积温,ADDw表示冬季世代幼虫发育的有效积温,虚线用以指示满足有效积温条件的时间点。图5 麦氏花翅蜉幼虫发育的有效积温
3)存在一个冬季世代。冬季世代出现低龄幼虫的最晚时间为2月,羽化的最晚时间为5月,即冬季世代幼虫的发育历期为3个月,其间的有效积温为1 286 ℃。根据该有效积温值,推测10月份增补的个体将在次年的1月下旬羽化,幼虫的发育历期为4个月。该结果与实际调查结果一致,推导过程见图5。尽管冬季世代在各样点的出现时间存在差异,但是各样点第一个夏季世代低龄幼虫的增补时间均为5月份。
4)交替期及个体密度状况。整合上、中、下游3个样点的麦氏花翅蜉各面积组的个体数量(图6)发现:冬季世代和第一个夏季世代的交替期在3—5月,呈现可辨识状态;两个夏季世代的交替期无法直观辨识;在第二个夏季世代与冬季世代的交替期,仅高龄个体可直观辨识。麦氏花翅蜉的个体密度呈现出两个高峰期,分别在1月和9月(图7)。
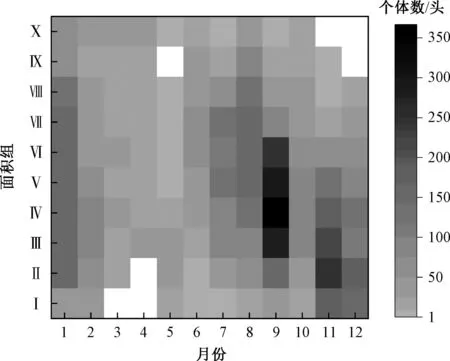
注:白色单元格表示个体数为0。图6 麦氏花翅蜉幼虫各面积组个体数的逐月分布
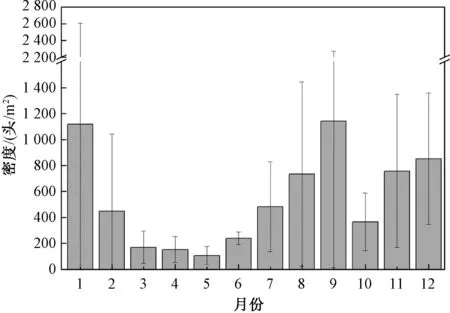
图7 麦氏花翅蜉幼虫个体密度的逐月分布
综上,最终确定麦氏花翅蜉为三化性昆虫,包含一个冬季世代和两个夏季世代。冬季世代的低龄幼虫增补期是从10月持续到次年的2月,成虫羽化期从2月持续到5月,幼虫发育历期为3~4个月,需要的有效积温为1 286 ℃,并且与第二个夏季世代的幼虫存在明显的世代重叠现象。夏季世代需要的有效积温为757 ℃。第一个夏季世代的低龄幼虫增补期为5—6月,成虫羽化期为6—7月,幼虫发育历期为1个月。第二个夏季世代的低龄幼虫增补期为7—9月,成虫羽化期为8—11月,幼虫发育历期为1~2个月。两个夏季世代间呈现严重的世代重叠现象。
4 讨论
目前,研究人员仅掌握少量国内水生昆虫生活史信息的现状,阻碍了解释水生昆虫及大型底栖动物群落发生动态变化的原因、监测和评估相关变化对水生态系统的影响等相关研究的进展,导致国内研究者在相关研究中主要借鉴北美地区的研究结果[32-35]。本研究获得了麦氏花翅蜉在三峡水库香溪河库湾入库溪流(螃蟹溪)中的生活史信息和幼虫生长发育需要的有效积温条件,在丰富中国水生昆虫生活史信息的同时,也首次呈现了花翅蜉属昆虫的生活史性状信息。麦氏花翅蜉在华中地区为三化性昆虫,幼虫在全年均有分布,与体型同样较小的光滑细蜉(Caenislubrica)在该地区的世代数[12]一致,比该区域体型较大的种类具有更多的世代数,如梧州蜉(Ephemerawuchowensis)为一年一代[11],三脊弯握蜉(Drunellasubmontana)为一年一代或两年一代[4]。
花翅蜉属是一类体型较小、偏好激流生境的类群[21],其种群的维持需要稳定的流水环境。尽管螃蟹溪上、中、下游间的海拔落差较小,但是上、中、下游样点的相同月份龄期结构和成虫出现时间存在明显差异。这可能是由于上游样点激流生境不稳定,且花翅蜉属具有漂流习性[20,23-24],从而造成上游高龄幼虫较少。尽管麦氏花翅蜉冬季世代成虫在溪流上、中、下游的羽化时间存在明显差异,但是低龄幼虫统一出现在5月份,因此,推测麦氏花翅蜉虫卵的孵化需要达到一个相对较高的温度条件,这可能也对麦氏花翅蜉在华中地区的分布造成了一定影响。此外,基于在香溪河流域进行的大型底栖动物长期调查发现,麦氏花翅蜉仅分布在低海拔区域,而该地区低海拔区域河流的自然生境受到小水电站的严重干扰[23]。
5 结论
在三峡水库香溪河库湾一条入库溪流(螃蟹溪)中,麦氏花翅蜉幼虫在全年均有分布,为一年三代,即属于三化性水生昆虫,包括一个冬季世代和两个夏季世代。3个世代间呈现明显的世代重叠现象,其中:冬季世代的幼虫发育历期为3~4个月,所需达到的有效积温为1 286 ℃;夏季世代的幼虫发育历期为1~2个月,所需达到的有效积温为757 ℃。研究结果丰富了中国水生昆虫生活史性状信息,并首次呈现了花翅蜉属昆虫的生活史信息。
