云南短头熊蜂体色模式多态性地理分布格局
2024-02-26秦加敏宗德琴李雨时丁桂玲黄家兴
梁 铖, 秦加敏, 宗德琴, 李雨时, 丁桂玲, 黄家兴
(1. 云南省农业科学院蚕桑蜜蜂研究所, 蒙自 661101;2. 中国农业科学院蜜蜂研究所, 农业农村部授粉昆虫生物学重点开放实验室, 北京 100093)
熊蜂Bombusspp.是众多野生植物和农作物的传粉昆虫类群之一,在自然生态系统和农业生产中均发挥着重要作用(Velthuis and van Doorn, 2006; Goulsonetal., 2008)。全球熊蜂约有260种,其分布广泛,主要分布在北半球温带地区(Williamsetal., 2017; Cameron and Sadd, 2020)。中国地形多样、植被丰富,是全球熊蜂多样性的热点地区。目前已确认中国有125种熊蜂,是全球熊蜂物种资源最为丰富的国家(Williamsetal., 2017; 黄家兴和安建东, 2018)。熊蜂的一个突出特征是其体表通常覆盖着较长并且颜色鲜亮的毛,不同体节的体毛颜色多变,从而形成丰富的体色模式(Williams, 2007)。研究发现,许多熊蜂种具有较高的种内体色模式多态性,体色模式的进化速率甚至超过了线粒体基因(Duennesetal., 2012; Hines and Williams, 2012; Huangetal., 2015; Dingetal., 2019)。同时,不同熊蜂种之间还会具有相似的体色模式(明亮熊蜂复合种B.lucorumcomplex: Carolanetal., 2012; 刘苹等, 2014),较多的复合种或多变种类群的存在影响了熊蜂物种的准确鉴定,从而给熊蜂资源的深入研究造成了一定障碍。
研究表明,不同的熊蜂种可能具有相同的体色模式,特别是在同一区域分布的不同种熊蜂之间常具有相似的体色模式。调查发现分属于不同亚属、亲缘关系较远的三条熊蜂B.trifasciatus、红尾熊蜂B.haemorrhoidalis、圆头熊蜂B.rotundiceps和短头熊蜂B.breviceps均具有多个体色模式,但其体色模式呈现出趋同性,均随分布区不同而共同变化(Williams, 1998; Hines and Williams, 2012)。这种相同地理分布区域内不同熊蜂物种之间体色模式的趋同以及不同区域间熊蜂体色模式的并行共变被认为可能是缪氏拟态(Müllerian mimicry)的结果(Plowright and Owen, 1980)。基于对全球219种熊蜂的632头工蜂的体色模式分析结果,Williams(2007)认为3种主要的体色模式类群与3种可能的功能相关:(1)体色最黑的熊蜂主要分布在热带地区,这种体色可能与体温调节相关;(2)体色较浅的熊蜂类群出现在北部中纬度地区,这种体色在干燥的草原中有隐蔽的作用;(3)具有鲜明条带的熊蜂类群分布广泛,这种体色类群在其分布占优势的区域可能作为对熊蜂捕食者发出的集体警告信号,即缪氏拟态。分布于东南亚地区的三条熊蜂、红尾熊蜂和短头熊蜂有14个体色类群相互模仿,是研究熊蜂缪氏拟态进化的典型范例。对这3种熊蜂的体色模式比较后发现,不同拟态类群间拟态的程度和地理范围存在较大差异,并且体色模式在一个区域内形成之后,会受到正选择压力,环境条件对熊蜂的体色影响很大(Hines and Williams, 2012)。
短头熊蜂主要分布于喜马拉雅山山脉西南麓及中国长江流域以南的东南亚地区(Williamsetal., 2009),是众多野生植物和农作物的重要传粉者,年活动周期长,群势大,工蜂数量可超过300头,是我国具有较大农业授粉应用潜能的一种本土传粉熊蜂资源(Liangetal., 2020)。再者,短头熊蜂在云南分布的36种熊蜂中,分布广、数量多,是优势种(梁铖等, 2018),也是具有较高体色多态性的典型蜂种,种内体色模式多样。Hines和Williams(2012)报道短头熊蜂有13种体色模式,但该研究在云南的采样点较少,标本数量也较少,不能全面反映该蜂种的体色模式多态性及其在云南的分布格局。云南省位于中国西南部,地处青藏高原东南部延伸区和云贵高原主体部分,境内气候、地形和植被类型多样,是全球生物多样性研究的热点地区之一(杨宇明等, 2008; Lietal., 2015)。在熊蜂生物地理学方面云南西北部横断山区属于古北区,而云南高原及南部山地河谷属于东洋区,物种交叉过渡现象特征明显(Williams, 1996; 梁铖等, 2018)。本研究通过广泛采样和深入检视云南短头熊蜂体色模式,解析其体色模式多态性以及地理分布格局,不仅可以为短头熊蜂的遗传多样性研究和资源开发利用奠定基础,还可以为熊蜂的体色遗传与环境适应性研究提供更清晰的参考资料。
1 材料与方法
1.1 样品采集和鉴定
2017-2021年的6-9月于云南省各地、2021年8月在贵州省贵阳市收集熊蜂样品。采集样品时,利用手持GPS(Garmin 60CS)记录每个采样点的海拔以及经纬度等信息。采集的样品制作成针插标本保存在云南省农业科学院蚕桑蜜蜂研究所标本室。参考Williams等(2009)报道的方法,对采集的标本进行鉴定,共鉴定出短头熊蜂957头(云南:919头;贵阳:38头),包括雌成蜂898头(蜂王103头、工蜂795头)以及雄成蜂59头,这些短头熊蜂标本共来自于127个采样点,使用Arc GIS 10.2绘制采样分布图(图1)。
1.2 成蜂体色模式分析
参照Williams(2007)对熊蜂成蜂体色模式的描述方法(图2),增加了胸部中间体毛色块形状(黑色长方形色块、黑色色块较宽趋于正方形、黑色色块趋于圆形)以及雌成蜂腹部第6背板和雄成蜂腹部第7背板的体毛颜色的检视,按照体节组成把雌成蜂和雄成蜂的整个背面分为不同的部分,分别检视记录957头短头熊蜂的胸、腹部背面各个部分的体毛颜色及胸部背面中间色块形状,绘制短头熊蜂的体色模式图谱。

图2 将熊蜂背板划分为不同部分进行体色检视 (改自Williams, 2007)
采用贝格-派克(Berger-Parker)指数D分析短头熊蜂雌成蜂体色模式的优势度(丁岩钦, 1994),计算公式为:D=Ni/N,式中Ni为表现为第i种体色模式的标本数量,N为雌成蜂标本的总数量。依据正态分布的原则,D≥0.10时为优势体色模式,0.05≤D<0.10时为丰盛体色模式,0.01≤D<0.05时为常见体色模式,D<0.01时为稀有体色模式。
基于整理绘制的雌成蜂体色模式,把胸部中间色块形状以及腹部背板每个组成部分的体毛颜色类型转换为数值,使用SPSS 22.0进行系统聚类分析,获取近似值矩阵,导入MEGA 7.0绘制环形聚类图,对短头熊蜂体色模式进行聚类。
1.3 雌成蜂体色模式的地理分布格局分析
根据短头熊蜂雌成蜂体色模式的聚类结果,结合相应标本采集点的地理信息,以20.50°N, 97.00°E为起点,将云南全境划分为175个经纬度单元格,每个单元格大小为0.50°×0.50°。统计短头熊蜂的体色模式在每个单元格内的分布情况,有分布时标记为“1”,无分布时标记为“0”,建立二元数据列联表,利用SPSS 22.0软件,选用组间连接法结合曼哈顿街区距离(city block distance)进行系统聚类,以聚类在一起的单元格边线为分界线,使用Arc GIS 10.0绘制体色模式的地理分布图,其中没有样本分布记录的方格根据地理和气候特征合并到相邻区域。利用SPSS 22.0软件,使用偏相关法分析单元格内体色模式多态性的丰富度与其分布地经纬度的相关性;对体色模式的分布地海拔进行单因素方差分析,基于分析结果利用GraphPad Prism 8.0.2绘图。
2 结果
2.1 短头熊蜂的体色模式多态性
短头熊蜂胸部背板体毛的颜色有黑色、灰黄色和橙黄色3种,腹部背板体毛颜色有黑色、灰黄色、黄色和深棕色4种,所有个体腹部第1背板的体毛均为黄色,而其他体节的整节或局部体毛颜色存在变化,表现为黄色体毛由腹部第1背板向后逐节延伸最多至第3背板的整节,第2和3背板的体毛颜色为黄色、黑色或灰黄色,而深棕色体毛由腹部第6背板向前逐节延伸最多至第3背板的下半节,雌成蜂第6背板或雄成蜂第7背板的体毛为深棕色或黑色,蜂王、工蜂和雄成蜂均有丰富的体色变化(图3, 4)。基于各个组成部分的体毛颜色及胸部色块形状,短头熊蜂雌成蜂共有81种体色模式,其中来自122个采样点的795头工蜂有80种体色模式,来自10个采样点的103头蜂王有13种体色模式,工蜂和蜂王重叠的体色模式为12种(w1&q1, w2&q2, w3&q3, w4&q4, w5&q5, w7&q6, w8&q7, w10&q8, w11&q9, w13&q10, w19&q11, w28&q12)(图3)。来自12个采样点的59头雄成蜂有37种体色模式,除少数样本的翅基部体毛颜色为白色(m10, m14, m16)(图4)外,其中28种体色模式与雌成蜂相似,在这12个采样点雌成蜂的体色模式较多,但均有与雄成蜂相类似的样本,表明短头熊蜂的体色不存在性二型。

图3 云南短头熊蜂雌成蜂背面体毛的体色模式

图4 云南短头熊蜂雄成蜂背面体毛的体色模式
雌成蜂的各体色模式的分布频率是极不均衡的,依据81种体色模式的Berger-Parker指数(图5),优势体色模式有2种(w1&q1,D=0.1169; w2&q2,D=0.1069),胸部中间为长方形黑色体毛、边缘体毛为橙黄色,腹部第4-6节背板体毛分别为黑色和深棕色;丰盛体色模式有5种(D: 0.0523~0.0969),为w3&q3, w4&q4, w5&q5, w6, w7&q6;常见体色模式有10种(D: 0.0111 ~0.0479),为w8&q7, w9, w10&q8, w11&q9,w12, w13&q10, w14, w15, w16和w17;其余均为稀少体色模式类型(D: 0.0011~0.0089),共计64种(图5)。

图5 云南短头熊蜂雌成蜂体色模式的贝格-派克指数
对短头熊蜂雌成蜂体色模式进行聚类分析,结果显示: 81种体色模式可聚为两大体色模式类群,即A类群及B类群(图6),这两类之间的最大区别在于胸部背板的体毛颜色不同,A类群(包含33种体色模式)胸部背板的体毛全为黑色或者胸部背板中间大部分为黑色毛、圆形色块,四周全部或部分为灰黄色,B类群(包含48种体色模式)胸部背板体毛全部为橙黄色或者中央为黑色、四周为橙黄色。

图6 云南短头熊蜂雌成蜂体色模式聚类
A类群腹部第2节背板全部为黄色或上部大部分为黄色、下边缘两侧为黑色毛,第3节背板体毛为黄色、黑色,两种颜色所占的比例变化较大,在全黄和全黑之间逐渐过渡,第4节背板全部为黑色或深棕色或上半节为黑色、下半节为深棕色,第5节背板全部为深棕色或上半节为黑色、下半节为深棕色,第6节背板全部为深棕色(图6)。根据腹部背板体毛的颜色特征,B类群又可分为两个小类群:B-1类群(包含6个体色模式)腹部第2背板体毛全部为黄色、黑色或灰黄色,或者为上部黄色、下部黑色,两种颜色所占的比例变化较大,第3-6节背板全为黑色;B-2类群(包含42种体色模式)腹部第2节背板与B-1类群相同,第3背板以全黑色为主,部分第2背板全黄色体色模式中第3背板中间部分或上半节中间部分被黄色毛,个别体色模式中第3背板上半节为黑色、下半节为深棕色,第4节背板体毛全部为黑色或上半节为黑色、下半节为深棕色,第5节背板体毛全部为黑色或深棕色或上半节为黑色、下半节为深棕色,第6节背板体毛全部为深棕色(图6)。综上所述,短头熊蜂体色模式多态性丰富,体色模式可聚类划分为典型的3个体色模式类群,即:A类群(黑胸棕尾),B-1类群(橙胸黑尾),B-2类群(橙胸棕尾)。
2.2 云南短头熊蜂体色模式类群的地理区划
对经纬度0.5°×0.5°单元格内3个典型体色模式类群(A, B-1和B-2)的“有”/“无”分布进行系统聚类,结果显示:在距离22.5可聚为两大类,距离17.5处进一步聚为4小类,其中贵阳的样本与云南昭通市北部的样品共同聚为Ⅱ2(图7)。
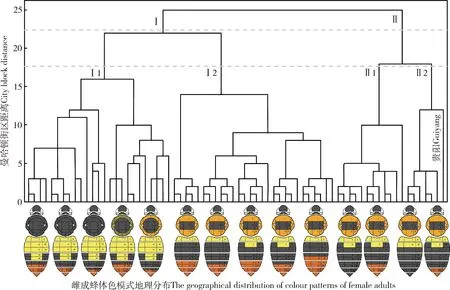
图7 基于经纬度0.5°×0.5°单元格内云南短头熊蜂雌成蜂体色模式类群地理分布的系统聚类分析
依据聚类结果和自然地理区划的区域共轭性原则,短头熊蜂体色模式在云南分布点的自然地理区划可分为云南高原西部(Ⅰ)和云南高原东部(Ⅱ)两个大区,Ⅰ区进一步可分为滇中亚热带高原区(Ⅰ1)和滇西南热带低山宽谷区(Ⅰ2),Ⅱ区进一步可分为滇东南亚热带岩溶山地河谷区(Ⅱ1)和滇东北亚热带中山河谷区(Ⅱ2)(图8)。短头熊蜂体色模式的分布及相应地理区划的自然景观特性,见表1。
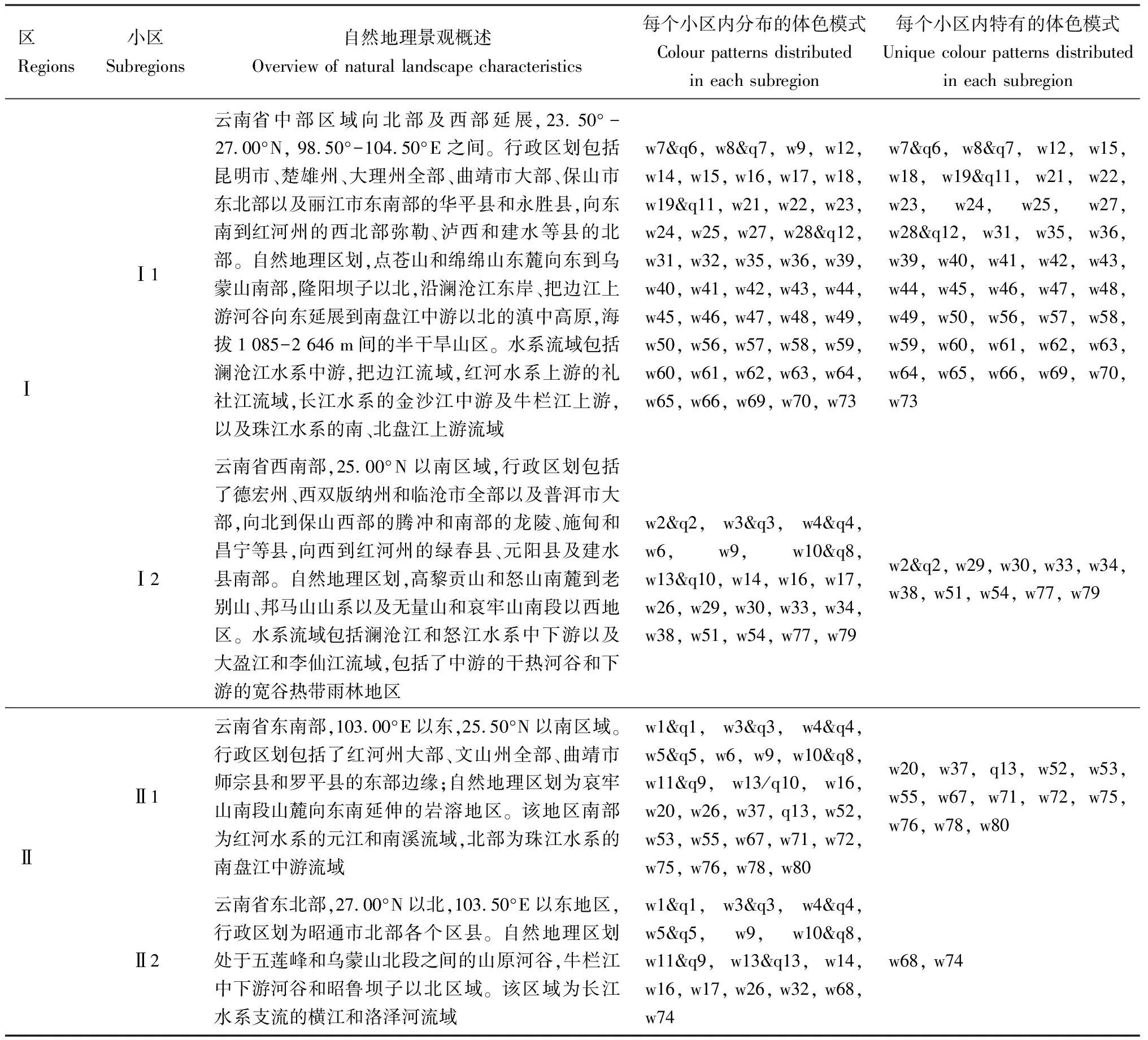
表1 云南短头熊蜂雌成蜂体色模式的分布及地理区划的自然景观特性

图8 云南短头熊蜂雌成蜂体色模式类群的地理区划[审图号: 云S(2023)129号]
雌成蜂的81种体色模式中,在Ⅰ1小区分布的有47种,其中特有体色模式42种,5种体色模式(w9, w14, w16, w17和w31)与其他小区共有,在Ⅰ2小区分布的体色模式有20种,在Ⅱ1小区分布的有24种,在Ⅱ2小区分布的有15种,4个小区均出现的体色模式为w9和w16(表1)。Ⅰ区分布的短头熊蜂腹部第4, 5, 6背板全节或部分体毛为深棕色,其中分布在Ⅰ1小区的大多数短头熊蜂为A体色模式类群,即胸部背板体毛全为黑色或中间为圆形色块黑色毛、四周全部或部分掺杂灰黄色,Ⅰ1, Ⅰ2和Ⅱ1小区交汇区域,建水-曲江坝子北部即Ⅰ1小区南部的短头熊蜂存在胸部中间体毛黑色,色块形状由小渐变大趋于圆形,四周橙黄色,腹部第1-3节背板均有黄色体毛,第4-6节体毛有深棕色的体色模式(w25, w46, w47, w48, w49, w69和w70)(图3),虽然聚到B-2体色模式类群,但是腹部颜色与A类群更相似;Ⅰ2小区仅分布有B-2体色模式类群,主要特征为胸部背板中间被长方形色块黑色毛、四周体毛橙黄色,腹部第4节背板全部深棕色或上半节为黑色、下半节为深棕色,第5和6节背板体毛全为深棕色,该区域内体色模式变异较小,腹部深棕色覆盖面积最多,第3背板下半节及第4-6节背板全棕色的体色模式仅在该区域发现。Ⅱ区分布的短头熊蜂胸部背板中间为方形或圆形色块,体毛黑色、四周为橙黄色或整个背板覆盖橙黄色毛,腹部第6节背板逐节向前延展至第4节,至少1节或多节的全节或部分体毛为深棕色或黑色,且两种颜色覆盖的范围变化较大,即Ⅱ1和Ⅱ2小区同时分布有B-1和B-2体色模式类群,但胸部背板体毛全为橙黄色的体色模式(w53, w72, w73, w75和w78)(图3)仅在Ⅱ1小区内发现。基于短头熊蜂体色模式类群的地理区划(图8),偏相关分析显示:体色模式多态性丰富度与其分布地的经纬度相关性不显著(P>0.05),云南高原中部(24.00°-24.50°N, 102.50°-103.00°E)(江川、华宁、通海等县及建水县的北部)的短头熊蜂的体色模式多态性最为丰富,包含18种体色模式,胸部全黑色、四周灰黄色和四周橙黄色类群均在该地出现。
综上所述,短头熊蜂体色模式分布有明显的地理区域性,不同地区的体色模式类群不同,且具有向区域内优势体色模式趋同的特性。
2.3 短头熊蜂体色模式类群的垂直分布
短头熊蜂采集地的海拔高度在346-2 646 m之间。基于3个典型的体色模式类群A, B-1和B-2的采样点海拔高度进行单因素方差分析,结果显示:A类群[(1 951±311) m]和B-1[(1 568±352) m],B-2[(1 556±534)m]类群分布地的海拔高度差异极显著(P<0.01),B-1和B-2类群之间海拔高度差异不显著(P>0.05),A类群分布地的海拔高于B类群,B类群分布地海拔高度变化范围较大(346~2 482 m)(图9)。
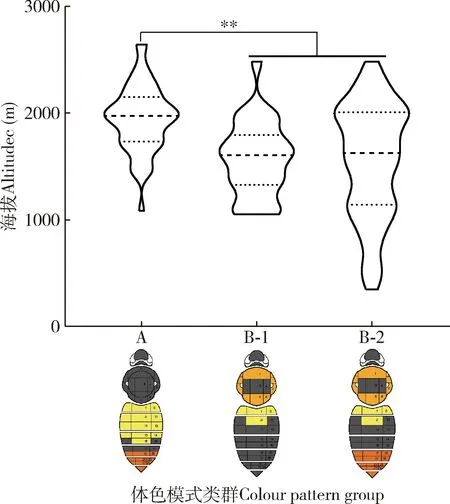
图9 云南短头熊蜂雌成蜂体色模式类群的垂直分布
3 讨论
短头熊蜂作为体色多态性与缪氏拟态研究的重要材料,分布于喜马拉雅山西南麓及中国长江流域以南的东南亚地区。虽然本研究的样品遍布云南高原和横断山脉南段,但由于云南熊蜂地理分布受到地形地貌和气候的影响,区系特征明显(梁铖等, 2018),所以本研究所采集的样品可能不能完全代表自然界中短头熊蜂体色模式多态性的全况,可能还存在未发现的体色模式。短头熊蜂的年活动周期较长(Williamsetal., 2009),在云南东南部,蜂王在3月下旬出蛰,12月上旬三型蜂才会消亡(Liangetal., 2020),而本研究中大多数样品的采集集中在6-8月期间,因此收集到的雄成蜂和蜂王的样品数量相对较少,再者其体色模式多态性在级型方面不存在性二型,因此本研究主要对标本数量较多的雌成蜂的体色模式多态性及其地理分布格局进行分析。已有研究报道,短头熊蜂的雌成蜂有13种体色模式(Hines and Williams, 2012),而本研究参照Williams(2007)熊蜂体色模式的绘制方法(图2),忽略翅基部体毛颜色的渐变(Williamsetal., 2009),发现云南短头熊蜂有81种体色模式,其中包含了文献报道的11种(Hines and Williams, 2012; Sinpooetal., 2019),具有非常丰富的体色模式多态性。
熊蜂以不同的体色模式适应不同的生境,缪氏拟态、体温调节、保护色等因素都可能对熊蜂的体色起到选择作用(Williams, 2007)。参照上述研究结果,云南短头熊蜂雌成蜂的81种体色模式均属于有着鲜明颜色条带的类群,可能作为对捕食者的集体警告信号。短头熊蜂与三条熊蜂和红尾熊蜂呈现出一系列相似的体色模式(Hines and Williams, 2012),是典型的缪氏拟态。而巨熊蜂亚属的山岭熊蜂B.montivagus与短头熊蜂在云南的同域分布范围更广,体色模式也趋同(黄家兴, 2015; 梁铖等, 2018),也可能是缪氏拟态的结果。物种内丰富的体色模式以及同域范围内不同蜂种之间体色模式的趋同影响了熊蜂物种形态鉴定的准确度。在没有模式标本的情况下,种内具有新体色模式的个体很容易被误认为是新物种,而具有相同体色模式的不同蜂种又被误判为同一物种,形成隐存种(Williamsetal., 2012; Huangetal., 2015),所以需要结合地理分布、面部和腹部轮廓特征、上颚“牙齿”、雄成蜂生殖器和翅形态等多个指标才能对熊蜂的种类进行准确鉴定。虽然不能单独依赖体色模式对熊蜂种类进行鉴定,但详实的体色模式图谱,对总结蜂种的特征及蜂种间的差异仍大有裨益。
东洋区物种双色熊蜂B.bicoloratus主要分布在中国南部的中、低海拔地区,体色变化广泛,来自海南、台湾的样本(胸部体毛为全黑色、腹部体毛全部为深棕色)与大陆样本的体色模式差异较大,这种差异主要与海岛隔离和气候变量(太阳辐射强度)相关(Dingetal., 2019)。短头熊蜂的地理分布与双色熊蜂相似,本研究发现的短头熊蜂的81种体色模式并不包括之前报道的台湾(胸部体毛为全黑色,腹部第1-3节背板全黑色,第4-6节背板深棕色)和泰国中部(胸部体毛为全黑色,腹部第1节背板黄色,第2-3节背板黑色,第4-6节背板深棕色)的两种体色模式(Hines and Williams, 2012)。台湾与大陆短头熊蜂的体色差异在于,台湾样品腹部第1-3节背板的体毛呈全黑色,这种差异也可能是由于海岛的隔离效应所致,但是否与气候变量有关还需进一步研究。泰国中部与云南的短头熊蜂有细微的体色差异,这种差异的形成原因尚需进一步调查研究。
云南短头熊蜂体色模式地理区划特征明显,可分为2大区、4个小区,与云南高原的气候区划相似,Ⅰ1小区主要为中亚热带气候,Ⅰ2小区的河谷、盆地地区为北热带气候,Ⅱ1小区主要为中亚热带气候,Ⅱ2小区的河谷地区为北亚热带气候(段旭等, 2011; 段旭和陶云, 2012)。Ⅱ2小区与其他小区不相连,26.50°-27.50°N间没有采集到短头熊蜂样本(图1),这种格局的出现,除采样受到行政区划的限制外,更可能的原因是短头熊蜂不适应间隔区域的地形和气候。该区域南部属于金沙江水系干热河谷和乌蒙山南段高海拔山地,可能阻碍了短头熊蜂向北扩散,接着向北为牛栏江中下游河谷和昭鲁坝子,整体为温带气候带(王声跃和张文, 2002),加之该区域土地利用以耕地和城市为主(李东梅等, 2010),短头熊蜂可能不适应该区域的生境。本研究还发现A体色模式类群仅分布于Ⅰ1小区,而Ⅰ2小区仅分布有B-2体色模式类群,Ⅱ1小区和Ⅱ2小区内有B-1和B-2体色模式类群同时分布,但Ⅱ2小区内B-2体色模式类群比Ⅰ2小区内的B-2体色模式类群腹部深棕色与黑色比例变化更大,这一分布格局的形成可能与气候相关。Ⅰ1小区的年均温和年降雨量均低于Ⅰ2小区和Ⅱ1小区,但平均海拔较高,日温差较大(陈宗瑜, 2001),分布在Ⅰ1小区的A体色模式类群胸部更多的黑色体毛可能利于短头熊蜂在光照下快速吸收热量、升高体温,来应对较高海拔分布区的更大的日温差变化。昆明地区(Ⅰ1)A体色模式类群与屏边、个旧地区(Ⅱ1)B体色模式类群的蜂王的繁育特性存在差异(秦加敏等, 2022),也佐证了云南短头熊蜂可能存在不同的地理种群。结合云南地形地貌特征可发现,Ⅰ1小区西部由怒江河谷,西南部由把边江和李仙江上游的河谷、盆地,南部由绿汁江河谷分别与Ⅰ2小区分隔,Ⅰ1小区东段的南部由南盘江河谷及弥勒-泸西山间谷地,东部由南盘江的南北向的支流河谷以及南北走向的罗平坝子和块泽河河谷分别与Ⅱ1小区分隔,Ⅰ2小区东南部由元江及支流河谷与Ⅱ1小区分隔;另外3个小区在建水-曲江坝子交汇(王声跃和张文, 2002)。Ⅰ1小区, Ⅰ2小区和Ⅱ1小区间的河谷和盆地多呈现干热气候,高温干燥(闵石头和王随继, 2008; 欧朝蓉等, 2015),山间盆地(坝子)多为农业种植区和城市发展区域(李东梅等, 2010),均不利于熊蜂生存(梁铖等, 2018)。综上所述,云南短头熊蜂主要分布在亚热带和热带地区,胸部颜色更黑的A体色模式类群对于高海拔地区的气候适应性更强,体色模式的有序分布可能是缪氏拟态和体温调节共同驱动,体色模式类群地理分布受到气候区划、地形地貌和土地利用类型的影响,地理区划特征明显。
Ⅰ1, Ⅰ2和Ⅱ1小区间交汇区域的少数采样点,短头熊蜂样品胸部的体毛存在多种颜色,如分布在建水北部、通海以及宣威的样品,其胸部体毛呈现出全黑和橙黄色条带间渐变,有中间类型灰黄色,而来自相同采样点的短头熊蜂的腹部黄色体毛的覆盖面积较固定。这种胸、腹部体色所表现出的不同分布特征可能与地理分布的环境选择和遗传调控有关,胸、腹部体色的调控机制很可能存在差异。典型的Ⅰ1南部即建水北部采样点存在胸部与B-2体色模式类群相似,而腹部颜色与A体色模式类群更相似的样本,可能是不同体色模式的短头熊蜂在该区域基因交流的结果,但这些体色模式未向四周扩散,推测环境压力对体色模式适应性选择,造成了遗传过程中部分不适应区域生境的基因型丢失,也可能是3个体色模式类群地理区划明显的成因。中国北方的朝鲜熊蜂B.koreanus具有9种体色模式,工蜂胸部颜色由棕色到黑色,中间还存在多个过渡颜色类型(Huangetal., 2015),云南短头熊蜂的体色也存在类似的变化,胸部体毛颜色有全橙黄色,全黑色,胸部背板中间为方形色块黑色毛、四周为橙黄色,或圆形色块黑色毛、四周为灰黄色等多种变化。根据人工饲养的B.melanopygus后代蜂群的体色模式特征,推测出B.melanopygus的腹部第2和3节背板体毛的黑色和深棕色是由单一位点的两个等位基因调控的,深棕色相对于黑色是显性(Owen and Plowright, 1980)。全基因组关联分析,进一步探明了位于腹部体色决定基因abd-A和Abd-B之间的顺式调控位点控制着B.melanopygus腹部深棕色-黑体色之间的转换(Tianetal., 2019)。Ⅱ区大多数采样点同时分布有B-1(橙胸黑尾,第4-6节背板体毛为黑色)和B-2(橙胸棕尾,第4-6节背板含深棕色体毛)两个类群,推测短头熊蜂可能存在与B.melanopygus类似的调控机制。但是,短头熊蜂腹部背板的黄色体毛由第1节背板向后逐节延伸,最多至第3节背板整节,深棕色体毛由第6节背板向前逐节延伸最多至第3节背板的下半节(图10),这种有序、 连续变化的调控机制不明。目前, 本研究团队已经实现了短头熊蜂的人工周年繁育(Liangetal., 2020), 为进一步分析其体色调控机理奠定了基础。

图10 云南短头熊蜂雌成蜂不同体节体色的渐变
本研究结合文献报道可发现:A体色模式类群(黑胸棕尾)分布于24.00°N以北、104.50°E以西地区,由Ⅰ1小区滇中亚热带高原向北延伸到四川境内普格县螺髻山,向西延展到西藏南部和喜马拉雅山脉西南山麓区域;而在24.50°N以南、103.00°E以西的横断山脉南段Ⅰ2小区滇西南热带低山宽谷区向南延伸到中南半岛区域,短头熊蜂的体色模式大多属于B-2体色模式类群(橙胸棕尾),且腹部第4-6节背板均含有深棕色体毛;B-1体色模式类群(橙色黑尾)仅分布在103.00°E以东的滇东南岩溶山地河谷(Ⅱ1),滇东北中山河谷(Ⅱ2)、贵州及四川盆地东部向东延伸到华南地区的浙江省境内以及越南的北部区域,该区域内同时分布有B-2体色模式类群,且腹部第4-6节背板由后向前逐节延伸,仅有1节或多节体毛深棕色,个体间较多变(何俊华和王淑芳, 2004; Williamsetal., 2009; Hines and Williams, 2012)。可以看出,短头熊蜂主要的3个体色模式类群在云南境内有明显的生物地理分区边界。
综上所述,短头熊蜂体色模式多态性丰富,目前全球发现83种雌成蜂体色模式,其中云南境内分布有81种,这些体色模式可聚为3个体色类群,其地理分布受到气候、地型地貌和土地利用类型影响,地理区划特征明显。以云南为中心,Ⅰ1小区向西延展到南亚北部地区,Ⅰ2小区向东南亚的西南部延展,Ⅱ区延展到东南亚东部地区,构成短头熊蜂不同体色类群分布的3个地理区划格局。
致谢云南省农业科学院蚕桑蜜蜂研究所的杨爽、周春涛、赵洪木以及贵州省农业科学院现代农业发展研究所的贺兴江参加了部分熊蜂样品的野外采集工作,云南省农业科学院蚕桑蜜蜂研究所的赵必安参加了标本整理和绘图工作,在采样过程中云南省各自然保护区和森林公园管理局(处)给予了大力支持和帮助,在此一并表示感谢。
