光照在甲壳动物养殖中的应用研究进展
2024-02-21牛林洋董宏标郑晓婷曾祥兵谭泽加张家松
牛林洋,董宏标,郑晓婷,曾祥兵,谭泽加,张家松
(1 中国水产科学研究院南海水产研究所,农业农村部南海渔业资源开发利用重点实验室,广东 广州 510300;2 水产科学国家级实验教学示范中心 (上海海洋大学),上海 201306;3 海南省海洋渔业资源高效利用与加工重点实验室,海南 陵水572426;4 广州利洋水产科技股份有限公司,广东 广州 510080)
光照是影响水产养殖生产的重要环境因子之一,有3个核心要素:光谱、光照强度、光照周期[1]。光谱为光通过光栅或棱镜进行分光后,按照波长或频率大小依次排列形成的图案[2]。常用人工光源(高压钠灯、荧光灯、金属卤化物灯)通过添加滤光片控制光色,随着LED灯的发展,光的波长可以由计算机控制,具有精度高、使用寿命长的特点[3]。光照强度是指单位面积上接受的光通量[4]。由于介质对光的吸收和散射,光照强度在传播时会衰弱[5]。自然界光照周期由光周期和暗周期构成,动物根据不同的光照周期,进化出不同的生物节律[6]。
在影响甲壳动物养殖的诸多环境因子(光照、温度、pH、盐度、营养等)中,光照为关键因子之一,对甲壳动物生长发育、行为、摄食、存活和繁殖等具有重要影响[7]。水体中甲壳动物会因接收光照信号强弱而产生上浮和潜底等行为[8]。有研究表明,补充额外光源可促进幼虾摄食,加速生长,进而缩短养殖时长[9],适当延长光周期能显著提高甲壳动物幼体摄食率和存活率,保证幼体生长稳定性[10]。因此,提供适宜的光源,对养殖动物苗种培育与养殖生产具有重要现实意义。
本研究从光谱、光照强度、光照周期三方面进行阐述,综合了近年来光照参数对甲壳动物行为、摄食、生长发育及产卵繁殖等方面的研究成果,以期为甲壳动物养殖提供理论及实践参考。
1 光在养殖水体中的传播特点
光源指能发出可见光的物体,可分为自然光源及人工光源[11]。光环境的构成包括光谱成分、光照强度和光照周期。光在水体与空气中传播有较大差异,空气因密度小而对光的吸收与散射相对较小,水体密度为空气的800多倍,因而对可见光有严重的吸收和散射[12]。水体对于自然光的吸收在不同光谱区域各有差异,水体对光谱中紫外(10~380 nm)和红外(780~2 526 nm)部分具有吸收作用,而在可见光谱区段(380~780 nm),对红光(622~780 nm)、黄光(577~597 nm)和浅绿色光(565~577 nm)吸收最强[13]。自然海水中,散射作用导致光波衰减60%,吸收作用导致光波衰减40%;而在养殖水体中,由于浮游植物的光吸收,红光衰减得最多,蓝光次之,绿光衰减最少[14]。甲壳动物养殖过程中,光波在养殖水体中的传播会受到水分子、无机溶质、悬浮物等多种因素影响。相关研究表明,养殖池内海水的透光率随水深的增加而呈现下降趋势,当水深为30 cm时,绿光透光率最大,白光最小,随着水深增加,绿光穿透率始终高于其余波长的光[15]。秦菲[16]研究同样发现,LED光照在养殖水体中会随水体深度的增加而衰减,此外,还会受到总悬浮固体(TSS)和化学需氧量(COD)的影响。根据不同光波在养殖水体中的传播特性,实际生产中,选择适宜的光照参数对甲壳动物养殖尤为重要。
2 甲壳动物的光感觉器官
普遍认为,甲壳动物具有单眼(ocellus)和复眼(compound eye)两种光感受器[17],其中,单眼结构相对简单,仅能感觉光线的强弱,不能呈现具体的像[18];而复眼是大多数甲壳动物成体的视觉器官,不同甲壳动物复眼的形态差异不大,如凡纳滨对虾(Litopenaeusvannamei)[19]、日本沼虾(Macrobrachiumnipponense)[20]、锯缘青蟹(Scyllaserrata)[21]、三疣梭子蟹(Portunustrituberculatus)[22]复眼均呈半球形。凡纳滨对虾复眼经H&E染色可显示复眼由许多小眼构成,超微结构由外至内分为折光系统、感光系统和色素细胞三部分,其中折光系统包括角膜、角膜生成细胞和晶锥;感光系统由众多小视网膜突起的微纤毛组成,微纤毛的排列顺序在光、暗环境下不同,如罗氏沼虾微纤毛在自然光环境下排列凌乱,而黑暗环境下则排列整齐[23];色素细胞则具有遮挡光线的作用[17]。甲壳动物的感光单元结构为感光细胞,由感光部、细胞体部和轴突构成[24]。感光细胞通过吸收和释放存在于潴泡囊和线粒体中的Ca2+改变其体积,光敏感性随着Ca2+浓度升高而降低[25-26]。甲壳动物对光色的敏感程度与其色素细胞对光色的过滤性相关[27];自然光环境下,日本沼虾视觉细胞中色素颗粒分散到细胞中,感杆束边缘小囊泡数量增多,溶酶体出现并靠近感杆束[28],这与中华绒螯蟹(Eriocheirsinensis)光环境下视觉细胞观察结果相似[29]。Sanudin等[30]研究发现,凡纳滨对虾幼体摄食主要依赖复眼进行,光照是影响其幼体摄食的主要因素之一,随着不断生长发育,复眼的分辨力逐渐增强,其主动摄食的能力亦逐步提高;进入成体阶段后,复眼在探测食物过程中的作用逐渐减弱,而由化学感受器起主导作用[31]。
甲壳动物眼柄神经内分泌系统为X器官-窦腺复合体(XO-SG complex),是甲壳动物内分泌调节中心[32]。凡纳滨对虾复眼经H&E染色显示(图1)[19]。
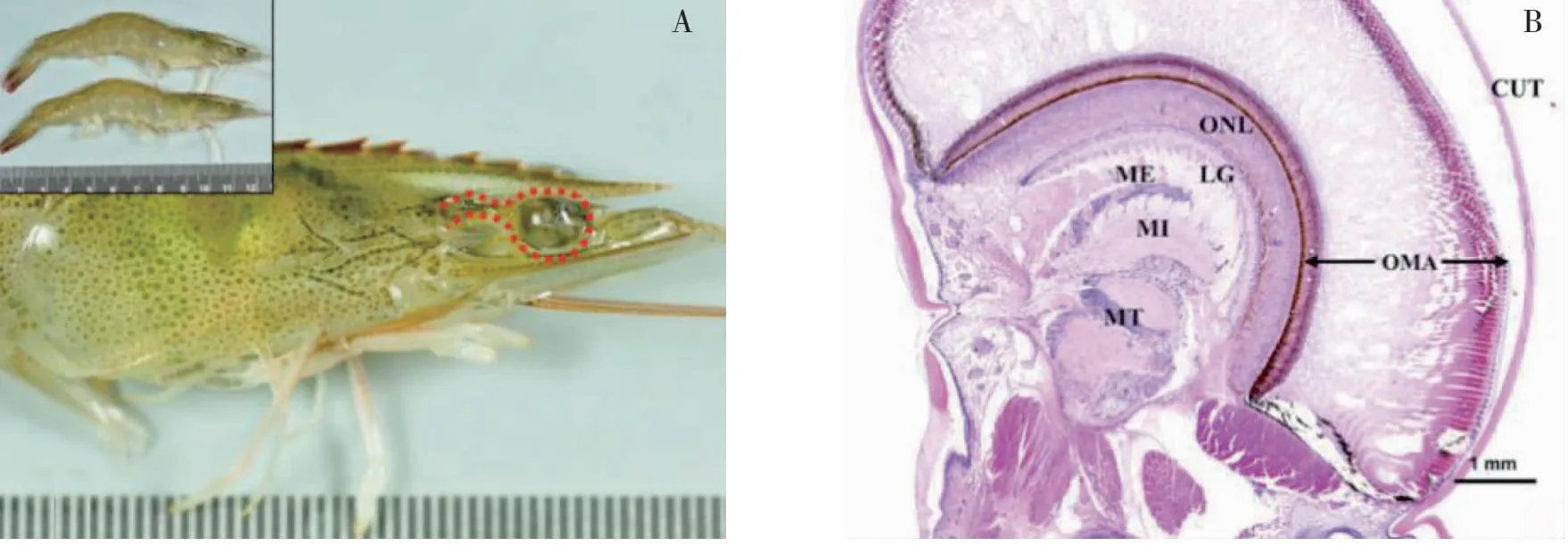
注:H&E,染色显示的结构;CUT,角质层;OMA,小眼;ONL, 视神经层;LG,神经节层;ME,外髓质;MI,内髓质;MT,终髓
从最外层角质层向内依次为角膜、小眼、视神经层,神经节层、外髓质、内髓质、终髓。窦腺位于内髓和终髓之间,储存蜕皮、生长、性成熟和代谢等相关激素[33]。光照的变化会引起甲壳动物体内激素或神经递质水平的变化,进而影响其生理状况[34]。Nakamura[35]研究发现,光照能够通过影响视神经脊髓终端细胞活性以调节日本囊对虾(Marsupenaeusjaponicus)摄食节律。此外,连续光照能够引起克氏原螯虾(Procambarusclarkii)眼柄、大脑、丘脑神经节、食管下神经节中5-羟色胺(5-HT)浓度显著升高,进而刺激克氏原螯虾的卵母细胞成熟[36]。
3 光照对甲壳动物养殖生产的影响
3.1 光谱成分对甲壳动物的影响
甲壳动物对不同波长的光敏感程度因种而异,而同物种不同的生长阶段对于光谱的敏感度也有差异[37]。据报道,摘除触角的日本沼虾在红光和绿光光照区域的平均摄食量远大于蓝光和黄光区域,其趋光行为在红光下频率最高[38]。克氏原螯虾拥有昼夜两套感光系统,其红光感光系统发育形成较蓝光感光系统缓慢,进而导致幼体阶段对蓝光敏感,成虾阶段则对红光敏感[39]。长毛明对虾(Fenneropenaeuspenicillatus)同样具有双重光感受器,分别对蓝光和红光两种波长的光敏感[40]。另有研究发现,相同光照强度下,三疣梭子蟹在红光下的趋光率最高,黄光下趋光率最低[41]。
不同种类甲壳动物适宜生长的光谱环境有所差异。Emmerson等[42]研究发现,蓝光和绿光可改善印度明对虾(Fenneropenaeusindicus)成虾的生长特性。采用金卤灯为光源养殖凡纳滨对虾时,与其他光源组相比,不延长光照时间就能显著提高对虾体内虾青素含量,改善其体色,增加对虾摄食率并促进生长[10]。表明养殖过程中补充光源时选择适宜的光谱有助于促进养殖对象生长。
光照是除营养外影响甲壳动物繁殖与幼体发育的另一重要因素[43]。黑暗条件下红条鞭腕虾(Lysmatavittata)亲虾的抱卵率为0,自然光照下抱卵率可提高到(95.8±3.6)%,适宜的光照能够对红条鞭腕虾亲虾抱卵起到促进作用[44]。光谱对甲壳动物繁殖的影响亦是因种而异。研究发现,蓝光环境能够促进东方巨对虾(Penaeusplebejus)卵巢发育[45];绿光和蓝光协同条件下,印度明对虾产卵量显著高于自然光条件[42]。光谱对主要甲壳动物生长生理的影响如表1所示。
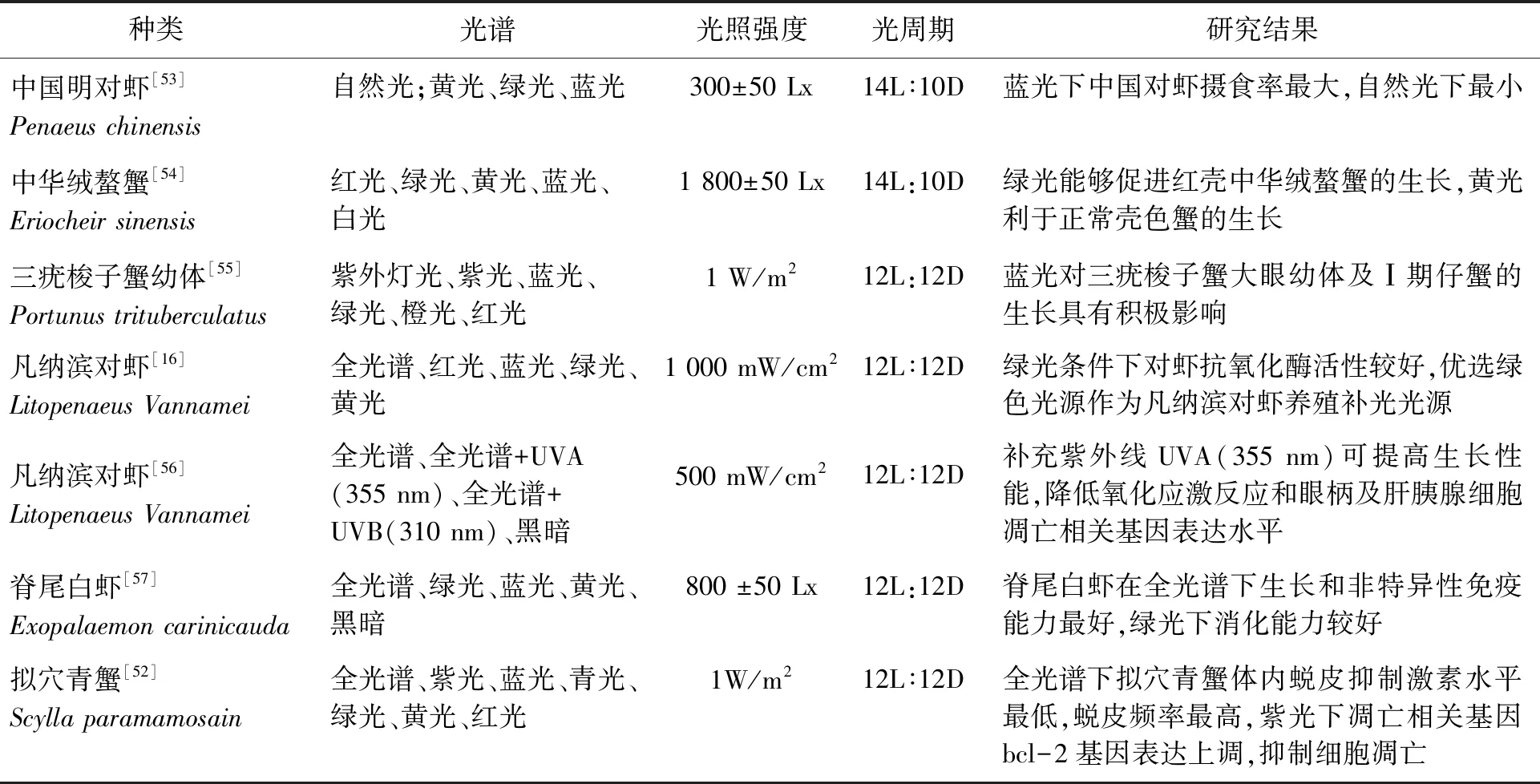
表1 光谱对主要甲壳动物生长影响
光谱通过影响甲壳动物的生理机能调控其生长与免疫。研究表明,甲壳动物的内在生理机制极易受到外界环境因素的刺激[46],一些甲壳动物如凡纳滨对虾、斑节对虾(Penaeusmonodon)、中华绒螯蟹等在受到环境因素胁迫,机体内的活性氧(ROS)增加,常通过分泌超氧化物歧化酶(SOD)、过氧化氢酶(CAT)等抗氧化相关酶以分解体内过多的活性氧,减少机体损伤[47]。蓝光条件下凡纳滨对虾胃蛋白酶(PEP)和胰蛋白酶(TRY)显著高于其他光照组,蓝光和绿光条件下脂肪酶(LPS)活性显著高于其他组,结果表明,蓝光和绿光通过调控消化酶的分泌,促进对虾的消化吸收[48]。综上,不同种类及生理阶段甲壳动物需要提供不同的光谱范围,使其能更快响应环境应激,维持内环境稳态,提高养殖效率。
光谱对甲壳动物蜕皮的影响与其对不同波长的光敏感性不同有关。甲壳动物蜕皮受内分泌系统和神经系统共同调节[49],同时也受到环境因子(光照、季节变化等)的刺激[50]。普遍认为甲壳动物蜕皮是位于头胸部前端的Y器官分泌蜕皮激素(molting hormone,MH)与位于眼柄的X器官窦腺复合体分泌蜕皮抑制激素(molt-inhibiting hormone ,MIH)互相拮抗作用的结果[51]。全光谱下拟穴青蟹(Scyllaparamamosain)体内MIH水平最低,蜕皮频率最高,紫光和黄光下MIH显著上调,蜕皮频率最低[52],表明光谱通过调控蜕皮相关激素水平,来影响甲壳动物的蜕皮频率与生长。研究发现,凡纳滨对虾在蓝光下蜕皮总次数显著高于其他光谱组,且蓝、绿光下对虾的生长速度较快[48]。
3.2 光照强度对甲壳动物的影响
不同种类与生长阶段的甲壳动物最适光照强度具有差异,其行为活动因光照强度而发生改变。已有研究表明,幼体阶段的甲壳动物对光照具有正反应,而成体阶段对光照则呈负反应[58]。光照会影响甲壳动物的垂直移动行为,白天光照强度减弱和夜晚的光照强度增强均会引起对虾的上浮或潜底[59]。如三疣梭子蟹喜弱光,惧强光,白天多潜伏在海底层或活动于靠近底层的水域,夜间或生殖抱卵期才游到上层[60]。自然条件下,大多数种类对虾同样于白天潜底,夜间上浮[61]。然而Kenyon等[62]研究发现,仔虾的上浮和潜底可能不受光照强度的影响,褐虾(Penaeusesculentus)仔虾在光暗环境下皆呈浮游状态,而不下潜。这与褐虾仔虾的复眼结构与对环境的高适应性有关。甲壳动物的摄食、发育及蜕皮等活动与其行为变化都有相关性。
关于光照强度与甲壳动物摄食之间的关系,Petersen等[63]提出了两种摄食模型:峰值摄食模型曲线和S型摄食模型曲线,其中,符合峰值摄食模型曲线的动物主要依赖视觉进行摄食,存在一个适宜的光照强度范围使动物的摄食量达到最大,光照强度过高或过低都会影响动物的摄食;S型摄食模型曲线为随着光照强度的减弱,动物的摄食量逐步增高,当光照强度降低到一定程度,摄食量达到最大且基本恒定。大多数虾蟹类动物的摄食模型属于峰值摄食模型。无论是处于哪一期的罗氏沼虾幼体,在完全黑暗的条件下都几乎不摄食,随着光照强度的增大,摄食率迅速升高,而到1 000 Lx以上摄食率变化不大[64]。研究发现高光照度下(5 000 Lx)中国明对虾的呼吸、排泄耗能较高,而用于生长的能量比例降低[65]。中国对虾在夜间的摄食量要高于在日间,且在夜间有两个摄食高峰,分别是18:00—21:00和03:00—06:00[66]。养殖车间需要根据相应物种摄食所需光照强度灵活调整光照条件,育苗阶段尤为重要。
光照强度通过影响食物转化效率和能量分配比例调控对虾生长发育[67]。中国明对虾在5 500 Lx照度下的特定生长率(SGR)和食物转化率(FCE)最低,在300 Lx照度下最高[53]。Neal等[68]研究发现,自然光条件下池塘养殖凡纳滨对虾养殖产量比弱光池塘中高出48%,分析其原因可能在于光照对轮虫和其他浮游动物的生长具有促进作用,为对虾提供了天然的饵料来源。相同光周期(14L∶10D)下,光照度波动组(2 700±1 800 Lx)凡纳滨对虾的摄食率(FI)显著高于恒定光照度组(2 700 Lx),而饲料转化率(FCR)低于恒定光照组,且2 700±1 800 Lx光照度下凡纳滨对虾的生长状况优于其他波动光强组[69]。养殖车间内适宜的光照强度对提高甲壳动物幼体存活率及摄食率有重要作用。
甲壳动物一般呈阶梯式生长,即在蜕皮后快速生长,蜕皮之后到下一次蜕皮之前其尺寸几乎很少增加[70]。对虾的蜕皮和生长是结合进行的,但蜕皮是一种节律性极强的活动,主要由体内激素调节,生长则受到周围环境中饵料的影响[71]。饥饿状态下一些甲壳动物依旧能够蜕皮,但蜕皮后的干重和能量比摄食组的少[72]。三疣梭子蟹幼体生长适宜的光照强度为9.96 μmol /(m2·s),该光强能够保证幼蟹的存活率、蜕皮率及生长性能[73]。
当前,光照强度对于甲壳动物繁育影响的相关研究仍不足。研究表明强光(11 00 Lx)能够抑制墨吉明对虾(Penaeusmerguiensis)卵巢成熟并降低其产卵量,弱光环境(2 Lx)下则相反[74]。弱光条件下可以促进红额角对虾(Litopenaeusstylirostris)和凡纳滨对虾的繁殖[75]。交配后的拟穴青蟹在完全黑暗条件下其卵巢中类胡萝卜素及蛋白质含量低于低光强(7.00±1.50 Lx)环境[76]。低光强或暗光条件下对大部分甲壳动物的性腺发育及繁殖有利,光强过高则不利于甲壳动物性腺的发育、成熟及产卵。光强对主要甲壳动物生长的影响如表2。
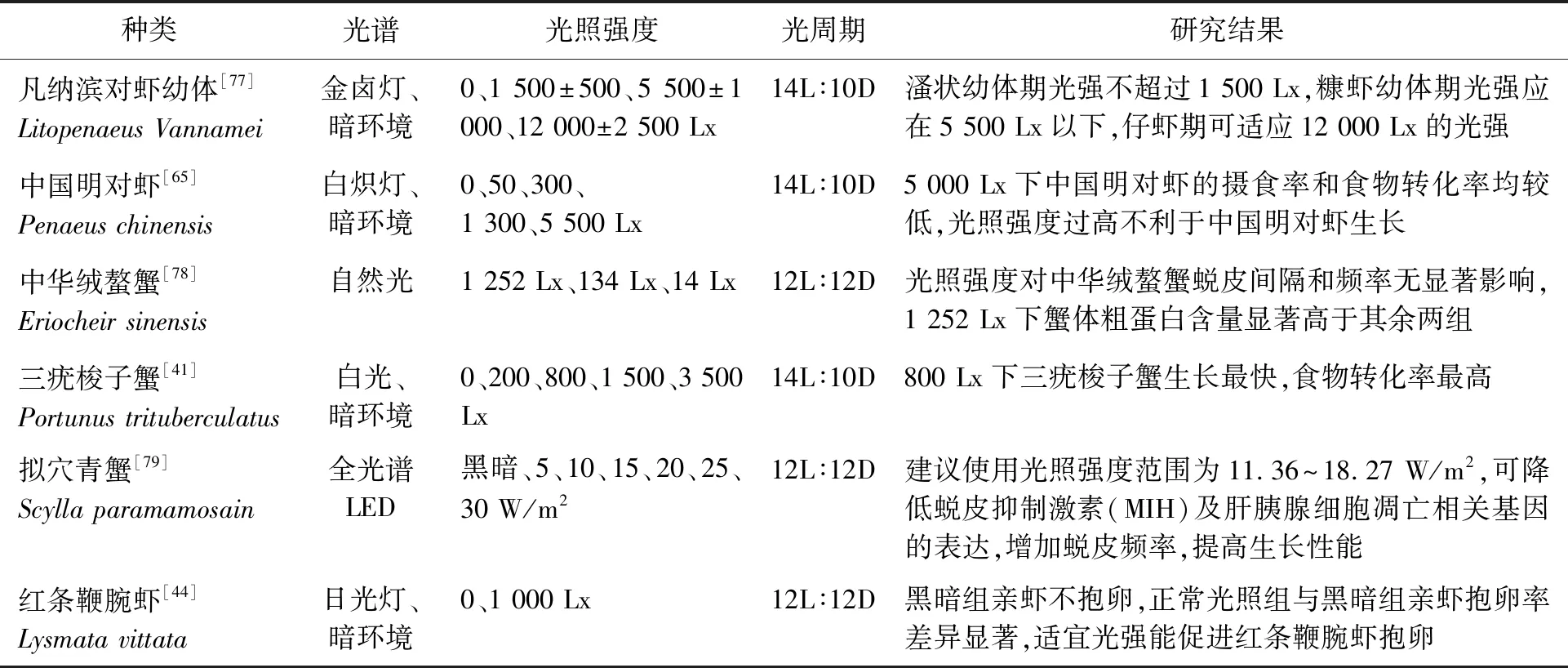
表2 光照强度对主要甲壳动物生长的影响
3.3 光照周期对甲壳动物的影响
生态环境中大多数的生物都经历着昼夜交替。因此,许多生物也进化出了相应的生理感官和行为活动来响应这种昼夜变化并及时对自身行为做出调整,如感光器官和生物钟机制[80]。普遍认为,眼柄组织是甲壳动物的特有感光器官,光线对其神经分泌细胞的分泌活动具有调控作用[81]。甲壳动物在不同光周期下的生长、代谢、存活和繁殖具有物种差异性[82]。
光周期是光环境中重要的因素之一,探究不同光周期下甲壳动物的摄食及生长很有必要。Sanudin等[83]研究发现,小规格(<0.5 cm)凡纳滨对虾培育时应延长光周期[12L(Light)∶12D(Dark)~24L∶0D],而大规格(>1 cm)对虾的摄食行为、生长状况和存活率不受光周期的影响。相关研究表明凡纳滨对虾养成期摄食活动受光周期影响较小,表明其摄食行为是一种内源性行为[84]。You等[10]研究发现,连续光照条件下,凡纳滨对虾的摄食率并没有显著的提高。在罗氏沼虾养殖过程中,适当延长光照周期,能够增加其日摄食量,促进幼体生长发育[85]。Andrés等[86]研究发现,持续黑暗导致远海梭子蟹(Portunuspelagicus)幼体存活率和发育率降低,明确远海梭子蟹幼体最适生长光周期为18L∶ 6D。
光照周期的改变对甲壳动物蜕皮同样产生影响。延长光照周期会抑制甲壳动物蜕皮抑制激素(MIH)的合成和释放,从而缩短其蜕皮周期,短时间光照则相反[87]。光照周期延长使日本囊对虾体内MIH激素水平显著升高,蜕皮激素(EH)水平下降,黑暗条件MIH浓度显著低于其他光照组,EH浓度显著高于光照条件,结果表明黑暗状态有利于日本囊对虾蜕壳[88]。黑暗条件下中华绒螯蟹大眼幼体蜕壳率降低、成活率低;光照可刺激并促进大眼幼体的蜕壳与生长,但在仔蟹I期以后黑暗条件下有利于其蜕壳生长[89]。
在甲壳动物生物学研究中,常通过调控光照周期来影响其产卵繁殖[90]。适当延长光周期(16L∶8D)可促进克氏原螯虾雌虾性腺发育,而过长的光照(20L∶4D)对雌虾性腺发育会有抑制作用[91]。Matsuda等[92]研究发现,水温为25 ℃时日本囊对虾卵巢发育和产卵最佳光照时间是12.5~14 h。据Akinouchi 等[93]报道,光照度为2~5 Lx,光照周期为17L∶7D时,斑节对虾的产卵量和孵化率均高于其他光照组。光照周期对主要甲壳动物生长的影响如表3所示。

表3 光照周期对主要甲壳动物生长的影响
4 问题与展望
4.1 光照参数的优化
光环境是设施渔业生产系统构建需要重视的因素之一,其在调控水产动物幼体发育、摄食行为及产卵繁殖等方面具有重要意义,如何营造并利用好光环境是实现水产养殖高效生产的重要环节。
(1)光谱的选择需要考虑到不同波长光照在养殖水体中的传播特性,同时需要注意养殖对象对光照的敏感程度。自然光源及传统人工光源发出的光谱范围广,缺乏对光谱的精准调控。(2)光照强度的设置对甲壳动物的摄食、生长及繁育有重要作用。不合理的光强将引起甲壳动物的应激反应,导致其生长受到阻滞。(3)光照周期对甲壳动物的摄食、蜕皮及产卵繁殖都有一定的影响。甲壳动物养殖是现代水产养殖中重要的组成部分,不同种类甲壳动物的最适光环境参数都有差异,生产过程中需要根据品种特性灵活调整光照参数。此外,光照参数对甲壳动物内分泌系统中各种激素和神经递质的生理调节仍然需要更为全面深入的研究。结合代谢组学、多组学、表观遗传学、全基因组关联分析等技术,全方面探索光照对甲壳动物相关基因调控的具体分子通路和作用机制,可作为未来探究光照对甲壳动物生长、发育、繁殖等的研究方向。
4.2 光源的选择
随着设施渔业智能化发展,工厂化养殖将是渔业智能化发展的重要方向。相较于传统养殖方式(池塘养殖、网箱养殖)具有节水节地、高产高效、生态环保、全程可控等优点。目前,大多数工厂化养殖车间用的还是传统光源,例如金卤灯、荧光灯等,这些光源不仅耗能高,长时间照射还会产生较高的温度,而且传统光源还含有对养殖动物生长不利的光谱成分,尤其会对甲壳动物的生长造成一定的负面影响。自20世纪90年代以来,国内外学者围绕光照对水产动物的影响逐渐开展相关研究,从传统光源对水生动物的影响逐步转变为更加精准的光谱、光周期和光照强度的调控,人们逐渐意识到LED光源对水产养殖的增益效果。使用LED养殖灯对甲壳动物生长光环境进行精准调控是未来对虾养殖效率的关键提升环节之一。设计针对养殖动物各生长时期(如对虾标苗期、标粗期、养成期、繁殖期等)的LED补光策略,以满足养殖动物对光照的需求。目前,大多数养殖车间对于LED补光系统中光照参数的搭配设计不完善,造成资源、成本的浪费。因此,建立设施渔业系统中对光谱、光强、光周期的需求体系,是未来推广智能化设施渔业的重要步骤之一。
5 结论
水产养殖中,养殖水体直接影响光的传播,甲壳动物接收到的光是经过空气与水体散射和吸收过后的,相关研究表明,绿光与蓝光在养殖水体中的穿透性较好。本综述中,多项研究表明,绿光与蓝光能够促进甲壳动物的摄食、幼体生长、蜕皮、产卵,增强抗氧化能力。不同甲壳动物对光强的适应性有所差异,如中国明对虾在300 Lx光照度下生长最快,三疣梭子蟹在800 Lx下生长最快,凡纳滨对虾能够适应较高的光照度,可达到12 000 Lx。甲壳动物产卵阶段,应使用较低的光强,有利于其卵巢发育。适当延长光照周期(如18L∶6D),对大多数甲壳动物的生长及产卵繁殖均有利,而不同种类甲壳动物的蜕皮行为对光照周期的响应有差异。实际生产阶段,应兼顾光照三要素的作用,不同时期选择合适的光照参数,以适应动物的生长。
□
