初植和补植阔叶树对红壤丘陵区湿地松养分获取和转运的影响*
2024-02-01夏成康吴高洋王晟楠陈伏生
夏成康 林 勇 兰 勇 吴高洋 王晟楠 陈伏生
(1. 江西农业大学林学院 江西省森林培育重点实验室 南昌 330045; 2. 江西农业大学鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室 南昌 330045; 3. 江西官山国家级自然保护区管理局 宜丰 336300;4. 江西省森林资源监测中心 南昌 330045)
我国人工林面积约0.69 亿 hm2,占森林总面积的31.8%(刘世荣等,2018),且这些人工林多为纯林,尤其是针叶纯林(FAO,2020)。由于树种单一、种植密度大、多次轮作等问题,导致出现林地养分失衡、土壤退化、生态系统稳定性差等现象(Heet al.,2022;Knokeet al.,2008;Forresteret al., 2006;张昌顺等,2005)。越来越多研究表明,营建针阔混交林可改善土壤养分状况,增强林分抗病虫害能力及提高林分生产力(Lianget al.,2016;Vilàet al.,2013)。初植(同龄混植)和补植(异龄混植)阔叶树被认为是人工林提质增效的有效措施之一。阔叶树通常比针叶树根系生物量高、生长速率快(马雪红等,2009),因此混交模式(阔叶树引入时间不同)可能影响针叶树对环境资源的获取和利用,进而产生不同的混交效应。Liu 等(2018)研究发现初植木荷(Schima superba)可促进杉木(Cunninghamia lanceolata)生长,而Bu 等(2020)研究表明补植木荷显著降低杉木生物量和组织养分含量。尽管对土壤养分动态和植物适应策略的混交影响已进行大量研究(Andiviaet al.,2016;Yesteet al.,2021;Dawudet al.,2017),但目前对针叶树生长的影响强度和方向存在较大争议(Linet al.,2022;Yanet al.,2019),其潜在机制仍难以明晰和界定。因此,探讨混交模式对针叶树的资源获取和生长的影响,可为我国亚热带针叶林管理提供科学依据。
资源获取,特别是营养获取直接影响生物体的能量和营养状态(Cahillet al.,2011)。以往研究表明,植物可通过根捕获和叶回收来维持其化学计量内稳态(Kouet al.,2017;Wuet al.,2021)。根捕获是一种地下养分获取策略,由吸收根实现(Kouet al.,2017),不仅受土壤养分有效性的影响,还受邻近植物的影响(Honget al.,2017;Yanet al.,2019),如补植深山含笑(Michelia maudiae)会抑制马尾松(Pinus massoniana)根系对无机氮的吸收(Liuet al.,2021)。叶回收是一种重要的地上养分保存策略,可降低植物对外部养分供应的依赖性(Killingbeck,1996;Chenet al.,2021)。Lin 等(2022)研究表明,在土壤养分贫瘠或种间竞争较强的混交林中,马尾松采取养分保存策略。由于每种策略都需要不同程度的成本投入,因此植物往往在根捕获和叶回收之间权衡(Kouet al.,2017;Wuet al.,2021)。阔叶树引入时间不同会导致初植和补植混交林间的土壤养分和种间关系不同,因此混交林中根系捕获养分的成本各异,针叶树可能采用不同的养分获取策略以应对阔叶树种的影响。目前尚不清楚初植和补植阔叶树是如何改变针叶树的养分获取策略。此外,除了捕获和回收外,养分在不同器官间的转运也是植物获取和利用的关键机制(Aldous,2002),但仍不清楚是否可通过植物养分转运过程反映根捕获和叶回收的权衡关系。
氮(N)和磷(P)是树木生长和养分循环的最基本和限制性元素(Chenet al.,2015)。由于受森林砍伐和强降雨的影响,我国亚热带红壤丘陵区的土壤N、P 元素大量流失(Wanget al.,2022),区域人工林的生态问题尤为突出,探讨不同混交模式对针叶树养分获取策略及其生长的影响至关重要。鉴于此,本研究选取红壤丘陵区广泛种植(栽培面积约3×106hm2,Dinget al.,2022)的湿地松(P. elliottii)林,基于长期初植和补植混交试验平台,分析湿地松养分获取和转运对初植和补植阔叶树响应的异同,旨在回答:1) 湿地松的养分获取策略是否会因混交模式不同而变化;2) 2 种混交模式如何调控湿地松的根捕获与叶回收的权衡;3) 植物体内的养分转运过程是否可以反映根捕获和叶回收的权衡关系。
1 研究区概况与研究方法
1.1 研究区概况
混交试验在江西省泰和县国家林业和草原局江西退化红壤森林植被恢复国家长期科研基地(26°44′N,115°04′E)进行。基地总面积133 hm2,属中亚热带季风气候,夏季温暖湿润,冬季凉爽干燥,年均降水量1 600 mm,年均气温18.6 ℃。海拔70~130 m,土壤类型为红壤(Fanget al.,2015)。
区域地带性植被类型为常绿阔叶林,但1990年以前因长期人为干扰,常绿阔叶林退化为次生灌木林。1991年,在除去杂灌和整地后,按完全随机试验设计,选用12 种针叶及阔叶树种:马尾松、湿地松、晚松(P. serotina)、火炬松(P. taeda)、杉木、木荷、枫香(Liquidambar formosana)、桉树(Eucalyptus robusta)、樟树(Cinnamomum camphora)、白栎(Quercus fabri)、刺槐(Robinia pseudoacacia)、臭椿(Ailanthus altissima),进行植树造林以改善区域环境、防止水土流失。其中,根据湿地松在当地的生长特性,以3 m×3 m 的行间距营建湿地松纯林、湿地松与木荷混交林(湿地松与木荷按1∶1 比例进行株间混交,简称初植混交林)。2006年,为评估针叶林的混交效应,在基地内随机选取5 个不同小班中的湿地松纯林进行中度间伐,并在林下均匀补植2年生木荷(种植在相邻两湿地松中间),形成补植混交林(湿地松与木荷按1∶1 比例株间混交,简称补植混交林)。
1.2 试验设计
2020年7月,基于长期混交试验平台,选择湿地松林龄为30年的湿地松纯林(slash pine plantation,SPP)、初植木荷混交林(initial mixed plantation,IMP)和补植木荷混交林(replanting mixed plantation,RMP)湿地松纯林16年后间伐补植木荷再生长14年)等3种林型各5 处(位于不同小班),按照相似的坡角(8%~10%)和方位角(南北方向)布设20 m×20 m 的样地。本研究共设置15 块样地,即为3 种水平5 次重复的完全随机化试验。此外,调查3 种林型的林分和根际土特征(表1)。
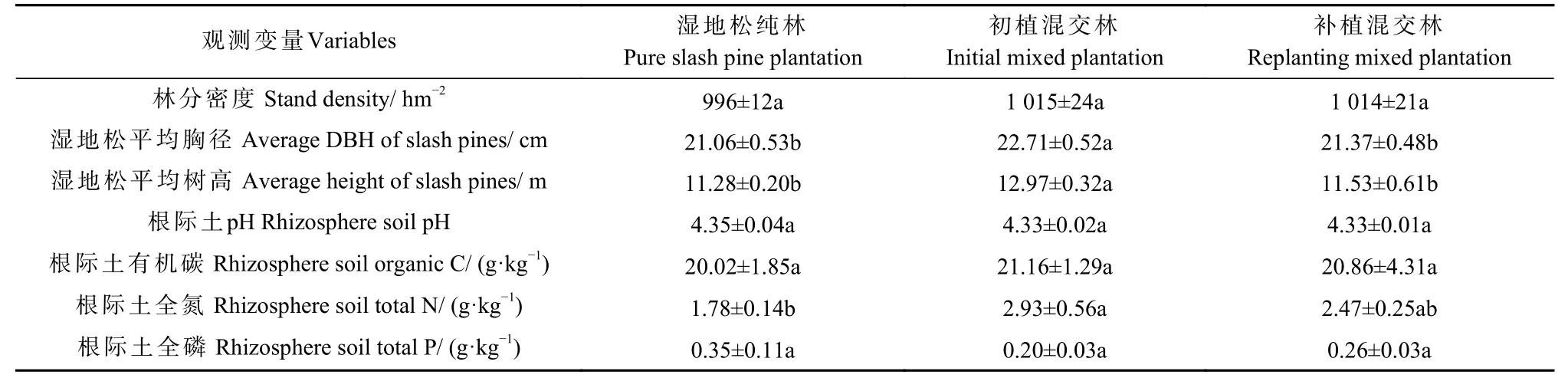
表1 3 种森林类型的林分特性和根际土特征①Tab. 1 Stand characteristics and rhizosphere soil properties in 3 forest types
1.3 样品采集
2020年7月,在各样地根据平均胸径和树高选取6 株湿地松作为标准木,收集每株标准木的细根、枝、新鲜叶、凋落叶和根际土壤,在距离树干基部0.5 m处(细根主要分布区)(Liet al.,2020)的东、西、南、北4 个方向各挖1 个20 cm(长)×10 cm(宽)×10 cm(深)的方形土块。去离子水清洗土块后,用手拣挑出直径小于2 mm 活细根,最末端的根尖为1 级根,2 个1级根交汇形成的根归为2 级根,2 个2 级根交汇形成的根归为3 级根,依此类推(Pregitzer,2002),其中1级根和2 级根被认为是吸收根(有完整皮层和较高菌根定植率,以吸收功能为主),3、4、5 级根被认为是运输根(无皮层和菌根定植,以运输功能为主)(Chenet al.,2015;Yinet al.,2020)。将每块样地中24 个土块吸收根和运输根分别混合成1 个样品,烘干至恒质量(65 ℃,72 h),研磨后过0.15 mm 筛,测定全氮(total nitrogen,TN)和全磷(total phosphorus,TP)含量。
用高枝剪从每株标准木冠层(高约8 m)东、西、南、北4 个方向上收集当年生小枝和长势良好的当年生叶作为新鲜叶。在每株标准木树冠下方放置5个1 m×1 m 的尼龙网凋落物收集框,采集新鲜叶时收集凋落叶。分别将每块样地中标准木的枝、新鲜叶和凋落叶混合成1 个样品,烘干至恒质量(65 ℃, 72 h)后研磨,测定TN 和TP 含量。
采用抖落法采集湿地松根际土壤样品(Rileyet al.,1970),在标准木树冠下随机挖取0~20 cm 具有完整根系的土体,轻轻抖落大块根系外围土,然后用刷子收集离根 4 mm 以内的土壤作为根际土壤(Primiciaet al.,2014)。同一样地中6 株标准木的根际土壤混合成一个样品,低温(4 ℃)带回实验室。土样过2 mm筛,部分用于测定NH4+-N、NO3−-N 和有效磷(available phosphorus,AP)含量;部分风干后测定pH 值、有机碳(organic carbon,OC)、TN 和TP 含量。
1.4 土壤理化分析
土壤pH 及养分含量测定参照《土壤农化分析》(鲍士旦,2000)中的相关方法:电位法测定土壤pH(土水比为1∶5);利用2 mol·L−1KCl 溶液浸提土壤后分别用靛酚蓝法和镉柱还原法测定NH4+-N 和NO3−-N 含量,矿质氮(mineral nitrogen,MN)含量为NH4+-N和NO3−-N 含量之和;利用0.5 mol·L−1NaHCO3浸提土壤后用钼锑抗比色法测定AP 含量;利用H2SO4-K2Cr2O7氧化法测定SOC 含量;土壤和植物样品经H2SO4消煮后分别用靛酚蓝法和钼锑抗比色法测定TN 和TP 含量。
1.5 根养分捕获能力、叶养分回收效率和树木养分转运因子的计算
根捕获能力(root capture ability,RCA)用于评估细根的养分捕获能力(Yeet al.,2014),计算公式为:
式中:NuAR和NuS分别为吸收根和根际土壤的养分含量。
养分回收效率(nutrient resorption efficiency, NuRE)用于量化针叶养分回收能力,计算公式为:
式中:NuFN为新鲜叶的养分含量;NuSN为凋落叶的养分含量;MLCF 为质量损失校正因子(Van Heerwaardenet al.,2003),针叶树的MLCF 为0.745(Vergutzet al.,2012)。
树木养分转运因子(translocation factors,TF)用于量化养分在不同器官间的转运能力(Wuet al., 2021),各相应器官的计算公式为:
式中:TFAR-TR、TFTR-TW和TFTR-FN分别为养分从吸收根到运输根、从运输根到枝、从运输根到新鲜叶的转运因子;NuAR、NuTR、NuTW和NuFN分别为吸收根、运输根、枝和新鲜叶中的养分含量。
1.6 统计分析
所有数据进行正态性检验,采用单因素方差分析(ANOVA)及最小显著差异法(LSD)检验3 类森林之间的各观测变量差异显著性(P<0.05)。整合湿地松纯林、初植混交林、补植混交林数据,利用Pearson’s相关分析分别检验初植和补植混交驱动的养分获取参数间的相关性。所有数据的分析和作图采用R(4.1.3)软件的agricolae、corrplot、ggplot2 包完成。
2 结果与分析
2.1 初植和补植混交对根际土壤和植株各器官养分含量的影响
初植和补植阔叶树显著提高湿地松根际土壤的NH4+-N 和MN 含量(P< 0.05),增幅为15.47%~19.48%;混交对NO3−-N 和AP 含量的影响较小(表2)。初植混交林和补植混交林之间的根际土壤养分含量均无显著差异(P> 0.05)。
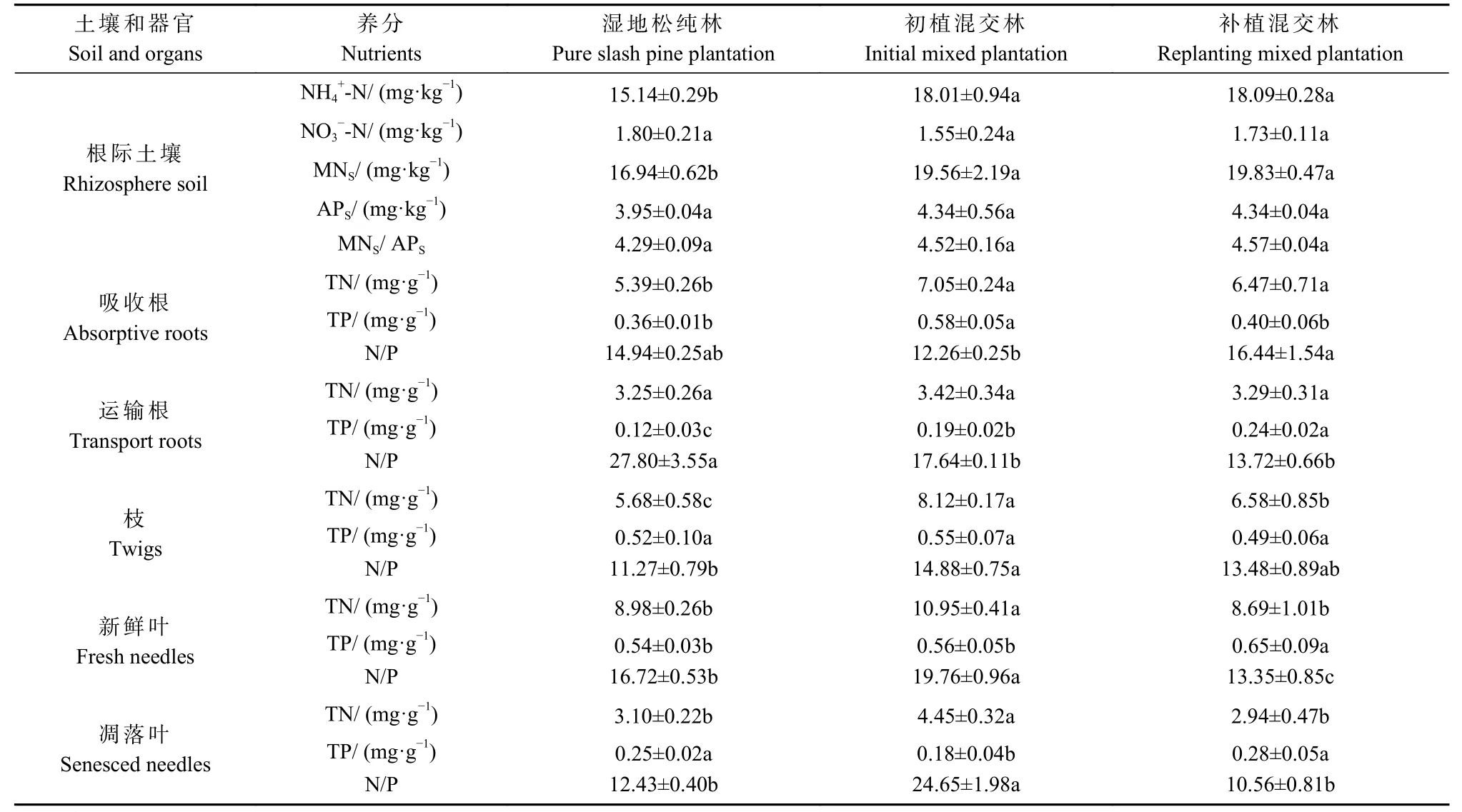
表2 3 类森林中湿地松根际土壤和器官的养分含量①Tab. 2 Nutrient contents in rhizosphere soil and tree organs of 3 forest types
整体看,混交阔叶树可改善湿地松的养分状况,但湿地松各器官的养分含量因混交模式而异(表2)。具体来说,混交显著提高吸收根的TN 和运输根的TP 含量,其中初植混交林吸收根的TP 含量显著高于补植混交林,而运输根的TP 含量呈相反趋势;初植混交林的枝、新鲜叶和凋落叶的TN 含量显著高于补植混交林,但新鲜叶和凋落叶的TP 含量显著低于补植混交林。此外,混交模式改变各器官的N/P,初植混交林中运输根、新鲜叶和凋落叶的N/P >16 且显著高于纯林和补植混交林(N/P < 14)。
2.2 初植和补植混交对根养分捕获能力和叶养分回收效率的影响
初植混交林中根的N、P 捕获能力高于纯林和补植混交林,而补植混交林与纯林的根N、P 捕获能力均无显著差异(图1)。初植混交林中根N 捕获能力分别比纯林和补植混交林高14.04%和11.04%(图1A),类似地,根P 捕获能力分别提高46.16%和43.88%(图1B),表明初植混交林中湿地松的根养分捕获能力比在补植混交林中更强。
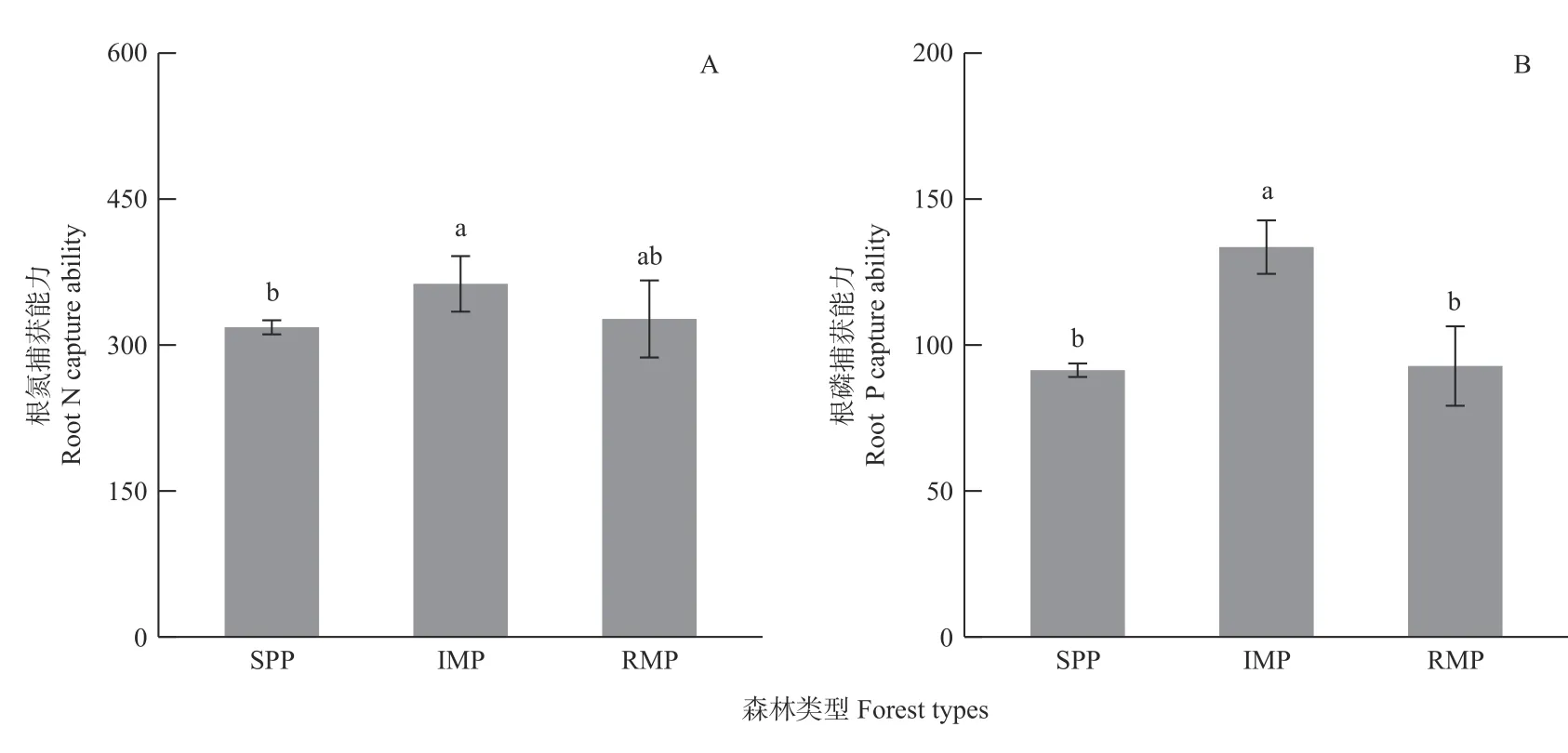
图1 3 类森林中湿地松的根氮(A)和磷(B)捕获能力Fig. 1 Root N (A) and P (B) capture ability of slash pine in 3 forest types
初植混交林中叶N 回收效率显著低于纯林和补植混交林,而叶P 回收效率显著高于纯林和补植混交林;补植混交林与纯林的叶N、P 回收效率均无显著差异(图2)。初植混交林中叶N 回收效率分别比纯林和补植混交林低6.07%和6.47%(图2A),但叶P回收效率分别提高15.49%和12.05%(图2B)。表明初植混交林中湿地松对N 元素优先采取根N 捕获策略,而对P 元素采取根捕获和叶回收2 种策略;补植阔叶树对湿地松的养分获取策略影响较小。
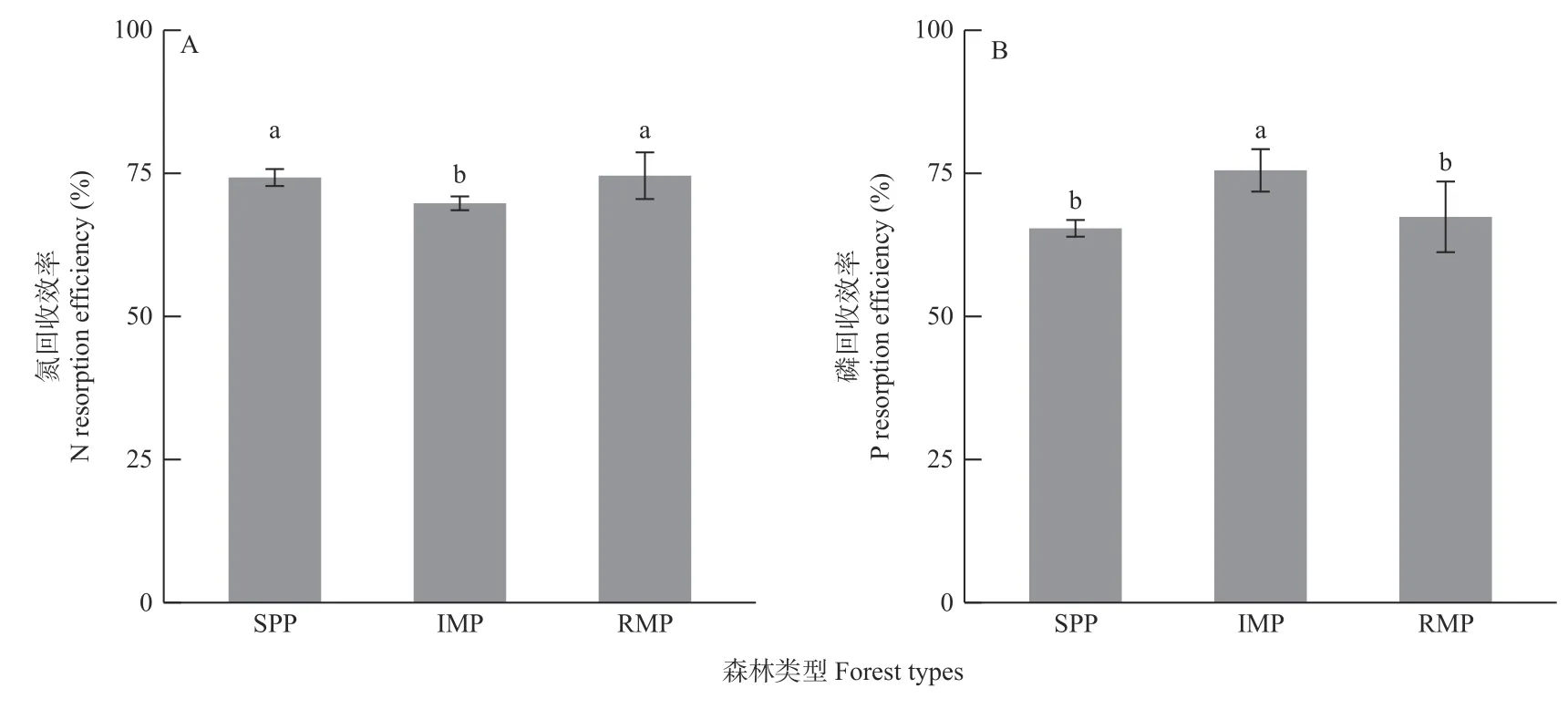
图2 3 类森林中湿地松的叶氮(A)和磷(B)回收效率Fig. 2 Needles N (A) and P (B) resorption efficiencies of slash pine in 3 forest types
2.3 初植和补植混交对植物体内的养分转运因子的影响
与纯林相比,初植和补植混交林中湿地松吸收根向运输根的N 转运分别降低19.90%和15.16%(图3A);初植混交林中的运输根向枝和新鲜叶的N 转运分别比纯林高36.05%和15.61%,比补植混交林高19.69%和21.39%(图3C, E)。补植混交林与纯林之间在从运输根向枝、新鲜叶的N 元素转运上无显著差异(图3C, E)。
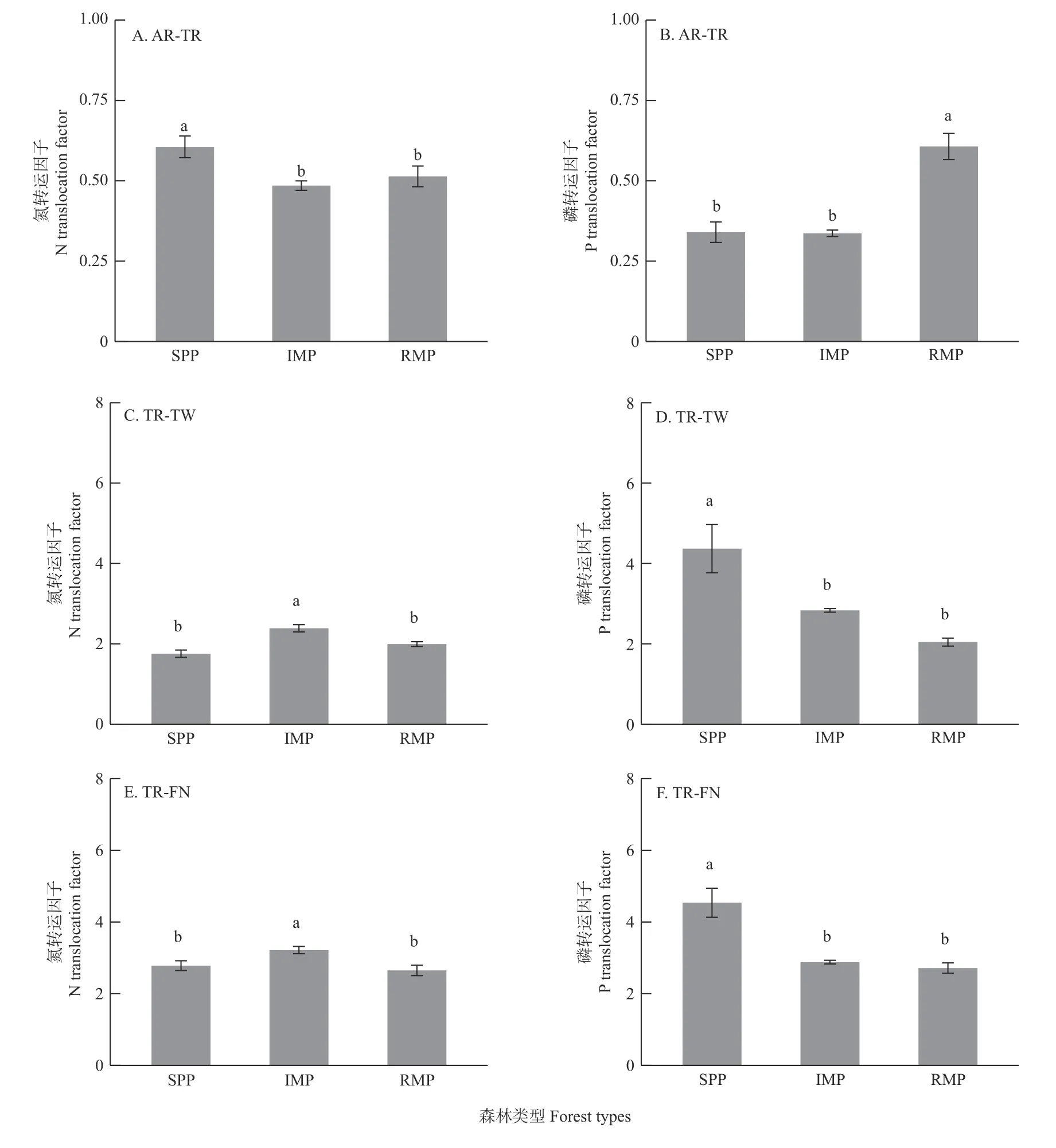
图3 3 种森林类型湿地松的氮和磷转运因子Fig. 3 N and P translocation factors of slash pine in 3 forest types
此外,补植混交林中吸收根向运输根的P 转运分别比纯林和初植混交林高78.48%和80.20%,而纯林和初植混交林之间在从吸收根向运输根的P 元素转运上无显著差异(图3B);与纯林相比,初植混交林中从运输根向枝和新鲜叶的P 转运分别降低35.13%和36.52%,补植混交林分别降低53.21%和40.17%(图3D、F);初植混交林和补植混交林之间在从运输根向枝和新鲜叶的P 转运上无显著差异(图3D、 F)。结果表明,初植混交林促进湿地松将N 元素向地上部分的转运,但降低湿地松将P 元素向地上部分的转运。
2.4 养分获取变量的相关性分析
初植混交驱动下,湿地松MNS和APS与NRE呈显著负相关,而与根捕获能力和转运无显著相关;根NRCA与N 的TFAR-TR呈显著负相关,与N 的TFTR-TW和TFTR-FN呈显著正相关;根PRCA与P 的TFAR-TR无显著相关,与P 的TFTR-TW和TFTR-FN呈显著负相关;N 回收效率与N 的TFTR-TW呈显著负相关,P 回收效率与P 的TFTR-FN呈显著负相关(图4A)。
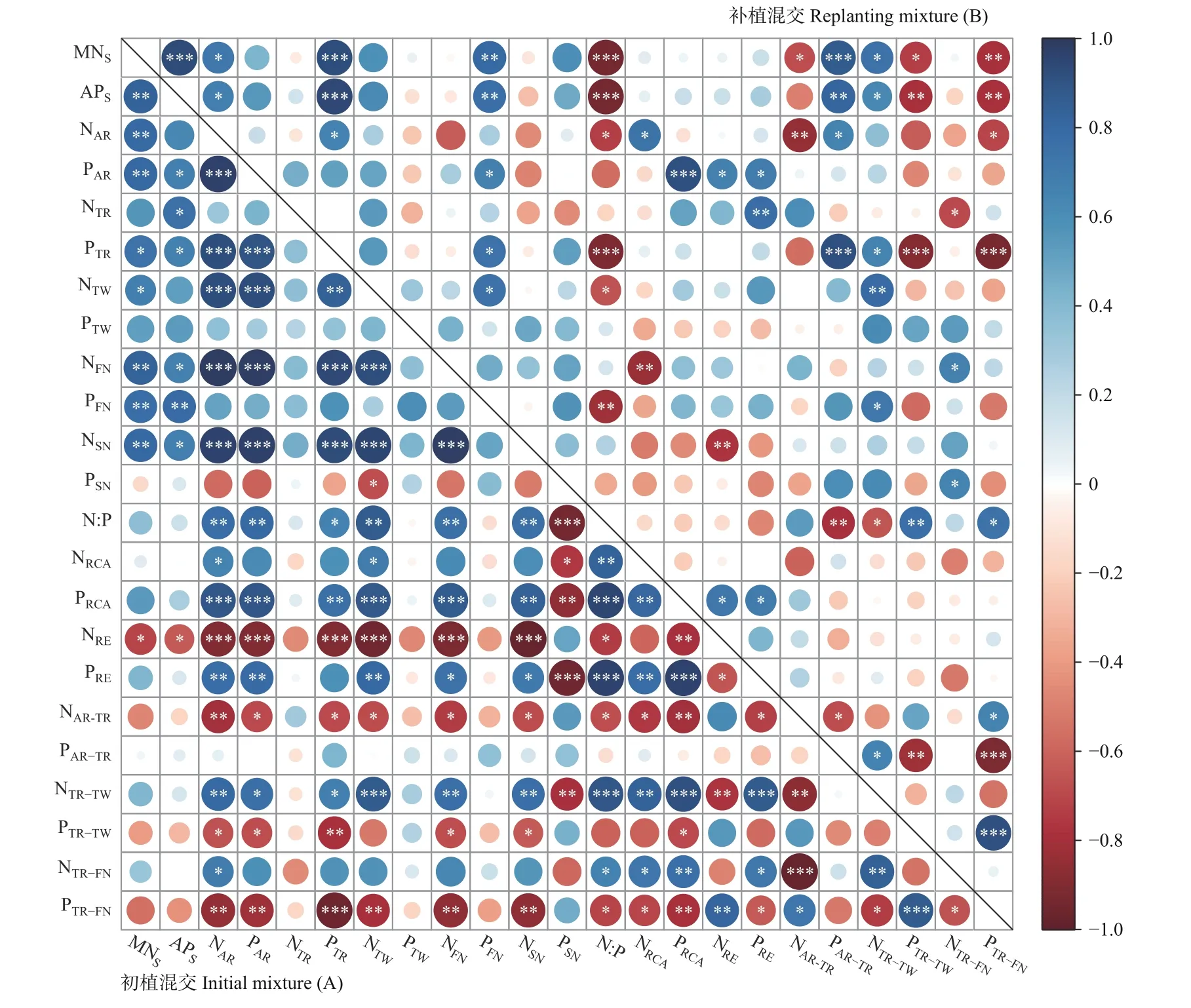
图4 初植混交(A)和补植混交(B)驱动的养分获取变量间的相关性Fig. 4 Correlations among main variables of nutrient acquirement in slash pine plantations driven by initial mixture (A) andreplanting mixture (B)
补植混交驱动下,湿地松根际土壤养分与N 的TFAR-TR、P 的TFTR-TW和TFTR-FN显著负相关,与P 的TFAR-TR和N 的TFTR-TW呈显著正相关,而与根捕获能力和叶回收无显著相关;根捕获能力和叶回收效率与转运均无显著相关(图4B)。
3 讨论
3.1 混交模式对湿地松根养分捕获能力的影响
湿地松根的N、P 捕获能力在初植混交后显著提高,而补植混交后无显著变化(图1),表明湿地松根养分捕获能力因混交模式不同而异,这回答了科学问题1。已有研究表明,根对养分的捕获能力与土壤养分有效性有关,一方面,混交阔叶树提高土壤养分有效性,因此湿地松根养分捕获能力增强;另一方面,随着土壤养分有效性增加,植物从土壤中获取养分的成本降低,因此植物更倾向于采取根捕获策略,从而使根捕获能力提高(Wrightet al.,2003)。本研究中,初植阔叶树提高根际土壤N 的有效性,促进根N 捕获能力;虽然初植阔叶树并没有改变根际土壤P 的有效性,但由于N、P 相互作用,N 有效性增加会影响P 的相对有效性(Olanderet al.,2000),进而促进根对P 的捕获。与初植混交林不同,补植混交林内湿地松根捕获能力没有随土壤养分有效性提高而显著变化,这可能是由于两混交林中湿地松与木荷的种间竞争不同。Filipescu 等(2007)研究表明,由于混交年限不同,群落中的种间竞争存在明显差异。Cabal等(2020)提出的进化稳定策略表明,在地下养分竞争条件下,植物在邻近茎干处产生大量细根,以增强植物对土壤资源的吸收能力。Yan 等(2019)研究发现增加地下种间竞争后,湿地松根系会增加根系的生长和菌根定殖,以提高根系对养分的捕获能力。与初植混交林相比,补植混交林内的湿地松根系受到更小的种间竞争,湿地松分配更少资源用于地下养分获取,根对养分的捕获能力未发生显著变化。
3.2 混交模式对湿地松叶养分回收的影响
叶养分回收是植物将衰老组织的养分转运到功能活跃组织的过程,是一种养分保存策略(Aerts,1996)。本研究表明,初植混交后湿地松叶N 回收效率降低,P 回收效率提高;此外,叶P 回收效率(75.52%)高于叶N 回收效率(69.76%)(图2),这与养分限制控制植物叶回收效率的研究结果一致(Chenet al.,2021)。以往研究表明,植物养分回收通常受到N、P 限制调控,如果植物生长受N 限制,则相对P 而言,植物回收更多的N,且N 回收效率高于P 回收效率;相反如果是P 限制,更多的P 被回收(Güsewell,2005;Chenet al.,2021)。植物叶片的N/P 可用来判断植物生长的养分限制状况(Güsewell,2004),通常认为N、P>16 为P 限制,N、P<14 为N 限制(Güsewell,2004)。初植混交林中湿地松针叶N/P 为19.76,显著高于纯林的16.72,显然初植阔叶树加了湿地松生长的P 限制。P 限制加剧后,植物回收获取P 的成本相比与根获取土壤中P 的成本降低,相反N 回收的相对成本提高,这使得初植混交林中湿地松叶片N 和P 回收分别呈现降低和提高的现象。此外,针叶N/P 与NRE显著负相关,而与PRE显著正相关,这也表明初植混交林中湿地松叶养分回收效率受养分限制调控。
补植混交后,湿地松针叶N/P 为13.35,表明湿地松生长由P 限制转为N 限制,但湿地松叶N 和P 回收效率均无显著变化(图2)。与初植混交不同,补植混交后叶的养分回收效率未随养分限制变化而变化,这可能与补植后土壤养分有效性提高有关。大量研究表明,土壤养分有效性对植物回收效率有影响,如Enoki 等(1999)研究发现,黑松(Pinus thunbergii)叶片养分回收效率随土壤N 有效性降低从43%提高到77%;赵琼等(2010)研究发现,在土壤N 供应充足的情况下,通过添加N 肥提高土壤N 有效性也会显著降低叶N 回收效率。土壤养分有效性提高,根系获取土壤养分的成本降低,而回收获取养分的成本则相对提高,因此湿地松可从外部环境获取更多低成本的养分,而不必增加对内部养分的依赖。
3.3 混交模式驱动的养分获取策略及其内部养分转运过程的动态变化
植物养分获取策略多种多样,其中根捕获和叶回收是最常见、最主要的2 种方式(Kouet al.,2017;Wuet al.,2021)。以往研究表明,根捕获和叶回收往往存在权衡(Kouet al.,2017;Wuet al.,2021)。然而,最近研究表明,根捕获和叶回收存在协同响应(Songet al.,2022)。初植阔叶树改变湿地松根捕获能力和叶回收效率,且NRCA与NRE呈负相关,PRCA与PRE呈显著正相关,说明初植混交林中湿地松对N 的根捕获和叶回收存在权衡策略,而对P 存在协同策略,这回答了科学问题2。初植阔叶树提高土壤养分有效性,湿地松根系从土壤中获取养分的成本降低,湿地松倾向于根捕获策略获取养分而不是叶回收策略(Wrightet al.,2003),因此湿地松的根N 捕获能力提高和叶N 回收效率降低。与N 获取策略不同,初植混交林中湿地松根P 捕获和叶P 回收均提高,对P 元素采取根捕获和叶回收的协同策略。湿地松对P 捕获和回收的协同,可能与初植混交后P 限制加剧,因而需要大量的P 维持自身的化学计量稳定有关。补植混交后,湿地松N 和P 的根捕获能力和叶回收均无显著变化,湿地松对地上和地下的养分获取策略的偏好无显著变化。与初植混交相同,补植混交后土壤养分有效性提高,湿地松根系获取土壤养分的成本降低,但补植后养分限制状况发生改变,同样降低湿地松内部养分回收所需的成本。因此,补植混交后根捕获和叶回收对养分获取的相对成本并未改变,湿地松的养分获取策略无显著变化。
本研究表明,初植混交后湿地松养分转运与养分获取间存在显著相关关系(图4A),回答了科学问题3,即养分转运可以体现植物养分获取策略的变化。初植混交后,N 的TFAR-TR降低并与N 捕获能力显著负相关,表明湿地松吸收根向运输根N 转运越少,植物对N 的捕获能力越强;N 的TFTR-TW和TFTR-FN均提高并与N 捕获能力显著正相关,而与N 回收显著负相关,说明植物向地上的N 转运越多,植物根系的N捕获能力越强,而N 回收效率越低;P 的TFTR-FN降低并与捕获能力显著负相关,与回收显著负相关,也说明湿地松向针叶的P 转运越多,植物根系N 捕获能力和回收效率越强。Wu 等(2021)在N、P 添加下杉木对微量元素获取和转运的研究中也发现,捕获和回收之间的权衡可通过转运来表征,这与本研究结果一致。
本研究还发现,补植混交后湿地松养分转运与养分获取间虽无显著相关(图4B),但也反映湿地松对养分获取的相对变化。补植混交后,湿地松降低了吸收根向运输根的N 转运,这表明湿地松对根系和根系捕获的投入。Lin 等(2022)研究也发现,补植混交后湿地松将更多的养分投入根系用于开发土壤养分。并且,湿地松吸收根向运输根的N 转运与土壤矿质N 含量显著负相关,显然补植混交后湿地松对吸收根和运输根的投入受土壤养分有效性的调控。这一结果与Song 等(2022)的研究一致,土壤养分有效性的提高降低根系开发单位土壤养分的成本,因此湿地松将更多养分投入吸收根用于地下养分获取。补植混交后,湿地松的P 获取策略没有发生显著变化,而运输根向枝叶的转运减少,P 转运与获取相关性不大,这可能与湿地松生长的养分限制变化有关。有研究发现树木的养分转运也受到植物生理中“源-汇”调控(周玉泉等,2019),补植混交后湿地松由P限制转为N 限制,湿地松枝叶对P 的需求降低,运输根成为P 的“汇”累积P,而当外界环境或自身生长发生改变,湿地松枝叶对P 的需求提高,运输根又成为P 的“源”提供P。
4 结论
初植和补植木荷显著提高湿地松根际土壤的NH4+-N 和MN 含量。混交阔叶树,尤其是初植混交显著提高植物各器官的N、P 含量。因此,混交模式显著影响土壤和植物的养分含量,初植林的混交效应优于补植林。
与纯林相比,初植林中湿地松的根养分捕获能力提高,叶N 回收效率降低,叶P 回收效率提高,表明初植林中湿地松对N 元素采取根获策略而非叶回收策略(权衡),而对P 元素采取根捕获和叶回收的协同策略;种间竞争使补植林中湿地松的根捕获和叶回收效率均无显著变化,湿地松养分获取策略不变。
初植林中湿地松优先采用根N 捕获策略,所以运输根向枝、叶的N 转运提高;由于P 是初植林中湿地松生长的限制性元素(N/P >16),P 获取的协同策略促使运输根向枝、叶的P 转运下降。因此,N、P 在植物体内的转运过程反映了根捕获和叶回收的权衡关系。
