中华鳖源按蚊伊丽莎白菌的分离鉴定及药敏特性研究
2024-03-02王玉柱蒋巍巍刘文舒路晶晶唐艳强肖海红郭小泽李思明
王玉柱 蒋巍巍, 刘文舒 路晶晶 唐艳强 肖海红 郭小泽 李思明
(1. 江西省农业科学院畜牧兽医研究所, 南昌 330200; 2. 四川农业大学动物科技学院, 成都 611130)
中华鳖(Trionyx sinensis), 隶属于龟鳖目(Tesudines), 鳖科(Trionychidae), 鳖属(Trionyx)。作为一种重要的水产养殖品种, 中华鳖在我国湖南、湖北、江西、江苏等多省份均有分布, 全国商品鳖年产量达37万吨以上[1]。近年来, 随着中华鳖养殖规模的扩大和集约化养殖模式的推广, 养殖密度过高、生产管理和病害防治技术落后等问题引发中华鳖细菌性疾病频频暴发[2], 给养殖户带来巨大经济损失。目前, 中华鳖常见的细菌性疾病有嗜水气单胞菌(Aeromonas hydrophila)、弗氏柠檬酸杆菌(Citrobacter freundii)、迟缓爱德华菌(Edwardsiella tarda)等致病菌引起的败血症、腐皮病、疥疮病、白底板病等病症[3—6], 然而, 在中华鳖病害的研究过程中, 仍不断有致病菌的新型症状和新型致病菌被发现[7,8]。
按蚊伊丽莎白菌(Elizabethkingia anophelis), 隶属于黄杆菌科(Flavobacteriaceae), 伊丽莎白菌属(Elizabethkingia); 2011年, Peter Kämpfer等[10]首 次从冈比亚疟蚊肠道中分离鉴定并命名[9]。按蚊伊丽莎白菌被认为是条件致病菌, 自该菌被发现以来,在中国、柬埔寨、丹麦、美国等全世界多个国家和地区报道了其感染人类的案例[11—15], 能够引起人类败血症、脑膜炎等疾病。值得注意的是, 按蚊伊丽莎白菌对多种抗生素具有较强耐药性[16], 并且根据基因组学分析, 其具有潜在的大规模暴发能力和全球快速传播能力[17]。
2021年10月, 江西省抚州市南丰县某中华鳖养殖基地暴发鳖大规模传染性病害, 症状表现为行动迟缓, 颈部肿大, 有腹水, 并伴有肠道出血, 严重者可致死。为探究该中华鳖传染病的病原, 我们以濒死中华鳖作为样本, 从中华鳖肝脏、脾脏中分离出一株优势菌群, 经人工回归感染试验证实其具有致病能力。对该致病菌进行形态观察、生理生化鉴定和16S RNA基因序列测定, 认定该分离株为按蚊伊丽莎白菌。进一步, 我们对该致病菌进行了药敏试验和全基因组测定, 并对其毒力因子进行了分析,以期为中华鳖养殖过程中预防和治疗相关病害提供参考。
1 材料与方法
1.1 实验动物
患病中华鳖和人工回归感染试验用鳖从江西省抚州市南丰县某中华鳖养殖基地获得。
1.2 致病菌分离纯化
用75%浓度酒精对濒死病鳖进行体表消毒, 在无菌环境下解剖, 取部分肝脏、脾脏组织磨成匀浆,用灭菌棉签蘸取组织匀浆, 划线接种在LB固体培养基(青岛海博生物技术有限公司)上。将培养基封口膜密封后, 转移至28.5℃恒温培养箱中培养24h。在培养结束后, 用灭菌接种环挑取培养基中优势菌落的单菌落, 接种至新的LB培养基上, 再次在28.5℃恒温培养箱中培养24h, 连续纯化3次后, 得到目标菌株EJX2021。现保存于江西省农业科学院畜牧兽医研究所和中国典型培养物保藏中心(武汉),保藏编号为CCTCC NO: M 2022507。
1.3 分离株的鉴定
形态学观察将冻存EJX2021菌株室温复苏, 在无菌环境下进行LB培养基划线培养, 封口膜密封后转移至28.5℃恒温培养箱中培养24h。选定独立菌落进行观察, 记录菌落的形状、大小、颜色、表面特征, 透明度等表型特征。用灭菌接种环将单菌落挑至灭菌LB液体培养基中过夜培养。将部分菌液送至武汉塞维尔生物科技有限公司进行革兰氏染色观察和扫描电镜观察。
16S RNA基因序列测定将上文部分菌液3000 r/min离心5min, 后弃上清, 只保留细菌沉淀。用无菌PBS吹打重悬, 再次离心。重复2遍后, 收集细菌沉淀, 移送至北京擎科(湖南)生物科技有限公司进行16S RNA基因序列测定。将测定序列在NCBI数据库(https://blast.ncbi.nlm.nih.gov/Blast.cgi)中进行Nucleotide BLAST对比检索, 并利用MEGA 7软件邻接法构建系统发育树。
生理生化特性测定生理生化特征指标选定参考《常见细菌系统鉴定手册》。生理生化鉴定管采购自青岛海博生物技术有限公司, 具体操作方法按照说明书进行。
1.4 人工回归感染实验
人工回归感染试验选取50只幼鳖(50±5) g, 随机分为5组, 每组10只, 置于暂养箱(72 cm×53 cm×42 cm)内。将EJX2021菌株活化后利用“麦氏比浊法”将菌液稀释为2×107、1×108、5×108和2.5×109CFU/mL待用, 分别给5组鳖注射200 μL的各浓度稀释菌液和无菌生理盐水, 连续观察15d。每天观察并记录中华鳖的死亡情况, 并对濒死病鳖进行组织切片观察和半数致死剂量(LD50)计算。
1.5 组织病理学观察
在1.2和1.4步骤基础上, 对健康鳖和人工回归感染试验濒死病鳖进行解剖, 取肝、脾、肺、肠组织后分别放入4%多聚甲醛溶液固定, 移送武汉塞维尔生物科技有限公司进行组织HE染色观察。
1.6 毒力因子分析
将上文收集的部分细菌沉淀移送至深圳微科盟科技集团有限公司进行全基因组测序。应用CAMS&PUMC病原生物学研究所毒力因子数据库(Virulence factors of pathogenic bacteria, VFDB)(http://mgc.ac.cn/cgi-bin/VFs/v5/main.cgifunc=VFanalyzer)的VFanalyzer在线工具对毒力因子进行筛选。
1.7 抗生素药物敏感性实验
选取23种常规抗生素用常规纸片扩散法(Kirby-Bauer)进行药物敏感性试验。在无菌条件下, 将分离株菌液调制成1个麦氏单位浓度, 用涂布器将200 μL菌液均匀涂布在LB培养皿(90 mm)上, 每个培养皿贴放3片药敏纸片(采购自杭州微生物试剂有限公司)。用封口膜将培养皿封口后倒置在恒温培养箱中28.5℃培养24h, 培养结束后测量抑菌圈直径。
2 结果
2.1 菌株的鉴定
形态学观察分离株在LB营养琼脂平板上28.5℃培养24h后, 菌落形态如图1A所示。分离株形态呈现白色或微黄色椭圆形, 表面湿润、光滑,中间隆起, 边缘整齐, 呈半透明状, 直径1—2 mm(图1A)。经过革兰氏染色后, 分离株呈现红色长杆状, 判定革兰氏阴性菌(图1B)。通过扫描电镜观察, 菌体直径约0.65 μm, 成熟个体长度约1.45—1.65 μm, 两端钝圆, 菌毛密集, 无鞭毛( 图1C)。
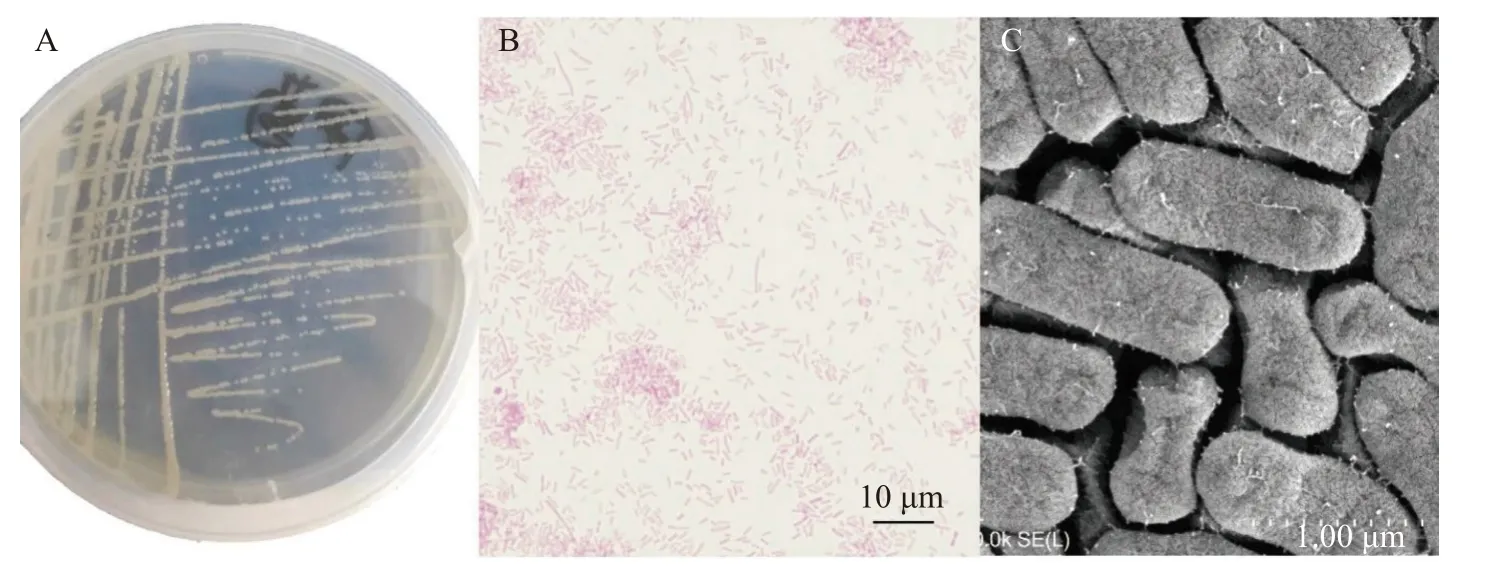
图1 分离株菌落形态、革兰氏染色及扫描电镜结果Fig. 1 Colony morphology, Gram staining and SEM results of the isolates
16S RNA基因序列测定测序结果表明,EJX2021菌株16sRNA基因片段大小为1282 bp (Gen-Bank: OL989210.1)。基因序列在NCBI数据库中的Nucleotide BLAST对比检索结果显示, 其与按蚊伊丽莎白菌的16S RNA基因序列(GenBank: AP022313.1、CP046080.1和CP071545.1)相似度为99.92%。选取部分与EJX2021菌株BLAST对比检索结果相似的菌株和常见水产致病菌株基因序列, 利用MEGA7软件对EJX2021菌株构建系统进化树(图2)。结果显示, EJX2021菌株与按蚊伊丽莎白菌同属一个分支。

图2 EJX2021菌株基于16S RNA基因序列的系统进化树Fig. 2 Phylogenetic tree of EJX2021 strain based on 16S RNA gene sequence
生理生化特性测定EJX2021菌株的生理生化特征如表1所示。按蚊伊丽莎白菌的生理生化指标在《常见细菌系统鉴定手册》等鉴定手册中无记录, 因此EJX2021菌株生理生化特征指标的选择参考黄杆菌属细菌鉴定指标进行测定。结果显示:糖(醇、苷)发酵试验中, EJX2021菌株对X-甲基-D-葡萄糖苷、卫矛醇半固体呈阳性反应; 对D-阿拉伯糖醇、D-甘露糖、D-山梨醇、L-鼠李糖、阿东醇、阿拉伯糖、半乳糖、甘露醇、海藻糖、肌醇、麦芽糖、蜜二糖、木糖、乳糖、葡萄糖、水杨苷、松三糖、纤维二糖、蔗糖、淀粉等均呈阴性反应。在明胶和甘油生化试验中, EJX2021菌株可使明胶生化管呈阳性, 甘油生化试验呈阴性。此外, VP (伏普试验)、ONPG (β-半乳糖苷酶试验)、硫化氢试验、赖氨酸脱羧酶、鸟氨酸脱羧酶、氧化酶、苯丙氨酸脱氨酶、丙二酸盐、唾液酸盐等测定均呈阴性, 硝酸盐、尿素酶测定呈阳性。

表1 EJX2021菌株的生理生化鉴定Tab. 1 Physiological and biochemical identification of EJX2021 strain
2.2 人工回归感染实验
如图3所示, 中华鳖腹腔注射不同浓度EJX 2021菌株后, 2×107、1×108、5×108和2.5×109CFU/mL注射菌液浓度的15d死亡率分别为10%、40%、70%和100%。2.5×109CFU/mL组在第11天时, 鳖全部死亡。利用改良寇氏法[18]计算EJX2021菌株LD50(表2), LD50=lg-1(Xk-i(∑p-0.5))=1.62×108CFU/mL。95%置信区间为lg-1{[Xk-i(∑p-0.5)]±1.96×108CFU/mL。即EJX2021菌 株 在15d内 的LD50为1.62×108CFU/mL, 95%置信区间为(0.78—3.37)×108CFU/mL。

表2 EJX2021菌株回归感染试验结果Tab. 2 Results of EJX2021 strain regression infection test

图3 不同浓度EJX2021菌株感染中华鳖死亡曲线Fig. 3 Death curves of Trionyx sinensis infected with EJX2021 strain at different concentrations
2.3 组织病理学观察
如图4所示,经EJX2021菌株回归感染后, 鳖肠道、肺脏、脾脏、肝脏等组织出现不同程度病变。回归感染后肠管组织发生肿胀, 肠壁明显变厚;肠绒毛变化表现在杯状细胞增多, 肠上皮柱状细胞排列松散, 细胞纹状缘不明显或缺失。回归感染的肺组织可见肺泡壁明显纤维化, 肺泡壁上皮细胞的细胞核溶解或消失, 肺泡腔内有炎症渗出物。回归感染的脾脏组织可观察到巨噬细胞数量增多, 细胞质空泡变性, 细胞核核仁减少; 淋巴细胞体积增大,胞核淡染。回归感染肝脏组织的肝小叶中央静脉可见瘀血, 个别肝细胞发生坏死。

图4 EJX2021菌株感染中华鳖组织病理学变化(HE染色)Fig. 4 Histopathological changes of Trionyx sinensis infected with EJX2021 strain (HE staining)
2.4 毒力因子分析
如表3所示,预测毒力基因共246个, 相关毒力因子分类包括黏附(Adherence)、分泌系统(Effector delivery system)、外毒素(Exotoxin)、胞外酶(Exoenzyme)、荚膜(Capsule)、运动性(Motility)、营养和代谢(Nutritional/Metabolic factor)、抗逆性(Stress survival)、竞争优势(Antimicrobial activity/Competitive advantage)、信号调控(Regulation)、炎症通路(Immune modulation; Inflammatory signaling pathway)、生物膜生成(Biofilm formation)、免疫调节(Immune modulation)、入侵(Invasion)、补体逃逸(Complement evasion/Serum resistance)和其他等16大类。

表3 EJX2021菌株毒力基因预测Tab. 3 Virulence gene prediction of EJX2021strain
2.5 药物敏感性实验
如表4所示,在23种抗生素药物中, EJX2021菌株对6种敏感(多西环素、米诺环素、卡那霉素、哌拉西林、恩诺沙星和氟苯尼考), 2种中度敏感(头孢曲松和头孢哌酮), 其余15种耐药。
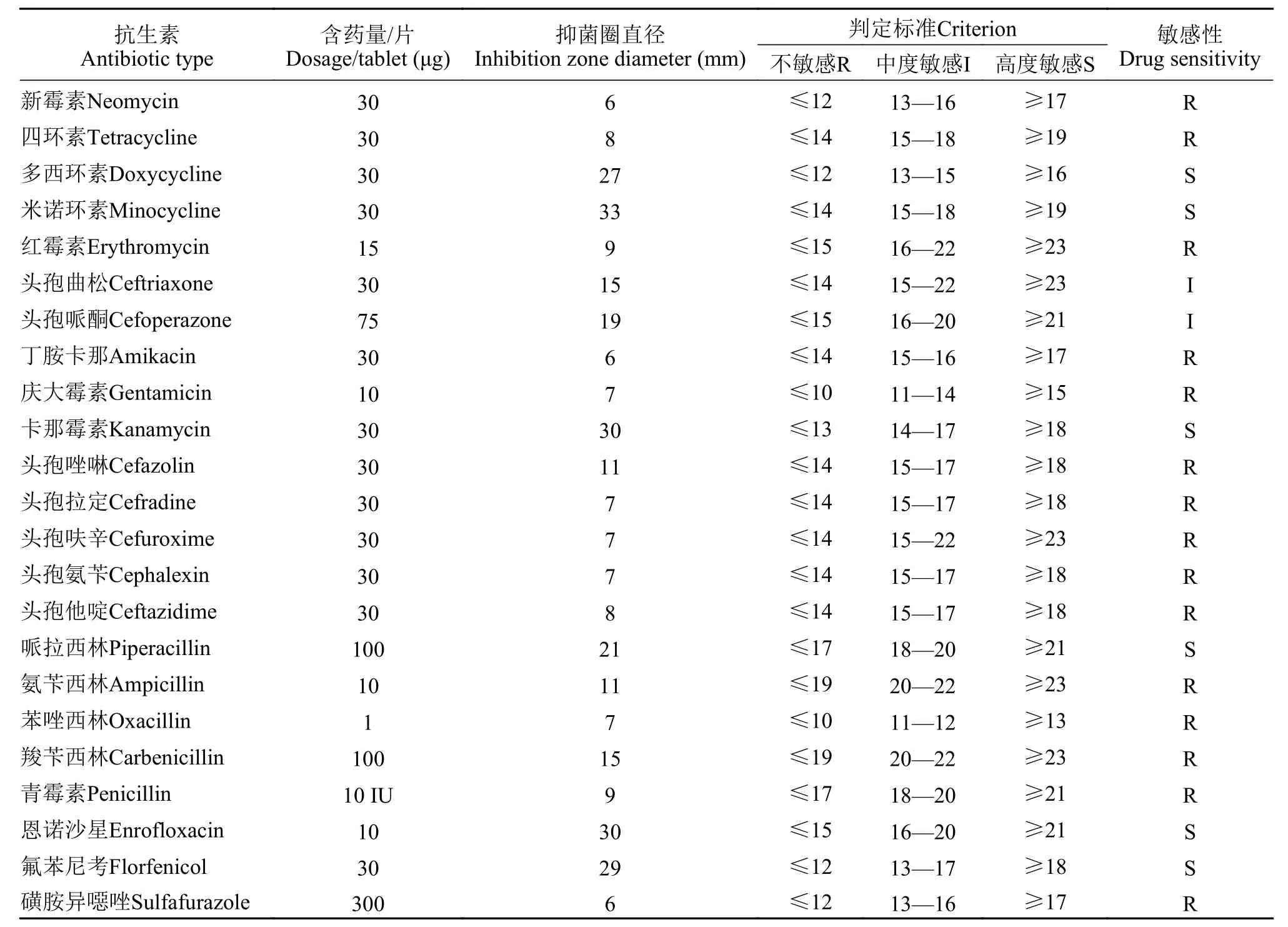
表4 23种抗生素对EJX2021菌株的敏感性试验结果Tab. 4 Sensitivity test results of 23 antibiotics to EJX2021strain
3 讨论
3.1 致病菌的分离与鉴定
伊丽莎白菌属细菌形态学特征明显, 通常呈两端钝圆的杆状, 大小约0.5 μm×(1.0—2.5) μm, 革兰氏染色呈阴性; 菌落形态特征呈圆形隆起、半透明或透明。边缘清晰, 表面光滑[19]。在本试验中, EJX 2021菌株与伊丽莎白菌属细菌形态学特征一致。伊丽莎白菌属设立较晚, 现包含了6个菌种。分别是Kim等[19]在2005年从黄杆菌属(Chryseobacterium)中重新分类的米尔伊丽莎白菌(Elizabethkingia miricola)和脑膜脓毒性伊丽莎白菌(Elizabethkingia meningoseptica), Kämpfer等[20]在2011年发现的按蚊伊丽莎白菌[9]和2015年发现的Elizabethkingia endophytica(后 经 证 实,Elizabethkingia endophytica归类于按蚊伊丽莎白菌[21]), Nicholson等[22]在2018年 发 现 的Elizabethella bruuniana、Elizabethella ursingii和Elizabethella occulta。《常见细菌系统鉴定手册》[23]《伯杰氏细菌鉴定手册》[24]等快速鉴定手册成书早于伊丽莎白菌属的设立, 因此, 参照细菌生理生化特征作为按蚊伊丽莎白菌的鉴定手段有较大局限性。本试验参考黄杆菌属等细菌生理生化特征鉴定指标, 对EJX2021菌株进行部分生理生化指标鉴定, 结果显示其对多数糖、苷、醇类物质不能利用, 对X-甲基-D-葡萄糖苷、卫矛醇等可利用; 可利用明胶, 不可利用甘油; 此外, EJX2021菌株还可分解硝酸盐、产生尿素酶。细菌的16S RNA基因片段的保守性很高, 且在不同种间存在相对变异区, 在微生物鉴定时具有快速、高效和准确的特点[25]。所以, 测定16S RNA基因序列和构建系统发育树的方法成为微生物鉴定的常用手段。在本试验结果中, EJX2021菌株和6株按蚊伊丽莎白菌位于系统进化树的同一分支(Bootstrap值为94), 证明EJX2021菌株和按蚊伊丽莎白菌属于同一物种。因此, 通过形态学和分子生物学鉴定, 确认EJX2021菌株为按蚊伊丽莎白菌。
按蚊伊丽莎白菌广泛分布在自然环境中[26], 自从2011年发现以来, 其持续被报道引起肺炎、菌血症、新生儿脑膜炎等人类临床医学病症[27,28], 被认为是医院等环境中的重要条件致病菌[29]。在水产养殖中, 伊丽莎白菌属的米尔伊丽莎白菌和脑膜脓毒性伊丽莎白菌导致蛙类等水产动物病害的报道屡见不鲜[30,31], 但是按蚊伊丽莎白菌感染水产动物的现象没有报道。本研究首次发现了按蚊伊丽莎白菌感染中华鳖。在早期按蚊伊丽莎白菌属的研究中, 对其生理生化鉴定手段缺乏, 基因序列对比已有伊丽莎白菌属基因序列和相关报道, 往往引起伊丽莎白菌属物种菌种鉴定的不准确判断[32], 这可能是按蚊伊丽莎白菌在水产动物上报道较少的原因之一。
3.2 EJX2021菌株感染中华鳖组织病理学观察
组织病理学观察能够直接、准确地反映器官、组织和细胞的病理学变化, 是研究自然发生和实验诱导疾病形态特征和病变程度的最可靠、敏感和全面的测定方法之一[33]。关于按蚊伊丽莎白菌引起的动物机体病理学观察报道较少, 本试验观察显示, 按蚊伊丽莎白菌可造成中华鳖肠道、肺脏、脾脏和肝脏等组织器官不同程度的病理损伤。水产动物的细菌性感染往往呈现综合性病症,会引起肝、肠等部位细胞变性、组织崩溃、形成炎症[34]。肠道、肺脏、肝脏、脾脏等是机体重要的免疫和代谢器官, 持续感染造成的组织器官渐进性损伤, 可能会引起中华鳖的呼吸障碍、营养吸收障碍、体液循环障碍等功能性障碍, 严重可致死[35]。按蚊伊丽莎白菌引起的组织器官病变可能是导致人工回归感染中华鳖死亡的原因之一, 有待进一步验证。
3.3 EJX2021菌株的致病性
中华鳖致病菌种类繁多, 致病力各不相同。前人实验验证, 苏云金芽孢杆菌(Trionyx sinensis)在10—15 g中华鳖上的LD50为1×104.87—105.30CFU/mL[36], 蜡样芽孢杆菌(Bacilluscereus)在200 g中华鳖上为4.91×106CFU/mL[37], 迟钝爱德华氏菌(Edwardsiella tarda)在150 g中 华 鳖 上 为2.45×106CFU/mL[38], 摩氏摩根菌(Morganella morganii)在4—6 g中华鳖上为1×107.24CFU/mL[39], 奇异变形杆菌(Proteus mirabilis)在150 g中华鳖上为1×107.83CFU/mL[40],嗜水气单胞菌(Aeromonas hydrophila)在500 g中华鳖上为1.5×108CFU/mL[41], 弗氏柠檬酸杆菌(Pelodiscus sinensis)在30 g中华鳖上为9.3×104CFU/g[6]。在本试验中, 按蚊伊丽莎白菌在50 g中华鳖上的LD50为1.62×108CFU/mL。与上述常见致病菌相比,本研究中按蚊伊丽莎白菌对中华鳖的致死能力较弱。但是, 按蚊伊丽莎白菌不同亚种在不同地域环境中的毒性还缺乏研究支撑, 其对水产动物的潜在危险性仍不容忽视。
自从2011年被发现以来, 按蚊伊丽莎白菌在全球至少有4次以上的中到大规模暴发, 然而, 人们对按蚊伊丽莎白菌的认识仍不够全面[42]。利用全基因组学对按蚊伊丽莎白菌毒力因子进行预测有助于我们深入了解和解释其致病机制和临床表现。Perrin等[43]对美国威斯康星州在2015—2016年暴发的细菌感染中分离的按蚊伊丽莎白菌样本进行毒力因子预测, 结果显示人源按蚊伊丽莎白菌株含有67个毒力基因, Teo等[44]在新加坡暴发疫情的按蚊伊丽莎白菌中预测得到关于脂多糖合成、铁载体合成、血红素摄取等分类毒力基因共216个。本试验对比VFDB数据库, 预测得到按蚊伊丽莎白菌的毒力基因246个, 共16类。前人研究结果显示按蚊伊丽莎白菌的毒力因子涉及荚膜构成、脂多糖和脂质生物合成代谢、转运蛋白、分泌系统、应激反应等多个方面[45]。细菌荚膜是位于细胞壁外的一层物质, 对细菌在动物机体中的依附、入侵和免疫逃避等有重要影响, 是引发脑膜炎等病症的主要细菌毒力因素[46]。Perrin等[43]在威斯康星州样本中发现了5类荚膜蛋白合成有关的基因, 本研究中发现了25个荚膜相关毒力基因。另外, 本试验发现按蚊伊丽莎白菌中检测到的部分毒力因子是许多致病菌的共同特征。例如,katG和zmp1因子能够参与应激反应和结核分枝杆菌(Mycobacterium tuberculosis)的发病过程[47,48],IlpA诱导产生的细胞因子在创伤弧菌引起的败血病中起重要作用[49],clpP、tuf、rmla和dnak等毒力基因在一种拟香味类香味菌(Myroides odoratimimus)中可能参与了机体的入侵和防御等步骤[50]。同时, 我们还在按蚊伊丽莎白菌的毒力基因中发现了巨噬细胞感染和存活所需要的Mip蛋白[51]编码基因。本研究对中华鳖源按蚊伊丽莎白菌毒力因子的预测显示, 该菌株具有比人类临床按蚊伊丽莎白菌更多且与人类致病菌高度重合的毒力因子群体, 表明中华鳖源按蚊伊丽莎白菌可能具有人类致病潜力。
3.4 EJX2021菌株的药敏特性
水产养殖过程中抗生素的不合理使用往往导致水体致病菌出现广泛耐药性[52]。为检验EJX2021菌株的耐药特征, 本试验采用23种常见抗生素进行药敏试验, 结果显示EJX2021菌株对四环素类的多西环素、米诺环素抗生素敏感, 对氨基糖苷类的卡那霉素敏感, 对β-内酰胺类的哌拉西林敏感, 对喹诺酮类的恩诺沙星和氟苯尼考敏感; 对头孢菌素类的头孢曲松和头孢哌酮中度敏感; 对其他15种抗生素不敏感。本试验结果与Pavel等[53]、王小龙等[54]的研究有较大差异, Pavel等[53]在俄罗斯产牛奶中分离出的按蚊伊丽莎白菌对哌拉西林、卡那霉素均不敏感; 王小龙等[54]在苏州地区肺部感染者体内分离得到的按蚊伊丽莎白菌对头孢曲松、头孢哌酮、哌拉西林敏感, 对米诺环素中度敏感, 对卡那霉素、多西环素不敏感。上述结果说明EJX2021菌株为多重耐药菌株, 且在不同环境中筛选出了耐药性差异; 并且, 随着按蚊伊丽莎白菌新品系不断产生[55], 其耐药性可能随之增强。因此, 根据药敏试验结果判定按蚊伊丽莎白菌的治疗方法显得尤为重要。
