绿色电子供给和细胞代谢调控型反硝化研究
2024-02-22范心韵张学萌朱梦茹陈银广
刘 超, 范心韵, 张学萌, 陈 闯, 朱梦茹, 陈银广
(同济大学 环境科学与工程学院 污染控制与资源化研究国家重点实验室, 上海 200092)
0 引 言
氮素循环是生物圈内基本的物质循环过程之一,微生物在不同价态的含氮物质相互转化过程中发挥重要作用。由于含氮产品的大量生产和使用,水环境中的氮污染问题日趋严重,主要包括市政生活污水、工业含氮废水以及氮肥施用过程中的地表水污染。作为全球氮循环的关键环节,微生物反硝化是去除水环境中硝酸盐污染、实现氮元素返回大气和维持氮平衡的重要途径[1-2]。反硝化微生物通常以有机或无机物质为电子供体,硝酸盐为电子受体,使硝酸盐逐步还原为亚硝酸盐、一氧化氮、氧化亚氮,最后形成氮气。其中,依赖于有机碳源供给的异养反硝化是目前主流的生物脱氮工艺,对于具有低碳氮比特性的市政污水等,通常需要额外补加昂贵的乙酸、葡萄糖等商品碳源以获得较好的反硝化效果,这导致了运行成本提高和碳排放量增加。在“双碳”目标和循环经济可持续发展的背景下,开发商品碳源投加减量或无机自养低碳反硝化的脱氮新技术成为学界和产业界的共识。
已有文献主要综述了以S、Fe、H2等无机物作为电子供体的硫基自养[3-4]、铁基自养[5-6]和氢基自养低碳反硝化技术[7-8]。近年来,科研界不仅致力于开发新型反硝化无机绿色电子供体如光生电子[9-12],还对短链气态烷烃[13-16]、有机废弃物[17-20]等绿色有机碳源开展了大量研究。同时,也有国内外学者在不增加有机碳源的前提下,通过提升电子供体的利用效率获得高效的反硝化性能,达到节约碳源的生物脱氮目的[21-24]。本文首先系统梳理了光电反硝化过程中光敏剂的种类、主导功能微生物、光敏剂-微生物复合方式以及光电营养反硝化的应用现状;然后归纳总结了近年来生物反硝化过程中使用的短链气态烷烃等绿色有机碳源;最后从促进电子传递和增强反硝化酶活性两方面,探讨分析了基于细胞代谢调控提升电子供体利用效率来促进反硝化的作用原理,以期拓展对高效脱氮机制的理解,并展望反硝化生物脱氮技术的未来发展方向。
1 光生电子驱动厌氧反硝化
光生电子驱动的厌氧反硝化是一种新兴的硝酸盐还原过程,主要基于光敏剂-脱氮微生物杂化复合体。清洁的光能激发光敏剂产生光电子,供给不能利用光能的脱氮微生物进行反硝化代谢,又称之为光电营养反硝化。光电营养反硝化效应因其在太阳能利用方面的优异表现和对生物地球化学过程的新认识,且光生电子兼具低碳绿色环保和成本低廉等优势,受到研究人员和环境工程师广泛关注。
1.1 光敏剂种类
在光敏剂-微生物杂化体系中,光敏剂的类别决定了光生电子的供给能力。目前,应用于光电反硝化体系的光敏剂主要包括固相的TiO2[25]、CdS[9]、赤铁矿[26]、C3N4[27]和溶解性有机质(DOM)等[11]。例如,CHEN等首次构建了TiO2光阳极与微生物阴极体系,阳极在30 mW/cm2的紫外光照射下产生的电子传递给阴极的脱氮微生物[25]。该反应体系硝酸盐还原率超过了95%,且该效率与乙酸作碳源、碳氮比为9的异养反硝化脱氮效率相当。研究人员又通过硫化镉-脱氮硫杆菌Thiobacillusdenitrificans杂化复合体(T.denitrificans-CdS)高效还原硝酸盐[28],但该体系受反应过程中产生的超氧化物自由基等活性氧物质和游离亚硝酸影响,反硝化副产物氧化亚氮N2O含量较高。随后,CHEN等利用Mn3O4纳米酶修饰T.denitrificans-CdS,构建了T.denitrificans-CdS@Mn3O4复合体系,发现Mn3O4可以有效淬灭活性氧,使得硝酸盐还原率显著提升和N2O排放量大幅降低[10]。与添加人工合成的光敏剂不同,生物自合成材料具有生物相容性好等特性。反硝化微生物铜绿假单胞菌Pseudomonasaeruginosa通过在胞内合成硒化物/硫化物量子点[12],自组装形成光敏剂-生物复合体,实现了低碳氮比废水中硝酸盐的有效去除。
考虑到光照的持续供给问题,研究人员开发了一种新型光敏剂,氰酰胺功能化的庚嗪环基聚合物(NCN-CNx)[27]。与传统的光电反硝化体系相比,该体系在黑暗环境下表现出优异的硝酸盐去除性能,因为深色光催化剂(NCN-CNx)可以储存光生电子,并根据需要在无光照条件下通过石墨烯助催化剂释放电子供反硝化微生物使用。进一步分析表明,NCN-CNx主要增强了反硝化微生物的硝酸盐还原酶、亚硝酸盐还原酶和过氧化氢酶活性,并提高了电子传递速率。这些发现为延时光催化剂促进反硝化的机理和建立硝酸盐昼夜连续去除的光催化系统提供了新的见解。
DOM的生物地球化学归趋在水生生态系统中具有重要的环境意义。DOM与微生物互作关系的认识通常仅限于DOM作为电子穿梭体或异养菌的碳源。有趣的是,研究人员最近证明了DOM在阳光照射的水生态系统中作为光敏剂,驱动模式反硝化微生物脱氮硫杆菌进行脱氮代谢,且微生物反硝化基因(包括Nar、Nir、Nor和Nos)的表达显著上调[11]。这不仅提供了一种促进地表水环境中光驱反硝化的策略,而且由于DOM成本低且应用广泛,DOM主导的光电反硝化技术极具前景。
1.2 功能微生物
脱氮微生物为光电反硝化过程提供关键的硝酸盐还原反应模块。微生物纯培养技术有利于确定实验因素和研究脱氮代谢途径。目前,应用于光电反硝化过程的主要纯菌为脱氮硫杆菌Thiobacillusdenitrificans和铜绿假单胞菌Pseudomonasaeruginosa。Thiobacillusdenitrificans是一种广泛分布的严格化能自养生物,具有“硝酸盐→亚硝酸盐→一氧化氮→氧化亚氮→氮气”的完整氮素还原途径,是研究反硝化过程的模式生物[29]。起初,它被发现能够接受外部固体电极的电子进行反硝化[30-32]。近年来,众多研究证明尽管T.denitrificans无法直接利用光能,却可以吸收光激发光敏剂产生的电子进行生长代谢并实现硝酸盐的去除[10-11, 28]。Pseudomonasaeruginosa也具有完整的硝酸盐还原为氮气的生化代谢途径,被证实可以有效利用光生电子进行厌氧反硝化反应[12, 26]。与T.denitrificans不同,P.aeruginosa可以进行异养生长,这表明能够接收外部光电子的微生物具备作为光电反硝化体系构建模块的潜力。后续研究可以深入解析不同微生物对光生电子的吸收和利用路径,如调控该途径的关键运输蛋白和/或介质,寻找更优的电子传递路径,并可借助基因工程等手段调控电子代谢过程,促进电子的高效吸收与利用。
混合微生物体系对培养环境条件要求相对于纯菌更低,稳定性高,更接近实际自然环境,被广泛应用于光电营养反硝化。ZHANG等通过在负载了C3N4/TiO2复合材料的海绵上原位培养生物膜,成功构建了光敏剂-生物膜杂化体系[33]。微生物群落分析表明,酸杆菌门Actinobacteria和绿弯菌门Chloroflexi得以大量富集。以往研究发现,Actinobacteria和Chloroflexi对于氮循环过程中硝酸盐和亚硝酸盐的还原具有重要作用[34-35]。与易受光催化影响的单一细菌物种相比,生物膜在该耦合系统中更能抵抗不利条件,表现出良好的适应性。ZHU等则发现在光电营养反硝化体系的构建过程中,反硝化微生物鞘氨醇单胞菌科Sphingomonadaceae、黄单胞菌科Xanthomonadaceae、几丁质菌科Chitinophagaceae和环杆菌科Cyclobacteriaceae的丰度大幅提升[36]。ZHOU等的研究也发现了类似的微生物富集现象[27]。
1.3 光敏剂-微生物复合方式与光电反硝化的应用
光敏剂与微生物的结合位置决定了微生物吸收光生电子的行为和路径。如图1(a)所示,最初的研究是将光敏剂制作成阳极,光照激发阳极半导体材料产生电子,经外部导线传输至阴极电极,阴极室的微生物从电极上摄取电子进行反硝化反应[25]。这种构建方式可以将光生电子产生模块与微生物催化单元有效分开,避免了光照对微生物细胞的潜在不利影响,但是该过程涉及“光生电子→电极→微生物”多级电子传输步骤。为了缩短电子的传输步骤和距离,光敏剂被负载在反硝化微生物的细胞表面,光敏剂中产生的光生电子直接穿过细胞膜并输送到细胞内以驱动或增强生物反硝化代谢过程[10, 28, 37](图1(b))。进一步研究发现电子跨膜转移需要克服细胞膜屏障,传输动力学较为缓慢[38-39],限制了光电反硝化反应速率。图1(c)展示了一种巧妙的杂化体系构建方式[12],通过在微生物胞内合成很小尺寸的量子点,使得光生电子直接在胞内产生,从而不仅明显缩减电子至胞内代谢反应位点的距离还规避了电子的跨膜传质障碍。
近年来,从最初的实验室硝酸盐配水实验,光电反硝化研究逐步面向实际废水和地表水等实际场景。例如,ZHU等在地表水中原位构建了光敏剂TiO2-周丛生物膜系统,其中,TiO2不但优化了周丛生物膜中反硝化微生物的组成,为微生物提供光生电子,而且通过刺激细胞外聚合物的分泌(尤其是腐殖酸和蛋白质)促进光生电子传递,从而有效去除了地表水中硝酸盐,为光电营养反硝化在实际水体环境中的应用提供了坚实基础[36]。又有研究发现,在不添加外源电子供体的条件下,Pseudomonasaeruginosa-CdSexS1-x生物杂化体系能够以光激发产生的电子为能源进行脱氮生物代谢反应,从而有效去除城市污水、养殖废水和电镀废水中的硝酸盐[12],有力验证了光电营养反硝化在实际水处理场景中的适用性。

图1 光电营养反硝化过程中光生电子传递途径Fig. 1 Photogenic electron transfer pathway in the process of photoelectric nutrition denitrification
2 绿色有机碳源介导生物反硝化
2.1 短链气态烷烃
天然气中的短链气态有机烷烃主要包括甲烷、乙烷、丙烷等。近年来,以气态烷烃作为电子供体,硝酸盐和亚硝酸盐作为电子受体的厌氧反硝化相关研究引起人们的广泛关注。其中,甲烷作为天然气中占比最高(60%~80%)的烷烃,其主导的反硝化过程被称为反硝化型厌氧甲烷氧化(Denitrifying Anaerobic Oxidation of Methane,DAOM),因兼具温室气体甲烷控制和氮素去除的双重功能备受推崇。DAOM一般发生在湖泊、河流底泥和湿地土壤等自然环境以及污水处理厂中,由DAOM古菌(模式微生物为ANME-2d中的CandidatusMethanoperedensnitroreducens)和DAOM细菌(模式微生物为NC10门中的CandidatusMethylomirabilisoxyfera)进行[14, 40]。近期研究者逐渐关注丙烷、丁烷等其他短链气态烷烃作为反硝化电子供体的可能性。例如,GUO等通过富集培养、同位素标记实验和多维宏组学方法,首次发现了一种新型的丙烷厌氧氧化耦合反硝化过程并揭示其细胞代谢机制。该细菌属于Firmicutes门Symbiobacteriia纲谱系,被命名为CandidatusAlkanivoransnitroreducens[15]。
如图2所示,DAOM古菌、DAOM细菌和丙烷厌氧反硝化菌的代谢途径存在较大的差异。在硝酸盐还原过程中,由于DAOM古菌仅表达硝酸盐还 原酶,只能将硝酸盐还原为亚硝酸盐[40]。DAOM细菌虽然缺少硝酸盐还原酶,但能够表达将亚硝酸盐还原为氮气的一系列反硝化酶[41]。与DAOM细菌相比,DAOM古菌还能够表达产氨亚硝酸盐还原酶,将亚硝酸盐还原为铵进而利用[14]。在甲烷氧化过程中,DAOM古菌通过反向产甲烷途径将甲烷转化为二氧化碳,而DAOM细菌通过甲烷单加氧酶利用反硝化过程产生的氧气来氧化甲烷[41]。与DAOM古菌和DAOM细菌不同,Ca.A.nitroreducens能够在氧化丙烷的同时进行完整的反硝化过程,并能够将亚硝酸盐还原为铵,同时还表达了一氧化氮→氧化亚氮→氮气的代谢通路[15]。研究结果表明,Ca.A.nitratireducens参与厌氧丙烷氧化的关键基因主要是Akylsuccinate synthase alpha subunit (AssAD),其在富马酸介导下完成丙烷活化第一步,生成琥珀酸丙酯/异琥珀酸丙酯。Ca.A.nitratireducens含有进一步降解琥珀酸丙酯/异琥珀酸丙酯所需的所有基因,包括用于碳骨架重排的甲基丙二酰辅酶a变位酶(McmA)基因、用于脱羧的丙酰辅酶a羧化酶(PccB)基因和β氧化的关键基因等(图2(c))[15]。
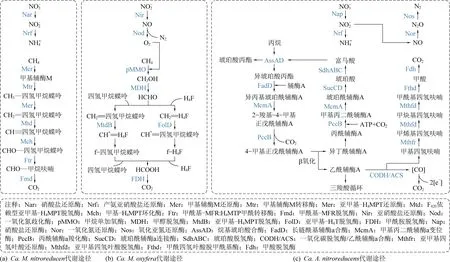
图2 DAOM古菌Ca. M. nitroreducens、DAOM细菌Ca. M. oxyfera和丙烷厌氧反硝化菌Ca. A. nitroreducens的代谢途径Fig. 2 Metabolic pathways of nitrate reduction in DAOM archaea Ca. M. nitroreducens, DAOM bacteria Ca. M. oxyfera and propane anaerobic denitrification bacteria Ca. A. nitroreducens
污水处理厂厌氧段产生的甲烷一部分逸散入大气,另一部分仍溶解在污水中(10~30 mg/L)[13]。通过在反硝化段驯化引入DAOM古菌和细菌,能够利用污水中溶解的甲烷作为电子供体进行反硝化脱氮,不仅可以减少乙酸等商品碳源的投入,还能够避免甲烷进入大气而加剧温室效应。已有较多研究在实验室规模下将DAOM古菌和细菌引入反硝化系统,并实现了长期高效的运行。经过驯化,同时存在DAOM微生物和厌氧氨氧化微生物的颗粒污泥,具备1.0 kg N/(m3·d)的脱氮速率[42]。将DAOM过程和厌氧氨氧化过程耦合的生物膜系统,能够实现85%的溶解性甲烷去除和超过99%的脱氮效果。质量平衡研究表明DAMO细菌和DAMO古细菌确实共同促进了甲烷和氮素的去除,这项技术可用于降低污水处理厂碳足迹和常规有机碳源的使用[13]。丙烷和丁烷同样在实验室规模下被用作反硝化的电子供体,研究表明微氧条件下丙烷和丁烷能够支持相关微生物进行反硝化,硝酸盐去除速率约为40~50 mg/(L·d)[16]。尽管烷烃营养型反硝化具有诸多优势,但稳定的烷烃供应、气液传质、关键功能微生物生长速率慢和富集周期长等方面仍亟待解决以实现规模化应用。
2.2 有机废弃物
不同碳源构成了异养反硝化生长与代谢的基础。一方面,反硝化微生物可利用的碳源多种多样,比如乙酸等挥发性脂肪酸(VFAs)、糖类、醇类,碳源的种类会影响反硝化微生物的脱氮性能[43-44],也决定了硝酸盐还原速率和其中间产物如亚硝酸盐与N2O的积累[45]。另一方面,有机废弃物富含多糖等高生物降解性物质,通过厌氧生物发酵作用产生的代谢产物VFAs、易降解糖类葡萄糖等,是异养反硝化过程的优质碳源[17, 20]。近年来,国内外大量研究使用市政污泥、餐厨垃圾发酵液为代表的液态碳源和木质纤维素类废弃物为代表的固体碳源作为反硝化外加碳源,以替代昂贵的甲醇、乙酸、葡萄糖等商品碳源,在降低反硝化的运营成本基础上,还能实现废弃物的减量化,充分响应低成本绿色脱氮水处理技术的发展趋势[18, 46-47]。
以有机废物发酵液作为反硝化脱氮的外加碳源不仅能够获得优异的脱氮效果,还能减少反应过程中氧化亚氮等的排放,因此备受研究人员青睐。例如,ZHU等发现使用剩余污泥碱性发酵液作为碳源,反硝化脱氮性能良好,且产生的N2O和NO的排放相比于商品碳源乙酸实验组分别减少了68.7%和50.0%[48]。又有学者综合比较了初沉污泥、浓缩的废活性污泥、餐厨垃圾、厨余垃圾、脂类废物和乳清粉6种有机废弃物厌氧发酵液作为反硝化脱氮的碳源潜力[18]。结果表明,以发酵液为碳源的反硝化脱氮性能均优于常规碳源投加组(甲醇和乙酸)。其中,乳清粉发酵液实验组的反硝化速率最高,达到了17.6 mg NOx-N/(g VSS·h),几乎是以甲醇为碳源组的4倍,且氮去除率高达99%,无亚硝酸盐积累。值得注意的是,发酵液中的铵和磷需要进行回收以防止氮磷负荷增加,同时过滤等相质分离工艺是处理发酵液获得反硝化液态碳源的重要步骤[17, 49]。
例如,富含纤维素的生物质例如稻草、秸秆、玉米芯、木屑等均可作为反硝化的固体缓释碳源,用于低成本处理含氮污水[18-19, 46]。ZHU等使用玉米粉作为反硝化菌PseudomonasstutzeriPCN-1的碳源,不但脱氮效果良好,而且与琥珀酸盐、葡萄糖、乙酸盐、柠檬酸盐等碳源相比,反硝化过程中温室气体氧化亚氮的排放降低了52.07%~99.81%[50]。又有研究证实碱预处理的玉米芯作为焦化废水反硝化脱氮的固体碳源时,废水中90%的硝酸盐能够被有效去除[51]。SUN等选取玉米芯、小麦秸秆、锯末和核桃壳4种农业废弃物作为发硝化的固体碳源,发现玉米芯和小麦秸秆实验组的脱氮性能与商品碳源乙酸钠组相当[18]。机理研究发现,反硝化性能最初依赖于固体碳源释放的溶解性有机物,而与纤维素和半纤维素分解相关的糖苷水解酶、碳水化合物酯酶表达上调和GAPDH等糖酵解功能基因的富集,是保证电子供体持续供应的内在驱动力。
2.3 微生物内碳源
除了常见的乙酸等胞外碳源,反硝化功能菌还可利用内碳源进行氮代谢。微生物的内碳源主要包括细胞内的聚羟基链烷酸酯、聚-β-羟基丁酸、聚-β-羟基戊酸、聚-β-羟基-2-甲基戊酸等,内碳源的使用能够有效减少外部碳源的投加,节省运营成本,是低碳脱氮水处理的重要途径。内碳源的形成过程通常包括厌氧-缺氧阶段。如图3所示,厌氧阶段反硝化聚磷菌和反硝化聚糖菌降解储存在细胞中的聚磷酸盐或糖原,获得吸收VFAs合成聚羟基链烷酸酯的能量;缺氧阶段,微生物分解细胞内碳源产生大量电子用于硝酸盐的还原,并合成聚磷酸盐或者糖原[52-55]。内源性反硝化适用于低碳氮比污水的深度脱氮[56]。FAN等研究发现,反硝化聚磷菌与反硝化聚糖菌在缺氧脱氮除磷过程中具有协同作用,其充分利用了内碳源实现低碳脱氮,有效解决了市政污水处理碳源不足的问题[57]。
根据文献报道,反硝化聚磷菌主要包括CandidatusAccumulibacter、Dechloromonas、Acinetobacter、Thauera等,反硝化聚糖菌有Competibacter、Defluviicoccus、Contendobacter、Sphingobium以及Amaricoccus等[58-60]。混合菌群中反硝化聚磷菌和聚糖菌的生存和内碳源合成能力主要受溶解氧、胞外物质等因素的影响。例如,低溶解氧浓度能够明显增强微生物细胞基质利用、能量合成和代谢调控相关功能基因的活性,有利于聚磷菌Dechloromonas的生长增殖和维持较高的脱氮除磷代谢效率[60]。此外,有研究发现胞外聚合物中的有机物如酪氨酸类物质,能够被微生物用于合成细胞内碳源聚羟基链烷酸酯[61]。有些内碳源反硝化菌还能够与同步硝化反硝化[58, 62]、厌氧氨氧化[63]、短程硝化[64]相关联。例如,WANG等发现反硝化聚磷菌Thauerasp.菌株以2.85、1.98和2.42 mg N/(L·h)的平均速率同时去除氨、亚硝酸盐和硝酸盐[58]。LU等采用内碳源反硝化耦合厌氧氨氧化工艺,在进水碳氮比为3.16时,使得出水总氮低至1.87 mg/L;反硝化聚糖菌在缺氧阶段使用细胞内碳源将硝酸盐还原并积累大量亚硝酸盐提供给厌氧氨氧化菌,实现深度脱氮[63]。

图3 反硝化聚磷菌和反硝化聚糖菌脱氮原理示意图Fig. 3 Diagram of nitrate removal by denitrifying polyphosphate accumulating bacteria and glycogen accumulating bacteria
3 基于细胞代谢调控的高效反硝化
反硝化的本质是微生物利用电子供体还原高价态硝态氮的过程。与前文所述优化电子供体的思路不同,学者们发现在不增加碳源的前提下,通过提升电子利用效率也能够促进反硝化,从而达到节约碳源的目的。其中,调控反硝化过程的电子传递和消耗备受关注。
3.1 促进电子传递

近年来,通过添加氧化还原介体,提高电子传递调控蛋白活性或表达水平来促进电子传递增强反硝化脱氮的研究备受关注。氧化还原介体能够可逆地氧化和还原,在多个氧化还原反应中充当电子载体,并通过降低总反应活化能来加速反应[24, 66]。一些外源的氧化还原介体,诸如腐殖酸[67]、醌类物质[23, 68]、Fe-Pd/多壁碳纳米管[69]等,已被广泛报道用于促进微生物反硝化脱氮。氧化还原介体大多含有醌、酚等氧化还原基团,在反硝化过程中可能具有类似于泛醇/泛醌的功能,在电子传输中起重要作用[70]。例如,生物炭中含有大量具有氧化还原特性的醌类化合物,一些官能团还可以作为反硝化细菌的直接电子供体,直接或间接促进种间电子转移。SATHISHKUMAR等探究了不同温度下制备的生物炭对反硝化的影响,结果表明,400 ℃制备的生物炭含有较多的氧化还原活性物质,刺激C型细胞色素等导电物质的生成,提高了电子传递效率从而促进反硝化过程[21]。又如,JIA等通过在微生物细胞中原位合成钯纳米颗粒[22],发现其能够显著提升Fe-S中心活性并加速电子传递,从而促进反硝化脱氮。外源投加半胱氨酸也被证实提高了反硝化细菌胞内电子传递相关蛋白的表达,增强了反硝化效果[71]。尽管外源氧化还原供体可以加速反硝化过程,但是由于其价格高昂且有副产物产生,不适合大规模工程应用。因此,近年来一些研究者聚焦于微生物分泌的内源性氧化还原介质,如黄素类、吩嗪类和细胞色素类等,并探究了其对反硝化过程的影响[72-73]。WANG等发现微生物分泌的胞外聚合物在电子传递链中起到类似于醌环的作用,可以作为电子载体,加速电子传输,使得硝酸盐还原率增加了1.42倍[24]。可见,通过内源产生的氧化还原介质提高电子传递效率能够强化电子供体的利用,在不额外增加商品碳源的前提下,提升反硝化性能,对低成本生物脱氮具有重要意义。

图4 常规反硝化过程电子传递途径示意图Fig. 4 The schematic diagram of canonical electron transport pathway of denitrification process
3.2 增强反硝化酶活性
反硝化过程中的电子消耗过程主要依赖与反硝化相关酶的活性,而反硝化酶的活性受到碳源种类和含量、温度、pH和溶解氧等因素的影响。除了优化上述反应条件,在电子传递基础上,还有一些研究探讨了对反硝化酶活性的调控。提高反硝化酶活性通常不是独立的生物化学过程,而是与群落结构优化以及电子传递等要素息息相关。例如,WANG等制备的Fe-Pd/多壁碳纳米管诱导电子流向反硝化代谢和促进电子传递的同时,使得亚硝酸盐还原酶活性提高了203.9%[69]。又有研究发现将废弃活性污泥中提取的胞外聚合物用作氧化还原介体,投加到反硝化生物反应器中,不仅促进了电子传递,还使得硝酸盐还原酶和亚硝酸盐还原酶活性分别提高了29.7%和25.4%[24]。SONG等利用生物炭与聚氨酯海绵复合的新型生物载体,不但提高了反硝化细菌和电活性细菌丰度,而且提高了硝酸盐还原酶的活性[74]。
除了上述采用功能性材料调控反硝化酶活性的方法,物理学的磁场和电场调控也是提高反硝化酶活性的重要手段。例如,WANG等采用物理方法设置微弱磁场,显著增强了以糖蜜为碳源的反硝化系统中各个反硝化酶的活性,并优化了脱氮微生物的群落结构[75]。又有研究证实0.2 V的直流输出电压的弱电刺激能够大幅增强硝酸盐还原酶和亚硝酸盐还原酶活性,使得低碳氮比废水的反硝化脱氮效率显著提升[76]。又如,LAGUM等开发出了一种新型电诱导膜生物反应器,通过增强反硝化酶的活性提高污水的处理效果,同时降低对碳源和碱度的依赖[77]。可见,反硝化酶主导的电子消耗过程虽然是反硝化氮代谢的末端环节,却直接关系着硝酸盐还原为氮气的性能,有必要对其进行进一步精准调控以获得更高效的电子利用效率和反硝化性能。
4 总结与展望
目前,国内外学者针对反硝化脱氮技术开展了大量研究。本文首先介绍了新型无机电子供体驱动的光电营养反硝化体系的构建原理与应用现状,然后着重综述了丙烷等短链气态烷烃、有机废弃物和细胞内碳源等绿色有机电子供体的研究进展,最后归纳总结了基于细胞代谢调控提升电子供体利用效率来促进反硝化的作用原理及应用进展。未来有必要重点关注以下几个方面的研究。
(1)在光电反硝化领域,重点开发低毒价廉和光捕获性能优越的环境友好型光敏剂,在分子层面揭示光敏剂-微生物界面电子转移的微生物学和材料化学机制,研发高效反应器,提高绿色光电子的供给效率,并尝试开展更多的工程规模研究,推进该技术的实际应用进程。
(2)通过优化气液传质、加快关键功能微生物的生长与富集等手段提高烷烃利用型反硝化脱氮性能;从底物结构等角度进一步解析有机废弃物碳源化过程的深层机制,探明发酵液中非碳源物质如辅酶、维生素等对反硝化脱氮的影响;从物种竞争协作等生态学角度研究微生物内碳源积累和利用机制,查明内源反硝化的关键限速步骤。
(3)进一步发展反硝化微生物的细胞调控手段,提升电子利用效率。例如,反硝化微生物细胞内电子受体硝酸盐和代谢中间产物(亚硝酸盐、一氧化氮、氧化亚氮)之间存在对电子的竞争,通过精准调控电子在各个反硝化酶之间的分配比例,有望构建更高效的反硝化生物脱氮系统。
