感赤星病与健康烟叶在腈菌唑应用后的叶际微生态
2024-02-05蔡刘体汪汉成陈兴江
张 艺,王 丰,蔡刘体,汪汉成,熊 晶,陈兴江
(1.贵州大学 农学院,贵州 贵阳 550025; 2.贵州省烟草科学研究院,贵州 贵阳 550081; 3.贵州省烟草公司毕节市公司,贵州 毕节 551700)
烟草赤星病(tobacco brown spot)是由子囊菌亚门链格孢属(Alternariasp.)真菌引起的烟草叶斑病,主要发生在烟叶成熟期,典型症状为褐色同心轮纹病斑,温湿度适宜时可大面积流行成灾[1-3]。目前,烟草赤星病的防治主要以化学防治为主。腈菌唑是一种高效、低毒、广谱、兼具保护和治疗作用的三唑类杀菌剂,对由子囊菌、担子菌引起的病害具有较好防效[4-5],主要通过抑制真菌麦角甾醇的生物合成、破坏细胞膜的完整性和流动性,以及细胞的抗逆性达到杀菌目的[6]。目前,该药剂在我国主要登记用于子囊菌引起的黑斑病、叶斑病、白粉病等病害的防治。在烟叶生产上,该药剂主要用于烟草白粉病、赤星病的防治[7]。
植物叶际作为微生物的主要栖息地,为微生物的定殖提供了良好的生存环境。叶际微生物的菌群结构与病害的发生密切相关[8-9]。已有的研究发现,健康与感病烟叶组织叶际微生物在菌群结构与多样性上均存在差异。目前,烟草赤星病危害期其叶际真菌和细菌的菌群组成已多有报道[10]。赤星病发生初期时,不同成熟度健康与感病烟叶的优势真菌和细菌具有相似性,真菌优势菌属均包括链格孢属、亚隔孢壳属、枝孢霉属,细菌优势菌属均包括假单胞菌属和鞘氨醇单胞菌属细菌[11]。赤星病发生中后期时,感赤星病烟叶叶际细菌群落中还存在大量泛菌属细菌[12]。病害的发生会改变叶际微生物群落结构与多样性,同时,叶际微生物的菌群组成也会受寄主叶片成熟度、环境及药剂的使用而发生变化[13]。已有的研究发现,菌核净可显著抑制烟叶叶际鞘脂单胞菌属、黄色杆菌属和沙雷氏菌属菌群的丰度[14],甲氧基丙烯酸酯类的醚菌酯在防治烟草赤星病的过程中可显著抑制链格孢属、假单胞菌属、泛菌属等菌属的丰度[15]。当前,腈菌唑在我国用于烟草叶斑类病害的防控已有多年,其对白粉病的防控效果优于赤星病。作为烟草叶斑类病害防控的常用药剂,其在赤星病发生期应用时的叶际微生态机制却缺乏认识,特别是其应用后,健康与感病烟叶叶际微生物的菌群结构与代谢功能的调控规律仍不清楚。了解杀菌剂应用后叶际微生物的响应规律对指导药剂的科学使用具有重要意义。
为此,本文采用高通量测序技术和Biolog代谢表型技术,分析腈菌唑施药后健康烟叶与感赤星病烟叶叶际真菌和细菌的群落结构、多样性及代谢功能,旨在了解腈菌唑施用不同时期内烟叶叶际微生物的变化规律,现将结果报道如下。
1 材料与方法
1.1 试验材料
供试品种云烟105,购自云南省烟草公司玉溪市公司;12.50%腈菌唑微乳剂(ME),购自北京东旺公司;GeneJET胶回收试剂盒和Ion Plus Fragment Library Kit 48 rxns建库试剂盒,均购自Thermo Scientific公司。多功能喷雾器,购自贵州黔丰源农业科技开发有限公司;自动气象站,配有温度计(MC-KWS)、雨量计(MC-YL)和湿度计(MC-KWS)等,购自北京新红科技有限公司;Biolog ECO代谢板,购自美国Biolog公司(USA, CA, Hayward)。
1.2 田间施药处理
1.2.1 试验设计
试验于2020年8月底在贵州省威宁县黑石头镇进行,选取烟株长势一致的烟田划分小区,各试验小区随机排列,每小区60株烟株,小区设3次重复,各小区之间设保护行。12.50%腈菌唑微乳剂(ME)的每667 m2用量为60 mL,每667 m2用水量为60 L,试验时采用多功能喷雾器将药液均匀喷雾至烟叶表面,直至液滴流失。使用自动气象站检测并记录试验过程中的降雨量、温度和空气相对湿度,并分别于施药前0 d、施药后5、10、15 d,各小区随机选取10株烟株调查烟叶发病情况,计算各时期的病情指数[16]。
1.2.2 样品采集
于施药前0 d,施药后5、10、15 d分别取样,采用消毒剪刀分别剪取烟株中下部相同部位的感病与健康组织烟叶样品,分别装入50 mL无菌离心管中,每处理3次重复。样品采集后放入低温保存箱,并迅速带回实验室用于后续研究,样品编号如表1所示。
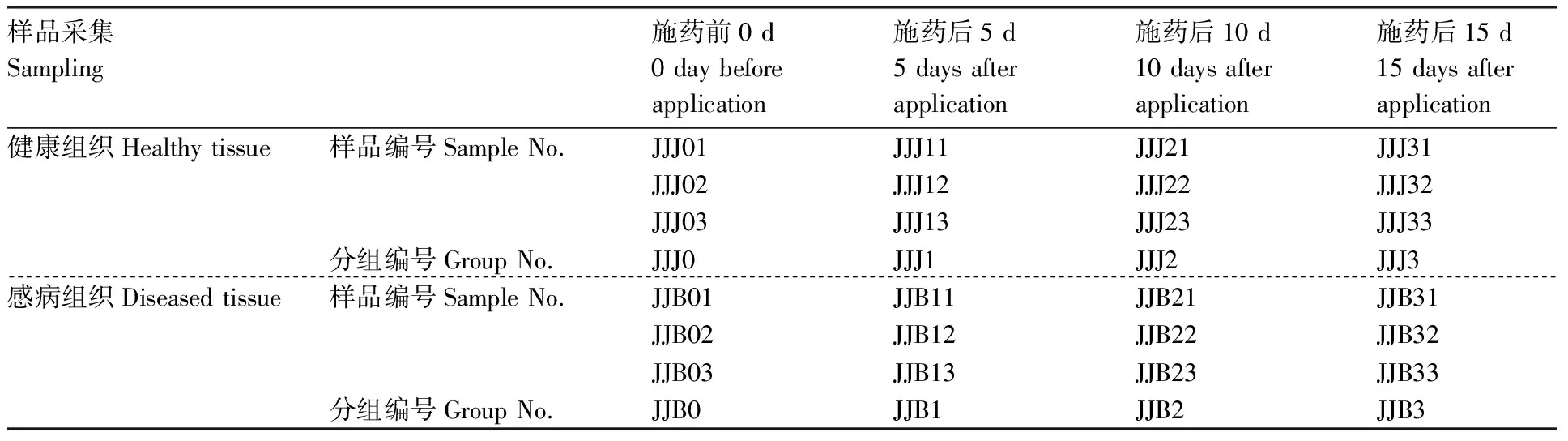
表1 样品采集信息
1.3 烟叶叶际微生物菌群结构与代谢功能差异分析
1.3.1 叶际真菌和细菌群落结构与多样性
采用CTAB[17]法提取烟叶叶际微生物的基因组DNA,使用NanoDrop 2000测定DNA浓度和纯度,检测合格后用于构建文库。以上述样品的总DNA为模板,以真菌引物ITS1-5F-F(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS1-1F-R(5′-GCTGCGTTCTTCATCGATGC-3′)对叶际微生物基因组DNA ITS1区域进行PCR扩增。以细菌引物515F(5′-GTGCCAGCMGCCGCGGTAA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对样品叶际微生物基因组DNA 16S-V4区域进行PCR扩增。PCR反应程序和扩增体系参照文献[18-19]的方法进行。扩增产物回收后,使用Ion Plus Fragment Library Kit 48 rxns建库试剂盒进行文库的构建,采用Illumina MiSeq测序平台对PCR扩增产物进行双端测序分析,以上分析均在北京诺禾致源科技股份有限公司完成。
1.3.2 叶际微生物代谢功能
分别取施药前后不同部位烟叶混合样品各1 g,将其置于盛有50 mL 0.8%的无菌生理盐水的100 mL三角瓶中,28 ℃下以180 r·min-1振荡摇培2 h,取出后静置30 min,取100 μL上清液依次分别加入到ECO代谢板的测试孔中[20]。将ECO代谢板置于OmniLog恒温培养箱,28 ℃培养7 d,采用Biolog D5E_OKA_data.exe软件收集烟叶叶际微生物在生长过程中代谢孔内颜色变化值,收集后使用HemI软件制作热图分析样品叶际微生物的代谢功能[21]。
1.4 数据处理与统计分析
使用Excel 2019软件进行数据处理及统计分析。测序数据通过FLASH和Trimmomatie软件进行过滤优化和双端序列连接[21]。优质序列
使用Uparse软件(Uparse v7.0.1001)对相似度≥97%的序列进行OTU(operational taxonomic units)聚类,并在聚类过程去除单序列和嵌合体[22]。真菌和细菌分别通过UNIT数据库[23]和SILVA的SSUrRNA数据库进行注释[24]。采用QIIME V1.9.1软件计算Alpha多样性指数[25],运用SPSS软件对真菌和细菌的Alpha多样性指数进行差异显著性分析。利用R软件(Version 2.15.3)绘制真、细菌稀释曲线图、物种积累箱形图、门属水平相对丰度图、借环境因子Spearman相关性图,分析样品微生物群落与多样性。
2 结果与分析
2.1 感赤星病与健康烟叶叶际微生物群落结构与多样性的差异
2.1.1 测序深度分析
本次测序共8组24个样品,在真菌群落中,测序深度接近800、OTU数量接近300时曲线物种累积箱和趋于平缓(图1-A、C)。在细菌群落中,测序深度和OTU数量均接近300时趋于平缓(图1-B、D)。二者趋于平缓表明测序深度已经基本覆盖了样品中绝大多数物种,抽样充分,可以进行数据分析。
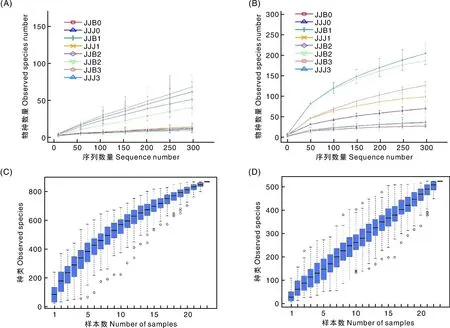
A,真菌OTU测序深度;B,细菌OTU测序深度;C,真菌水平物种累积箱形图;D细菌水平物种累积箱形图。A, OTU sequencing depth of fungi; B, OTU sequencing depth of bacteria; C, Horizontal species accumulation of fungi; D, Horizontal species accumulation of bacteria.图1 烟叶样品真菌、细菌的OTU测序深度和水平物种累积箱形图Fig.1 Box diagram of OTU sequencing depth and horizontal species accumulation for fungi and bacteria in tobacco samples
2.1.2 叶际真菌和细菌Alpha多样性差异
腈菌唑处理前后感病烟叶与健康烟叶叶际真菌和细菌群落的覆盖度指数分别在0.97、0.81以上,表明测序数据合理,可以真实、合理地反映真菌和细菌的群落分布。在真菌群落结构中,施药前后感病烟叶叶际真菌群落的多样性和丰富度指数均低于健康烟叶。感病烟叶的多样性指数在施药5 d时升高,10 d时降低,15 d时升高,丰富度指数则一直保持上升趋势;健康烟叶的多样性指数和丰富度指数均保持上升趋势(表2)。

表2 烟叶叶际真菌和细菌Alpha多样性差异(OTU水平)
在细菌群落结构中,施药前后感病烟叶叶际细菌群落的多样性指数均高于健康烟叶,感病烟叶和健康烟叶的多样性指数在施药5 d时降低,10、15 d时升高。感病烟叶在施药5 d时丰富度指数降低,而后保持上升趋势;健康烟叶丰富度指数一直保持上升趋势(表2)。
2.1.3 施药不同时期的群落多样性与相对丰度的差异
真菌群落在门水平上,子囊菌门(Ascomycota)、担子菌门(Basidiomycota)为施药前后感赤星病烟叶与健康烟叶的优势菌群。施药前后感病烟叶的子囊菌门相对丰度均高于健康烟叶,而担子菌门与之相反。施药前感病烟叶和健康烟叶的子囊菌门相对丰度分别为93.67%、63.90%,施药后相对丰度均降低。施药前感病烟叶和健康烟叶的担子菌门相对丰度分别为2.52%、6.77%,施药后感病烟叶的担子菌门相对丰度在施药5、10 d时降低,15 d时增加;健康烟叶担子菌门相对丰度在施药5 d时降低,10 d、15 d时增加(图2-A)。

A、C分别为真菌门水平和属水平;B、D分别为细菌门水平和属水平。A and C are at the fungal phylum and genus level, respectively; B and D are at the bacterial phylum and genus level, respectively.图2 感病与健康烟叶真菌和细菌群落组成相对丰度图Fig.2 Relative abundance of fungal and bacterial community composition of diseased and healthy tobacco leaves
细菌群落在门水平上,变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)为施药前后感赤星病烟叶与健康烟叶的优势菌群,且施药前后感病烟叶的变形菌门相对丰度均高于健康烟叶。施药前感病烟叶和健康烟叶的变形菌门相对丰度分别为38.81%、6.04%,感病与健康烟叶施药后5 d、10 d时相对丰度均降低,15 d时均增加。施药前感病烟叶和健康烟叶的厚壁菌门相对丰度分别为0.56%、16.33%,施药后感病烟叶相对丰度增加,而健康烟叶施药后相对丰度降低(图2-B)。
施药前后感赤星病烟叶与健康烟叶的主要真菌菌属为链格孢属(Alternaria)、谷菌根菌属(Archaeorhizomyces)、Boeremia、枝孢霉属(Cladosporium)、Symmetrospora、亚隔孢壳属(Didymella)、附球菌属(Epicoccum),其中链格孢属为优势菌属,施药前感赤星病烟叶中链格孢属相对丰度(84.40%)远高于健康烟叶(36.48%)。施药5 d时,感病烟叶与健康烟叶中相对丰度降低的菌属为链格孢属(分别降低了5.52%、11.40%)、枝孢霉属(0.80%、9.72%)、Boeremia(0.35%、5.29%)、Symmetrospora(1.57%、0.73%);而亚隔孢壳属(0.37%、0.1%)相对丰度增加。施药10 d时,感病与健康烟叶中相对丰度降低的菌属为枝孢霉属(4.93%、2.37%),以及感病烟叶中的Boeremia(0.15%),健康烟叶中的附球菌属(0.59%)、链格孢属(6.13%);感病烟叶相对丰度增加的菌属为链格孢属(11.59%)、Symmetrospora(0.02%)、亚隔孢壳属(0.02%);健康烟叶相对丰度增加的菌属为谷菌根菌属(4.15%)、Boeremia(0.09%)、Symmetrospora(5.89%)。施药15 d时,感病与健康烟叶中相对丰度降低的菌属为链格孢属(23.08%、8.43%),健康烟叶相对丰度降低的菌属为枝孢霉属(0.24%)、Symmetrospora(0.79%)、亚隔孢壳属(0.03%)、附球菌属(0.87%);感病与健康烟叶中相对丰度增加的菌属为Boeremia(0.74%、0.20%),感病烟叶相对丰度增加的菌属为亚隔孢壳属(1.21%)、附球菌属(0.60%),健康烟叶相对丰度增加的菌属为谷菌根菌属(1.21%)(图2-C)。
细菌群落在属水平上,施药前后感赤星病烟叶与健康烟叶的主要菌属为科萨克氏菌属(Kosakonia)、乳球菌属(Lactococcus)、泛菌属(Pantoea)、假单胞菌属(Pseudomonas)、魏斯氏菌属(Weissella)、鞘脂单胞菌属(Sphingomonas)、马赛菌属(Massilia),甲基杆菌属(Methylobacterium)、其中科萨克氏菌属为优势菌属,施药前感赤星病烟叶中科萨克氏菌属相对丰度(23.38%)远高于健康烟叶(3.47%)。施药5 d时,感病烟叶中相对丰度降低的菌属为魏斯氏菌属(0.1%)、假单胞菌属(4.92%)、马赛菌属(2.01%),健康烟叶相对丰度降低的菌属为科萨克氏菌属(2.46%)、泛菌属(1.90%);感病烟叶相对丰度增加的菌属为科萨克氏菌属(4.14%)、乳球菌属(18.12%)、泛菌属(6.04%)、甲基杆菌属(0.22%),健康烟叶相对丰度增加的有假单胞菌属(1.23%)、乳球菌属(0.78%)。施药10 d时,感病与健康烟叶中相对丰度降低的菌属为科萨克氏菌属(25.84%、0.78%)、乳球菌属(17.45%、0.78%)、泛菌属(4.70%、0.22%),以及感病烟叶的甲基杆菌属(0.11%);感病烟叶相对丰度增加的菌属为鞘脂单胞菌属(0.89%)、假单胞菌属(8.05%)、马赛菌属(1.23%),健康烟叶相对丰度增加的菌属为鞘脂单胞菌属(0.22%)、甲基杆菌属(0.56%)。施药15 d时,感病与健康烟叶中相对丰度降低的菌属为鞘脂单胞菌属(0.34%、0.11%)以及感病烟叶的乳球菌属(0.67%)、假单胞菌属(6.38%)、马赛菌属(1.01%),健康烟叶的鞘脂单胞菌属(0.11%)、甲基杆菌属(0.34%);感病烟叶相对丰度增加的菌属为科萨克氏菌属(12.42%)、泛菌属(17.00%),健康烟叶相对丰度增加的菌属为科萨克氏菌属(3.92%)、泛菌属(9.62%)、假单胞菌属(2.46%)、马赛菌属(0.11%)(图2-D)。
2.1.4 环境因子对烟草叶际真菌和细菌群落结构的影响
在试验开展期间该地区长时间有小雨。自动气象站测量结果显示,在施药前0 d、施药后5 d、10 d、15 d时的降雨量分别为13.60、19.00、11.30和25.40 mm;温度分别为18.32、18.97、17.51和16.54 ℃;空气相对湿度分别为82.49%、83.81%、78.99%和90.98%;病情指数分别为44.44、48.14、63.58、70.55(表3)。
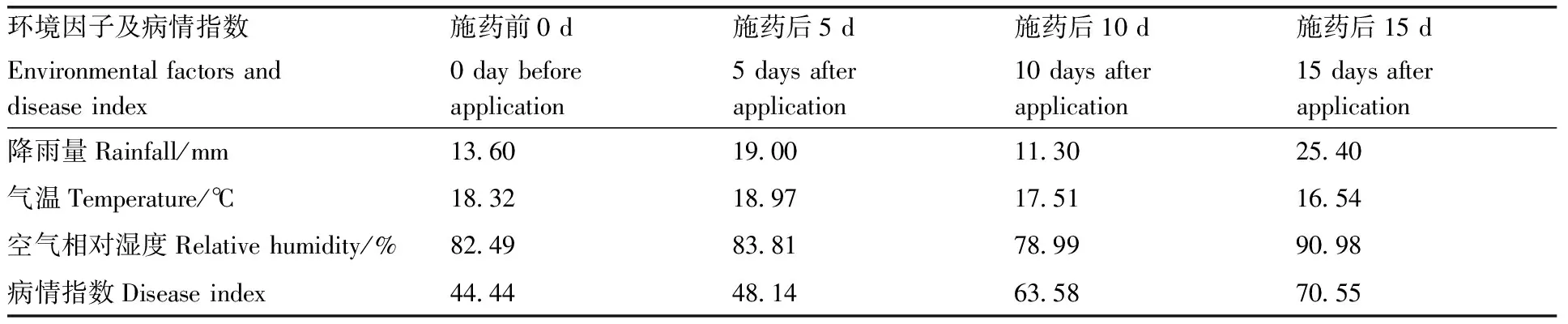
表3 环境因子与病情指数
本文对叶际真菌和细菌top50的菌属进行Spearman相关分析,结果显示,在真菌属水平上,取样时间与粉褶蕈属(Entoloma)、青霉菌属(Penicillium)、拟孔菌属(Fomitiporia)、红酵母属(Rhodotorula)、木霉属(Trichoderma)、踝节菌属(Talaromyces)、亚隔孢壳属(Didymella)、枝孢霉属(Cladosporium)的相对丰度呈显著正相关;与浪梗霉属(Polythrincium)的相对丰度呈极显著正相关;与谷菌根菌属(Archaeorhizomyces)的相对丰度呈显著负相关。温度与粉褶蕈属(Entoloma)、青霉菌属(Penicillium)、拟孔菌属(Fomitiporia)、红酵母属(Rhodotorula)、木霉属(Trichoderma)、踝节菌属(Talaromyces)、枝孢霉属(Cladosporium)的相对丰度呈显著负相关,与浪梗霉属(Polythrincium)的相对丰度呈极显著负相关。相对湿度和降雨量与拟孔菌属(Fomitiporia)的相对丰度呈显著正相关。病情指数与Plectosphaerella、毕赤酵母菌属(Pichia)、黑团孢属(Periconia)的相对丰度呈显著负相关;与尾孢菌属(Cercospora)、镰刀菌属(Fusarium)、曲霉菌属(Aspergillus)、Boeremia的相对丰度呈极显著负相关,与链格孢属(Alternaria)呈极显著正相关(图3-A)。
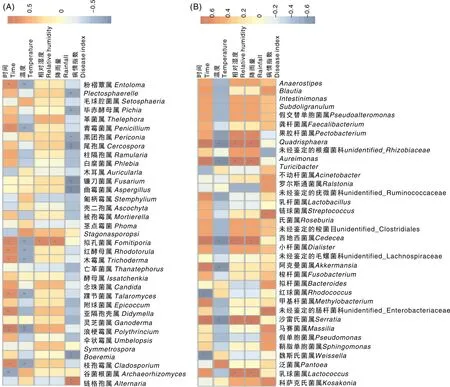
*, P<0.05; **, P<0.01.图3 腈菌唑处理后叶际真菌(A)和细菌(B)群落的Spearman相关分析热图Fig.3 Heat map of Spearman correlation analysis based on phyllosphere fungal (A) and bacterial (B) communities after myclobutanil treatment
在细菌属水平上,取样时间与红球菌属(Rhodococcus)、魏斯氏菌属(Weissella)的相对丰度呈显著负相关;与Quadrishaera、Aureimonas、西地西菌属(Cedecea)、阿克曼菌属(Akkermansia)、沙雷菌属(Serratia)的相对丰度呈显著正相关。温度与Quadrishaera、Aureimonas、西地西菌属(Cedecea)、阿克曼菌属(Akkermansia)、沙雷菌属(Serratia)的相对丰度呈显著负相关。相对湿度和降雨量与Quadrishaera、金色单胞菌属(Aureimonas)、西地西菌属(Cedecea)、沙雷菌属(Serratia)、泛菌属(Pantoea)的相对丰度均呈显著正相关。病情指数与链球菌属(Streptococcus)、未经鉴定的肠杆菌科(undentified-Enterobactericeae)的相对丰度呈显著正相关关系(图3-B)。
2.2 腈菌唑对感赤星病与健康烟叶叶际微生物代谢功能的差异
本文采用Biolog-ECO微孔板对施药前后烟叶样品进行代谢功能分析,Biolog ECO微孔板中含有羧酸类、氨基酸、糖类、双亲化合物、聚合物和胺/氨基化合物共31种碳源。施药前,感病烟叶叶际微生物对α-丁酮酸、L-苏氨酸的代谢较弱;对葡萄糖-1-磷酸盐、4-羟基苯甲酸、α-环式糊精、D,L-α-甘油的代谢程度一般,对其他碳源代谢均可高效代谢,尤其是对α-D-乳糖、D-甘露醇、D-半乳糖酸内酯、γ-羟基丁酸、L-天冬酰胺酸、吐温40、吐温80。健康烟叶叶际微生物对α-丁酮酸的代谢较弱,对2-羟基苯甲酸代谢程度一般,对其他碳源代谢均可高效代谢。施药5 d时,感病烟叶叶际微生物对α-丁酮酸代谢程度增加,对D-葡糖胺酸、α-环式糊精代谢程度保持不变,其余碳源代谢程度均降低;健康烟叶叶际微生物对所有碳源代谢程度均降低。施药10 d时,相较于施药前和施药5 d时,感病烟叶叶际微生物对α-丁酮酸代谢程度增加,对其余碳源代谢均降低;健康烟叶相较于施药前,α-丁酮酸代谢程度增加,其余碳源代谢程度均降低,相较于施药5 d时,α-D-乳糖、D-半乳糖醛酸、苯乙基胺等11种碳源代谢程度保持不变,D-纤维二糖、D-木糖、丙酮酸甲酯等14种碳源代谢程度增加,对其余碳源代谢程度均降低。施药15 d时,相较于施药前,感病烟叶叶际微生物对α-丁酮酸代谢程度增加,其余碳源代谢程度均降低,相较于施药10 d时,D-纤维二糖、α-D-乳糖、D-葡糖胺酸等8种碳源代谢程度均增加,2-羟基苯甲酸、4-羟基苯甲酸、α-丁酮酸代谢程度均降低,其余碳源代谢程度均增加;健康烟叶相较于施药前,所有碳源代谢程度均降低,相较于施药10 d时,苯乙基胺、L-精氨酸、L-苯基丙氨酸等6种碳源代谢程度增加,D-木糖、L-丝氨酸、D-半乳糖醛酸等10种碳源代谢程度不变,其余碳源代谢程度均降低(图4)。
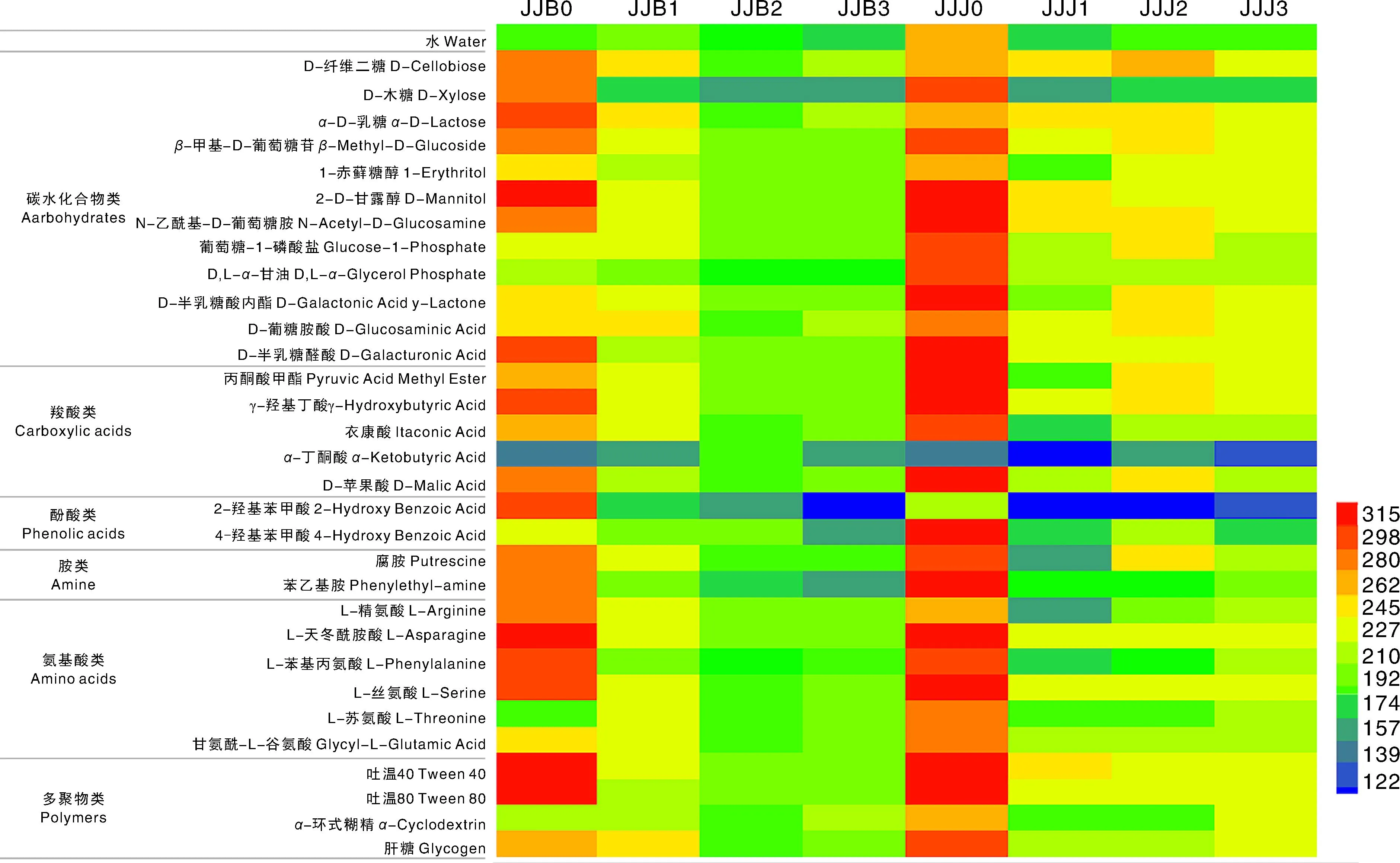
颜色值分别代表微生物代谢碳源的程度,122~<192为代谢程度低,192~262为代谢程度一般,262以上为代谢程度高。The color values represent the degree of the microbes metabolizing carbon sources. The value 122-<192, 192-262 and >262 represent the leaf microbes metabolized in Biolog ECO microplate poorly, moderately, effectively, respectively.图4 烟叶叶际微生物代谢功能聚类热图Fig.4 Cluster heat map of phyllosphere microbial metabolic function in tobacco leaves
3 讨论
叶际微生物在维持生态系统平衡和叶片健康方面均具有不可忽视的作用[26-27]。已有研究发现烟草赤星病危害期其叶际优势真菌为链格孢属,优势细菌有泛菌属和假单胞菌属[11]。本文采用Illumina高通量测序技术,对应用腈菌唑前后感赤星病烟叶和健康烟叶叶际真菌和细菌群落结构与多样性进行分析,发现施药前后感病和健康烟叶叶际优势真菌菌群均为子囊菌门,其次为担子菌门;优势细菌菌群为变形菌门,其次为厚壁菌门。感赤星病烟叶和健康烟叶施药前后的真菌有链格孢属、枝孢霉属、亚隔孢壳属、Boeremia、Symmetrospora等菌属,优势菌属为链格孢属。施药前后感赤星病烟叶与健康烟叶的主要细菌有泛菌属、假单胞菌属、鞘脂单胞菌属等,其中优势菌属为泛菌属。这与前人对于感赤星病烟叶采烤后优势真菌菌群和感赤星病烟叶成熟期优势真菌与细菌菌群的研究结果一致[12,28]。进一步证实链格孢属为赤星病烟叶叶际优势真菌,泛菌属、假单胞菌属为叶际优势细菌。本文研究发现,感病烟叶的真菌链格孢属相对丰度远高于健康烟叶,说明烟草感赤星病时,链格孢属占据主导地位;其余真菌(枝孢霉属、Symmetrospora、亚隔孢壳属等)和细菌(泛菌属、假单胞菌属等),在烟草亚隔孢叶斑病和烟叶霉烂病危害期均出现[29-30],为此,推测这些菌属为烟叶上的习居菌。
药剂的使用会改变叶际微生物群落结构[31]。研究发现,施用菌核净会使烟叶叶际细菌中鞘脂单胞菌属、黄色杆菌属和沙雷氏菌属的相对丰度降低[14];高效氯氰菊酯在推荐使用剂量范围内使用会引起植物叶际真菌生物量的降低和细菌生物量的增加[32];白僵菌的施用也会使水稻叶际微生物群落结构发生较大改变[33]。本文发现,施药后感病烟叶和健康烟叶叶际病原真菌链格孢属的相对丰度均降低,说明腈菌唑能有效防治烟草赤星病的发生。本文通过Spearman相关分析发现,病情指数与链格孢属呈极显著正相关,表明病情指数越大,链格孢属的相对丰度越高,而病情指数与尾孢菌属、镰孢菌属、曲霉属、Symmetrospora的相对丰度呈极显著负相关,推测这些菌属与链格孢属之间存在潜在的竞争作用,但其具体机理及这些真菌是否参与侵染有待进一步研究。除靶标病原菌外,腈菌唑会影响烟叶包括附球菌属、枝孢霉属、Symmetrospora、亚隔孢壳属等在内的多种病原真菌,同时也会影响包括假单胞属、泛菌属在内的多种非致病细菌菌群结构的变化,这与前人研究结果类似[12]。此外,腈菌唑作为三唑类杀菌剂的典型代表,已在多种作物上登记,本文研究结果也进一步验证了腈菌唑的广谱性。
高多样性和丰度水平的叶际微生物群落可能有助于提高叶际环境的功能冗余,维持宿主植物的健康水平[34]。本文利用Shannon指数和ACE指数用作表征微生物群落多样性和衡量微生物群落丰富度,研究发现,感赤星病烟叶的叶际真菌群落多样性指数和丰富度指数均低于健康烟叶,且存在显著性差异,结果与烟草靶斑病[35]等真菌性病害的相关叶际微生物多样性与丰富度研究结果一致,推测真菌性病害的发生均会降低叶际菌群的丰富度与多样性,也进一步证明菌群高丰富度和多样性更有利于维持宿主植物的健康水平。腈菌唑应用后感病烟叶真菌的多样性先升高,后降低,而后升高;丰富度一直保持上升趋势,而细菌多样性和丰富度均先降低,而后升高。健康烟叶真菌多样性和丰富度一直保持上升趋势,而细菌多样性先降低,而后升高,丰富度则一直保持上升趋势。说明药剂处理显著改变了叶际微生物群落多样性与丰富度,本文研究结果与前人研究发现施用拮抗菌群、杀菌剂会改变感野火病、赤星病烟叶叶际微生物群落结构研究结果类似[15,36]。
叶际微生物群落对碳源代谢活性的强弱能反映代谢功能的强弱。本文利用Biolog-ECO技术研究分析了施药前后感赤星病烟叶与健康烟叶对常见31种碳源的碳源利用差异,体现烟叶叶际微生物整体活性和微生物群落生态功能,发现施药前感病烟叶对α-丁酮酸、L-苏氨酸利用较弱外,还有少数几个碳源利用程度较低。而健康烟叶除了对α-丁酮酸的代谢较弱,对2-羟基苯甲酸代谢程度一般,对其他碳源代谢均可高效代谢。说明感病烟叶叶际微生物对于碳源的代谢能力低于健康烟叶,健康烟叶的生物活性高于感病烟叶。腈菌唑处理后,相较于施药前,感病烟叶与健康烟叶的碳源利用降低,随着时间的增加,对碳源的利用又逐渐升高,健康烟叶恢复对碳源利用能力比感病烟叶强。表明腈菌唑能快速降低感赤星病烟叶与健康烟叶叶际微生物的代谢活性,很大程度上改善了烟叶叶际微环境,但对于健康烟叶的活性影响比感病烟叶小,随着施药时间的增加,代谢活性缓慢增加,表明腈菌唑在烟叶上的持效期有限,其用于赤星病防控时可适当增加药剂施用次数。
4 结论
赤星病发生期,感病烟叶和健康烟叶叶际优势菌属均为链格孢属、科萨克氏菌属,但赤星病烟叶中链格孢属、泛菌属相对丰度高于健康烟叶,而真菌和细菌的多样性指数和丰富度指数均低于健康烟叶。腈菌唑施用后降低了赤星病烟叶和健康烟叶叶际主要优势菌属链格孢属的相对丰度、增加了科萨克氏菌属相对丰度;感赤星病烟叶叶际真菌多样性和丰富度指数先降低,而后增加,健康烟叶中多样性和丰富度指数均增加。施药前赤星病烟叶对碳源的代谢能力低于健康烟叶,健康烟叶能高效代谢几乎所有碳源。施药后二者碳源代谢程度均降低,随着时间的延长,碳源代谢程度逐渐恢复。
