香榧藻斑病病原的分离鉴定及防治药剂筛选*
2024-02-01刘艾涛叶碧欢陈友吾宋其岩李海波沈建军
刘艾涛 叶碧欢 陈友吾 宋其岩 李海波 沈建军 张 昕
(1. 浙江农林大学林业与生物技术学院 杭州 311300; 2. 浙江省林业科学研究院森林保护所 杭州 310023)
藻斑病是一种世界范围内常见的木本植物叶部病害,在国外已报道可危害黑莓(Rubus fruticosus)(Browneet al.,2019)、榕树(Ficus benghalensis)(Hanet al.,2011)、橡胶树(Hevea brasiliensis)(Pitalokaet al.,2015)、番石榴(Psidium guajava)(Sunpapaoet al.,2016a)和龙贡果(Lansium parasiticum)(Sunpapaoet al.,2016b),在我国主要发生于山茶(Camellia japonica)(何美仙等,2017)、肉桂(Cinnamomum cassia)(郑宝荣等,2004)和夏橙(Valenciasp.)(王大平等,2005)等植物。藻斑病的病原为寄生藻,引起不同寄主植物藻斑病的病原藻有所不同,头孢藻(Cephaleurossp.)、虚幻球藻(Apatococcus lobatus)、小球藻(Chlorellasp.)等藻类均被报道可引起藻斑病发生,这些藻在叶片上表面定殖,掠夺植物营养,导致寄主植株长势衰弱(Chanthapatchotet al.,2019;Vasconceloset al.,2016)。
香榧(Torreya grandiscv. ‘Merrillii’)是我国特有的珍稀经济树种,野生香榧主要分布于浙江省会稽山区一带(戴文圣等,2006)。香榧具有丰富的营养价值、较高的经济价值和药用价值(王鸿等,2007),是浙江多地林业经济支柱产业,为此浙江省还提出“香榧南扩”的发展战略(吴连海等,2013)。随着香榧产业化种植的不断发展以及种植面积的逐渐扩大,香榧病虫害问题日益严峻,一些以往零星发生、危害较轻的病害,如香榧藻斑病,近年来在不同新老产区普遍发生,且呈逐年加重趋势,严重地区发病率达70%以上。病原藻主要侵染香榧叶片和嫩枝,在受害部位表面繁殖后产生粉末状堆积的藻斑,发病重时藻斑连片,严重影响植株的光合作用和新芽的抽出,常造成香榧落花落果和减产,给榧农带来重大经济损失。徐志宏等(2005)对香榧藻斑病研究认为,其病原为一种小球藻,但对病原藻的形态特征和鉴定未给出明确描述和数据。叶晓明等(2019)从浙江和安徽两省的香榧藻斑病病叶中分离获得病原藻,经细胞形态学结合藻株的18S rDNA 以及ITS 序列分析,将所得藻株鉴定为栅藻科(Scenedesmaceae)的Asterarcys quadricellulare。笔者前期研究发现,香榧藻斑病由多种藻混合发生引起,且病原藻形态与已有报道有所不同。本研究在浙江省香榧主产区广泛采集病叶样本,拟通过形态学与分子生物学相结合的手段探明引起香榧藻斑病的病原种类,了解其生物学特性,并进行室内和室外药剂筛选研究,以期为香榧藻斑病防治提供科学依据。
1 材料与方法
1.1 试验材料
1.1.1 病叶采集 2021年4月—2022年11月,多次于浙江省遂昌三仁乡、诸暨东白湖镇和桐庐旧县3个香榧主产区采集新鲜发病的香榧藻斑病病叶,带回室内用于病原藻分离。
1.1.2 主要试剂 用于藻类培养的蓝绿藻培养基BG-11 购自青岛海博生物技术有限公司,扎鲁克培养基(Zarrouk)、SE 培养基(Selenite Enrichment)和BBM培养基(Bold’s Basal Medium)购自北京谱蓝生物科技有限公司;供试农药分别为20%噻菌铜剂悬浮剂(浙江龙湾化工)、33.5%喹啉铜悬浮剂(浙江台州顺毅股份有限公司)、12%松脂酸铜悬浮剂(广东植物龙生物技术有限公司)、80%波尔多液(通州正大农药化工)和45%石硫合剂(河北双吉化工);藻株DNA提取试剂盒TIANamp Genomic DNA Kit 购自北京天根生化科技有限公司。其他试剂均为国产分析纯。
1.2 试验方法
1.2.1 病原藻分离 按照说明书方法,称取1.70 g BG-11 固体粉末溶于1 000 mL 无菌水中,混匀灭菌后制得BG-11 液体培养基。取新鲜发病的香榧藻斑病病叶,用无菌刮刀刮取适量叶面绿藻混合物置于含100 mL 液体培养基的三角瓶中,25 ℃恒温培养箱内培养。其间设置光照/黑暗光周期为12 h/12 h,光照强度6 000 lx。2 周后,取藻液利用平板划线法在含琼脂粉1.5%(m/v)的BG-11 固体培养基上分离藻株。通过肉眼与显微镜观察相结合,对平板长出的藻的菌落进行分类并用无菌接种环蘸取菌落边缘的绿藻在新平板上进行多次分离纯化,直至获得单一绿藻藻株。
1.2.2 致病性测定 取生长状况相近的50 cm 高香榧盆栽幼苗,幼苗叶片表面用75%酒精消毒后,取上述获得的、用无菌水配制成浓度为4×107~6×107个∙mL−1的各藻悬液,采用喷雾法按每株60 mL 均匀喷洒至幼苗叶片,喷洒后香榧幼苗置于25 ℃人工气候培养箱中培养,其间保持相对湿度80%、光照/黑暗光周期为12/12 h、光照强度6 000 lx。试验持续30 天,其间观察记录叶片发病情况,并从发病叶片上重复藻种分离过程,验证其同一性。试验每藻种5 次重复,以喷雾无菌水的处理为对照。
1.2.3 藻株分子鉴定 刮取培养基表面纯培养藻株采用TIANamp Genomic DNA Kit 试剂盒提取基因组DNA,以其为模板,采用藻通用引物对EAF3/055R(EAF3:TCGACAATCTGGTTGATCCTGCCAG;055R:CT CCTTGGTCCGTGTTTCAAGACGGG)扩增香榧绿藻的SSU-ITS 片段(Marinet al.,2003),目标片段经测序后在NCBI 数据库中进行BLAST 比对,选取相似度较高的序列利用MEGA 6.0 软件采用邻近法(neighborjoining,NJ)构建系统进化树。模型为K2P(Kimura 2 parameter),Bootstrap 置信值估算重复数设为1 000 次。本试验PCR 扩增的反应体系为20 μL:模板DNA 3 μL(20 ng∙L−1), 2×TSINGK Master Mix 10 μL,10 μmol∙L−1引物对各 1.5 μL,ddH2O 4 μL。扩增程序为:94 ℃预变性5 min 后;94 ℃变性1 min,60 ℃退火1 min,72 ℃延伸2 min,共40 个循环;于72 ℃补平10 min,终止温度4 ℃。
1.2.4 藻株培养特性 取对数期的各藻株藻液(OD680=0.2)100 μL 分别加入盛有10 mL BG-11 液体培养基的试管中,于不同初始pH(3.0、7.0 和11.0)、温度(20 ℃、25 ℃、30 ℃)和光照(无光照、光照/黑暗=12 h/12 h 和光照24 h)条件下培养。每隔4 天测定680 nm 波长下藻液的吸光值(OD680)(Caiet al.,2012),比较不同藻株对环境的适应性。采用同样方法将藻液接入不同培养基(BG-11、Zarrouk、SE 和BBM),研究各藻株对营养基质的需求差异。本试验其他培养条件同1.2.1,每处理3 次重复,以添加无菌水为对照。藻株的相对生长率[Y(%)]为:Y(%)=(ODt−ODCK)/ODCK×100,其中ODt表示t时间时藻液的吸光值,ODCK表示0 时间时藻液的吸光值。
1.2.5 病原离体抑制药剂筛选 以无菌刮取的香榧叶片藻斑为试验对象,测定20%噻菌铜剂悬浮剂、33.5%喹啉铜悬浮剂、12%松脂酸铜悬浮剂、80%波尔多液和45%石硫合剂5 种化学药剂对其生长的毒性效应。具体步骤如下:将各供试药剂与BG-11 配制成5 种不同体积分数梯度(1∶26 667、1∶8 000、1∶2 667、1∶800 和1∶267)(V/V)的含药培养液,每种药剂每种梯度7 mL 分装试管。向各处理试管中接种培养至OD680=0.2 的藻液3 mL 后将试管置于光照培养箱中同1.2.1 所述条件培养,16 天后测定680 nm波长下藻液的吸光值。试验分别设置添加等量无菌水替代藻液的阴性对照(CK1)和含有藻液但不含药剂的阳性对照(CK2),每处理3 个平行。药剂对病原藻的抑制率(%)=[ODCK2−(ODT−ODCK1)]/ODCK2×100,其中ODT表示药剂处理组的吸光值,ODCK1表示阴性对照的吸光值,ODCK2表示阳性对照的吸光值。
1.2.6 林间防治试验 2021年10月,在浙江省诸暨东白湖镇王坑村香榧林(120°26′32″E, 29°35′21″N)选取藻斑病发生较为严重的20~30年生香榧人工林开展林间防治试验。分别取室内测定效果较好的12%松脂酸铜悬浮剂和33.5%喹啉铜悬浮剂1 000 倍液,采用喷雾法均匀喷洒病叶和枝条至药液微微下滴为止。该试验每处理10 株香榧,3 次重复,以喷雾清水为对照(CK)。喷雾前和喷雾后30 天,每株香榧从东南西北4 个方向统计发病情况。藻斑病的发病情况分级如下:0 级,无症状;1 级,只有个别枝叶发病;3 级,发病枝叶数<1/3;5 级,1/3≤发病枝叶片数<1/2;7 级,1/2≤发病枝叶数<2/3;9 级,发病枝叶数≥3/4。病害的防治效果计算公式如下:
式中:CK0表示空白对照区施药前的病情指数;CK1表示空白对照区施药后的病情指数;T0表示药剂处理区施药前的病情指数;T1表示药剂处理区施药后的病情指数。
1.3 数据分析
利用Excel 软件整理数据;运用SPSS19.0 软件对数据进行方差分析、毒力回归方程和有效中浓度(median effective concentration,EC50)计算;采用Origin 2016 软件绘图。
2 结果与分析
2.1 藻株的分离和形态学观察
分离香榧藻斑病病原时发现,该病害由多种藻混合侵染引起。数十次分离中均可在同一病叶上获得多种形态差异较大的藻株,选取能获得纯培养、外观形态稳定且有明显差异的3 种类型藻株,编号后保存于浙江省林业科学研究院森林保护所。从3 种藻群中分别选择典型藻株XFLZ928.6、XFLZ928.8 和XFLZ928.15.1 用于后续试验。
形态学观察发现,藻株XFLZ928.6 为单细胞,多以2 个或多个细胞整齐排列和聚集在同一平面,细胞壁表面平滑,胞体呈椭圆形,长5.35~6.51 μm,宽2.92~3.81 μm,胞体一端有时长有体刺,色素体周生(图1A)。藻株XFLZ928.8 的单个细胞常首尾相接形成不分枝的丝状体,其细胞圆柱状,单细胞长9.54~13.50 μm,宽9.40 μm,细胞壁薄,细胞内可见清晰淀粉粒和色素体。色素体多沿细胞边缘分布,常覆盖整个细胞长度(图1B)。藻株XFLZ928.15.1 为单细胞,常离散分布,细胞圆柱状,两端钝圆,长4.87~7.13 μm,宽2.34~3.00 μm,长宽比1.87~2.88,色素体多聚于中间或一侧,不充满细胞(图1C)。根据形态学观察, 初步认定藻株XFLZ928.6、 XFLZ928.8 和XFLZ928.15.1 分别属于链带藻属(Desmodesmus)、克里藻属 (Klebsormidium)和Tritostichococcus属绿藻。

图1 藻株XFLZ928.6(A)、XFLZ928.8(B)和XFLZ928.15.1(C)的形态Fig. 1 Morphology of algae XFLZ928.6(A), XFLZ928.8(B) and XFLZ928.15.1(C)a:体刺 Spines ;b:色素体 Chloroplasts; c:淀粉粒 Starch granules
2.2 致病性
将上述所得绿藻分别制成藻悬液接种香榧后第14 天观察发现,经3 个藻株处理的香榧幼苗叶片表面均开始出现零星、浅绿色藻点,后随藻的繁殖,藻点面积逐渐扩大并形成呈粉末状堆积的不规则藻苔,至23 天时藻苔连接成片,几乎覆盖整个叶片表面(图2B、图2C、图2D),与田间发病症状一致(图2E)。而接种清水对照的幼苗生长正常,叶片上未有藻斑发生(图2A)。对发病叶片再次分离纯化和镜检发现,其形态与接种的病原藻形态相同,最终确定试验中分离获得的3 种绿藻皆为香榧藻斑病的病原藻。

图2 3 种藻类的致病性测定Fig. 2 Pathogenicity assay of three algae
2.3 藻株的系统发育分析
为进一步明确所获3 种绿藻的系统发育地位,对藻株的SSU-ITS 区间序列进行分析。绿藻XFLZ928.6、XFLZ928.8 和XFLZ928.15.1 经PCR 扩增后均出现清晰的SSU-ITS 区间条带(图3),分别获得2 288、2 491和1 546 bp 的序列片段(GenBank 登录号分别为ON040656、ON040657 和ON040658)。经Blast 比对发现,藻株XFLZ928.6 与链带藻属的同源性最高,与D. armatus的相似性大于97%,选取序列相似性较高的藻株构建系统发育树发现,本研究分离获得的XFLZ928.6 与之聚为一支。结合形态学特征,本研究将XFLZ928.6 最终鉴定为链带藻(D. armatus)。藻株XFLZ928.8 与克里藻属的绿藻有很高同源性,与软克里藻(K. flaccidum)的相似性大于99%,系统发育树也显示该绿藻与软克里藻聚为一支,支持率为100%,结合形态特征,XFLZ928.8 最终被鉴定为软克里藻。藻株XFLZ928.15.1 与共球藻纲(Trebouxiophyceae)的Tritostichococcussp.、 裂 丝 藻 属 (Stichococcus)和Desmococcussp.均具有较高同源性,其中与T. corticulus的相似性最高,系统发育树也显示二者聚为一支,支持率为100%,结合形态特征可判定XFLZ928.15.1 为T. corticulus(图4)。

图3 藻株XFLZ928.6、XFLZ928.8 和 XFLZ928.15.1 的SSU-ITS 电泳分析Fig. 3 Electrophoresis analysis of SSU-ITS of algae XFLZ928.6,XFLZ928.8 and XFLZ928.15.1
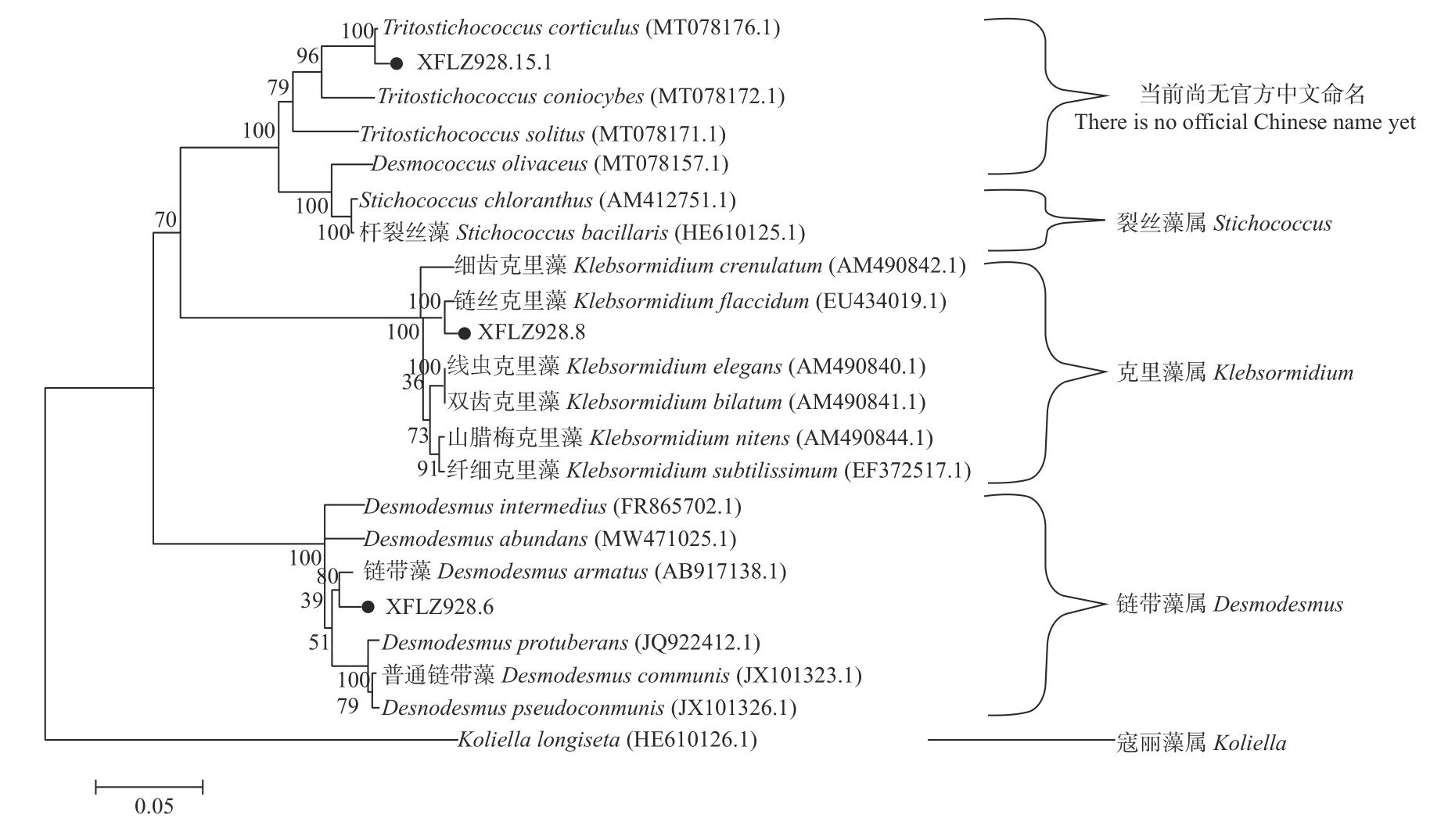
图4 基于SSU-ITS 基因序列构建的3 种绿藻系统发育树Fig. 4 Phylogenetic tree of three green algae based on SSU-ITS gene sequence
2.4 病原藻的培养特性
2.4.1 光照对病原藻生长的影响 试验比较不同光照条件对3 种供试藻株的影响,结果见图5。光照对藻的生长影响较大,且3 种藻株对光的响应表现出一致性。在完全黑暗的情况下,藻不能够正常生长,且随着时间延长生长情况日益恶化。而全天候(24 h)的光照也非藻种最适的光照条件,本试验与自然条件类似的12 h 光照/12 h 黑暗光周期更适宜供试的藻株XFLZ928.6、XFLZ928.8 和XFLZ928.15.1生长。

图5 不同光照条件对3 种香榧绿藻生长的影响Fig. 5 Effects of light on the growth of three green algae of T.grandis cv. ‘Merrillii’
2.4.2 pH 对病原藻生长的影响 由图6 可知,3 种藻株在初始pH 为3 时生长受到严重抑制,在初始pH为11 的条件下生长状况最优,说明初始碱性环境比中性与酸性环境更适宜藻的生长。藻株XFLZ928.6和XFLZ928.8 在初始pH 为11 时,培养4~16 天增长较快,之后生长趋于平缓,而藻株XFLZ928.15.1 在初始pH 为11 时整个试验周期生长率一直稳步增长。

图6 不同pH 条件对3 种香榧绿藻生长的影响Fig. 6 Effects of different initial pH levels on the growth of three green algae of T.grandis cv. ‘Merrillii’
2.4.3 温度对病原藻生长的影响 温度是影响藻类生长的重要环境因素。由图7 可知,整个试验周期内,3 种株藻在20、25 和30 ℃条件下的生长趋势相似,随着培养时间延长,藻株生长率均先稳步上升然后趋于平缓,其最适宜生长温度为25 ℃,在该温度条件下,各供试绿藻均表现出良好生长状态,相同培养时间的相对增长率最高,其次为30℃,20 ℃条件下生长率最低。

图7 不同温度条件对3 种香榧绿藻生长的影响Fig. 7 Effects of temperature on the growth of three green algaes of T.grandis cv. ‘Merrillii’
2.4.4 不同培养基对病原藻生长的影响 将藻接入各供试培养基后测定藻的相对生长率发现,BBM 和Zarrouk 培养基不适合香榧绿藻生长,3 种藻株在BBM 和Zarrouk 培养基中均生长较差,藻株XFLZ 928.6 和XFLZ928.15.1 在培养16 天、藻株XFLZ928.8在培养12 天时出现峰值,随后生长率逐渐下降。而在BG-11 培养基中,3 种藻株均良好生长,在试验期间生长率随时间变化逐渐增加,在24 天时仍有增长趋势,且藻的相对生长率始终处于最高位,远远高于位居第二的SE 培养基,该趋势对于藻株XFLZ928.8尤其明显(图8)。

图8 不同培养基对3 种香榧绿藻生长的影响Fig. 8 Effects of culture medium on the growth of three green algae of T.grandis cv. ‘Merrillii’
2.5 室内药剂筛选及林间防治
为防治香榧藻斑病,选取当前市面上常用的5 种化学药剂进行室内灭藻毒力测定试验,结果见表1。12%松脂酸铜悬浮剂的灭藻效果最佳,16 天时EC50最小,为8.33 mg·L−1,显著低于其他4 种药剂(P<0.05),而45%石硫合剂的灭藻效果最差,EC50最大,为2 778.18 mg·L−1。33.5%喹啉铜悬浮剂的灭藻效果仅次于12%松脂酸铜悬浮剂,但优于其余3 种药剂,80%波尔多液与20%噻菌铜剂悬浮剂的灭藻效果相近,EC50差异不显著。

表1 5 种化学药剂的室内毒力测定①Tab. 1 Laboratory evaluation of the toxicity of five algicides
将室内毒力试验效果较好的12%松脂酸铜悬浮剂和33.5%喹啉铜悬浮剂用于香榧林间防治,结果(表2)表明,随着时间推移,未进行药剂防治的对照林区藻斑病发生逐步加重,病害的病情指数由63.33升至67.20,而供试2 种药剂的林区施药30 天后均有较好防治效果(大于60%),其中12%松脂酸铜悬浮剂的防治效果最佳,为78.11%,高于33.5%喹啉铜悬浮剂的71.62%,说明前者的灭藻效果优于后者,与室内毒力测定结果一致。

表2 2 种不同药剂的林间防治效果Tab. 2 Field control effect of two different algicides
3 讨论
3.1 香榧藻斑病症状的特殊性
藻斑病是热带和亚热带地区木本阔叶植物上的多发病害,尽管已有报道中不同寄主其病原藻可能存在差异,但寄生藻在叶片内部侵染造成植物组织坏死,导致染病叶片或茎秆表面形成近圆形、凸起的黄褐色或红褐色、边缘不整齐病斑(Sunpapaoet al.,2016a)为该病害的共有特征。本研究中香榧藻斑病的症状与之截然不同,由于香榧藻斑病的病原藻为气生藻而非寄生藻,不能侵入植物细胞内部,因此仅在香榧叶片和嫩枝的上表面形成浅绿色至灰绿色不规则堆积的粉末状藻斑,病重时藻斑可覆盖整个叶片,严重影响树木的光合作用,导致香榧落花落果并最终减产。发病症状及病原致病机制不同,可能是香榧的病原藻与已有报道阔叶树的寄生藻不同的原因之一。
3.2 3 种新香榧病原藻群
分离香榧藻斑病的病原发现,该病害由多种不同形态的藻群混合发生引起。叶晓明等(2019)通过形态学以及藻株的18S rDNA 和ITS 序列分析,将形态和大小有所不同的藻株A 和藻株B 均鉴定为A. quadricellulare。本研究从香榧藻斑病的感病叶片上获得3 种形态差异较大的藻株类群,与叶晓明等(2019)描述的病原藻均有明显不同。鉴于藻的形态受环境因素的影响较大,采用分子生物学手段可更客观反映藻株的系统发育地位。Pröschold 等(2020)在研究裂丝藻属的SSU 及ITS 序列时发现,SSU+ITS的系统发育分析方法在藻类鉴定中的可靠性高于ITS 和rbcl 等的单独分析鉴别。因此,本研究在形态鉴定基础上,在分子层面通过SSU-ITS 测序对3 种香榧绿藻进行系统发育分析,最终将3 种香榧绿藻类群分别确定为栅藻科的链带藻以及丝藻科(Ulotrichaceae)的软克里藻和T. corticulus。目前,国内外对链带藻属藻类的研究主要集中在生物资源利用和环境修复方面(Liet al.,2015;Chandelet al.,2022;Öztürk,2019),迄今为止未见该属藻类侵染植物的报道。本研究中,藻株XFLZ928.6 侵染香榧是链带藻属藻种引起植物病害属首次发现。由于Tritostichococcus于2020年才被作为新属划分,且缺乏相关的基因序列数据,当前国内外缺乏关于Tritostichococcussp.引起植物病害的报道,本研究则证实T. corticulus对香榧的致病作用。软克里藻是重要的生物土壤结皮微生物,对该属藻的已有研究多集中在生态效应和物种多样性方面(Glaseret al.,2017;Karstenet al.,2016;Linet al.,2012;Ryšaneket al.,2016),除Lin 等(2012)首次从中国台湾柏树林的阔叶植物叶片上分离获得软克里藻外,未见其他相关研究证实克里藻属的绿藻引起植物病害。本研究中,藻株XFLZ928.8 侵染香榧也是软克里藻危害针叶树的首次报道。
综上,本研究从香榧藻斑病病叶上分离获得的3种绿藻与已有报道皆不相同,其中,链带藻和T.corticulus为侵染植物致病的首次报道,而软克里藻为侵染针叶树的首次发现。本研究不仅为探明香榧藻斑病的病原提供新的有效信息,同时也拓展了对植物藻斑病病原的认知。
3.3 藻株的生长特性
本研究从光照、温度、初始pH 和营养物质需求角度对链带藻、软克里藻和T. corticulus 的生长特性进行研究,结果表明,这3 种香榧绿藻具有类似的生态特性,相近的生态习性可能是3 种藻在香榧上引发混合侵染的主要原因。藻对环境的适应性是长期进化和选择的结果,具有种属的特异性。本研究中分离的3 种藻均在初始pH 为11 时生长良好,与叶晓明等(2019)发现香榧绿藻A. quadricellulare适合在碱性条件下生长的研究结果一致。然而就温度适应性而言,本研究3 种藻株与已报道的病原虚幻球藻(Apatococcus lobatus)(20~25 ℃)(王大平等, 2005)、铜绿微囊藻(Microcystis aeruginosa)(33 ℃)(王霞等,2005)、小球藻(Chlorella marina)(37.5 ℃)(欧阳峥嵘等, 2010)、 汉斯冠盘藻(Stephanodiscus hantzschii)(3~15 ℃)、小环藻(Cyclotellasp.)(15 ℃)和斜生栅藻(Scenedesmus obliquus)(30 ℃)(许珍,2017)均不相同,表明本研究所分离藻株与以上可能均不同。而香榧藻斑病在早春的5—7月发生最严重,除了梅雨季节的雨水因素外,藻株对最适生长温度为25 ℃,也是病害在此时段发生较为严重的一个重要原因。
3.4 藻斑病的药剂筛选
藻斑病的严重发生已成为困扰我国长江以南地区香榧等特色经济林产业发展的瓶颈问题,过去在生产实践中常用的石硫合剂(吕军美等,2015),可能是因长期施用导致其产生抗药性,防治效果逐渐变差,因为藻对铜离子具有较强的敏感性,铜制剂被当作常用的除藻剂。本研究测定当前市面上5 种药剂对香榧藻斑病病原的毒力和林间防治效果,发现有机铜制剂12%松脂酸铜悬浮剂和33.5%喹啉铜悬浮剂的灭藻效果最佳,80%波尔多液次于前二者,效果与20%噻菌铜悬浮剂相近,而生产实践中常用于防治藻斑病的石硫合剂效果最差。此外,与波尔多液等无机铜制剂混配性差、容易污染树木叶片和果实、产生药害的特性相比,松脂酸铜等有机铜制剂高效安全、不易产生药害以及对环境友好的特点令其更具选择优势。本研究所得试验结果为香榧藻斑病的高效防控提供了科学依据。
4 结论
本研究在香榧病叶上新发现3 种混生的病原绿藻类群,经鉴定分别属于链带藻、软克里藻和T. corticulus,3 种类群的藻株具有类似的环境适应性,均适宜在温度25 ℃、初始pH11、12 h 光照/12 h 黑暗光周期的BG-11 培养基中生长。结合室内离体抑制试验和林间实际防治效果,筛选出防治香榧藻斑病的优良药剂12%松脂酸铜和33.5%喹啉铜悬浮剂。研究结果拓宽了当前对香榧藻斑病病原的认知,也为病害的防治提供了理论依据和参考。
