中亚热带喀斯特森林树木功能型的生态位特征*
2024-02-01庞庆玲徐超昊张忠华
胡 刚 庞庆玲 胡 聪 徐超昊 张忠华
(1. 南宁师范大学广西山地生态系统动植物进化与保护重点实验室 南宁 530100;2. 南宁师范大学地理科学与规划学院 南宁 530100)
生态位是物种对多种环境因子适应的综合结果,反映物种在群落中的时空位置、功能和地位(Hutchinson,1957;Pielou,1972)。生态位宽度和生态位重叠指数是生态位研究中常用的2 种量化指标,其中生态位宽度体现物种对环境资源利用状况的尺度,表征物种的生态适应性和分布幅度(Smith,1982;Carscaddenet al.,2020),生态位重叠指数反映2 个物种对同一资源的利用效率和共享程度,重叠指数较大预示2 个种可能具有相似的生态学和生物学特性或对资源的需求有一定互补性(Hurlbert,1978;龚雪伟等,2017)。不同物种在环境资源利用和分布上的差异体现生存策略和进化过程(Petersonet al.,2011;Pastoreet al.,2021),生态位理论是评价物种和生态因子相互关系,解释种间关系、物种多样性格局、群落构建与进化过程的基本理论,一直是生态学研究的核心问题(Silvertown,2004;Levineet al.,2009;陈磊等,2014;储诚进等,2017;Atwateret al.,2018;Costa-Pereiraet al.,2019;Godoyet al.,2020;Pastoreet al.,2021;牛克昌等,2022)。
植物功能型指生态系统中扮演着相似角色且对环境条件表现出相似响应的一组具有相似特征(形态或生理特征)的植物群组(Woodwardet al.,1996;胡楠等,2008),通过植物功能型划分可将大量物种分类成数量相对较少的组合,从而区分对环境变化有类似响应或影响的植物类群(Duckworthet al., 2000;Lavorelet al.,2007)。群落是种群的集合,同时也可看作不同植物功能型的组合,同一功能型内物种往往具有结构、生理等方面的共性,而在生境相似的局域尺度上,不同功能型植物对同种资源获取的方式或能力不同,对环境的适应策略和生理过程可能会出现分化(Golluscioet al.,1993;Lamontet al.,2021),因此,研究植物功能型可在个体、群落以及生态系统水平上将植物生理、生活史过程以及适应策略等联系在一起(Verheijenet al.,2016)。
国内外学者对植物种群的生态位特征进行了大量研究(Anthwalet al.,2008;O'Kaneet al.,2011;龚雪伟等,2017;董雪等,2020;Xuet al.,2021;Huet al.,2022),如Anthwal 等(2008)分析印度喜马拉雅区域草丛中优势种和次优势种的生态位宽度和生态位重叠特征,揭示了地形、竞争作用等对种群生态位的影响;Xu 等(2021)通过计测生态位宽度和生态位重叠指数分析我国喀斯特森林中濒危植物薄叶金花茶(Camellia chrysanthoides)与其他共生物种间的潜在竞争关系,并基于此评估了该物种的分布和保育现状;Hu 等(2022)研究表明,我国青海湖流域高寒草地荒漠化致使优势植物的生态位宽度下降,并且改变了种间生态位重叠指数和竞争关系。上述研究均通过计测种群的生态位宽度和种间的生态位重叠指数分析物种的适合度、种间关系或物种共存格局,迄今鲜有研究分析群落中不同植物功能型间的生态位特征(Rutherfordet al.,1995;金超等,2021)。从植物功能型维度开展森林树木的生态位特征研究,有助于比较不同树木功能型组合的资源空间占据状况以及竞争排斥关系,从而推断群落构建和物种多样性维持的潜在机制。
喀斯特地貌是世界上广泛分布的一种地质景观类型,占全世界近15%的土地面积(袁道先, 1994;Fordet al.,2007)。我国喀斯特地貌区约344 万km2,约占国土面积的36%、占世界喀斯特(2 200 万km2)分布面积的15.6%,其中贵州、广西、云南等8 个省(区)有约51 万km2裸露碳酸盐岩区,占其总土地面积的5.8%(Jianget al., 2014)。由于喀斯特生态系统脆弱,在人为干扰和资源不合理利用等多因素综合作用下,生态系统易于退化,甚至出现石漠化现象(Wanget al.,2019)。然而,喀斯特环境孕育着丰富的生物多样性,是物种形成演化和生物多样性保育的“诺亚方舟”(Clementset al.,2006)。喀斯特森林对维持岩溶脆弱生态系统的结构和功能稳定性极为重要,同时也可作为石漠化地区植被近自然恢复的参考样本。近年来,国内学者针对喀斯特森林的植物生理生态(Nieet al.,2017)、种群结构与动态(Zhanget al., 2010)、植被恢复与重建(Tonget al.,2018)等开展了丰富的研究工作,而对喀斯特森林树种生态位特征的研究均基于种群水平(黄甫昭等,2018;李婷婷等,2021;Xuet al.,2021),未从植物功能型维度探讨树木的生态位特征。
鉴于此,本研究以中亚热带喀斯特常绿落叶阔叶混交林为对象,在样地调查基础上根据冠层高度、种群大小、耐荫性和种子传播方式将样地中40 种树木划分为不同功能型,对不同树木功能型组合的生态位宽度和生态位重叠指数进行量化分析,并探讨:1) 喀斯特常绿落叶阔叶混交林中不同树木功能型的生态位宽度有何差异及其差异形成有何潜在原因;2) 不同树木功能型间的生态位重叠存在怎样的变化以及这种变化的潜在原因。通过分析喀斯特森林树木功能型的生态位特征,推断不同树木功能型对资源的利用程度和种间竞争差异,以期为喀斯特地区的植被恢复与重建提供理论指导。
1 研究区概况
研究区位于贵州省荔波县茂兰国家级自然保护区(107°52′10″—108°05′40″E,225°09′20′′—25°20′50″N),该保护区被联合国教科文组织纳入“国际人与生物圈保护区网络”,为“中国南方喀斯特”世界自然遗产地的重要组成部分,其主要保护对象为亚热带喀斯特常绿落叶阔叶混交林生态系统。茂兰保护区拥有地球同纬度地区残存下来的连片面积最大、保存完整且原生性强的喀斯特森林生态系统,保护区总面积21 285 hm2,主要地貌类型为由石灰岩及白云岩构成的典型喀斯特峰丛洼地和峰丛漏斗,海拔多在800 m 以上。属中亚热带季风湿润气候,气候温和,雨量充沛,年均气温15.3 ℃,7月平均气温26.4 ℃,1月平均气温8.3 ℃,≥10 ℃年积温5 727.9 ℃,平均年降水量1 320.5 mm,平均年蒸发量1 343.6 mm,全年平均空气相对湿度83%,年日照时数1 272.8 h,日照百分率29%。常见乔木有西南米槠(Castanopsis carlesiivar.spinulosa)、青冈(Cyclobalanopsis glauca)、杨梅叶蚊母树(Distylium myricoides)、 圆果化香(Platycarya longipes)和黄杞(Engelhardtia roxburghiana)等;灌木以榄绿粗叶木(Lasianthus japonicus)、大果卫矛(Euonymus myrianthus)和柄果海桐(Pittosporum podocarpum)等为主; 草本植物有边缘鳞盖蕨(Microlepia marginata)、翠云草(Selaginella uncinata)和江南卷柏(S. moellendorffii)等。
2 研究方法
2.1 样地调查
在茂兰国家级自然保护区常绿落叶阔叶混交林内建立1 块100 m×100 m 样地,将样地划分为100 个10 m×10 m 样方,调查每个样方内所有胸径(DBH)≥1 cm 的木本植物的种名、相对坐标、胸径、树高、冠幅和生长状况等数据。具体调查方法参见文献Zhang 等( 2012)。
2.2 树种选择与功能型划分
样地调查记录到DBH ≥ 1 cm 的木本植物达199 种,随机选取40 种树木作为研究对象。参照Wang 等(2010)方法,按冠层高度、种群大小、耐荫性和种子传播方式划分树木功能型(表1)。冠层高度(H)根据每种树的平均高度确定,其中上层种高度H≥12 m,中层种高度8 m ≤H< 12 m,下层种高度3 m ≤H< 8 m;根据种群多度(A)确定常见种、偶见种和稀有种,其中常见种A≥ 20,偶见种5 ≤A< 20,稀有种1 ≤A< 5;耐荫性根据《贵州植物志》、《中国植物志》中树种生物学特征介绍并结合野外观察来确定,其中耐荫性分为喜光、中生和耐荫3 种;根据种子的质量、形态和动物的取食喜好程度,并基于野外观察将种子传播方式分为风力、重力和动物3 种类型。需要说明的是,一种植物的种子传播方式可能同时有多种途径,本研究选择其主要的传播方式,如青冈的种子同时受重力和动物传播,但以动物传播为主。
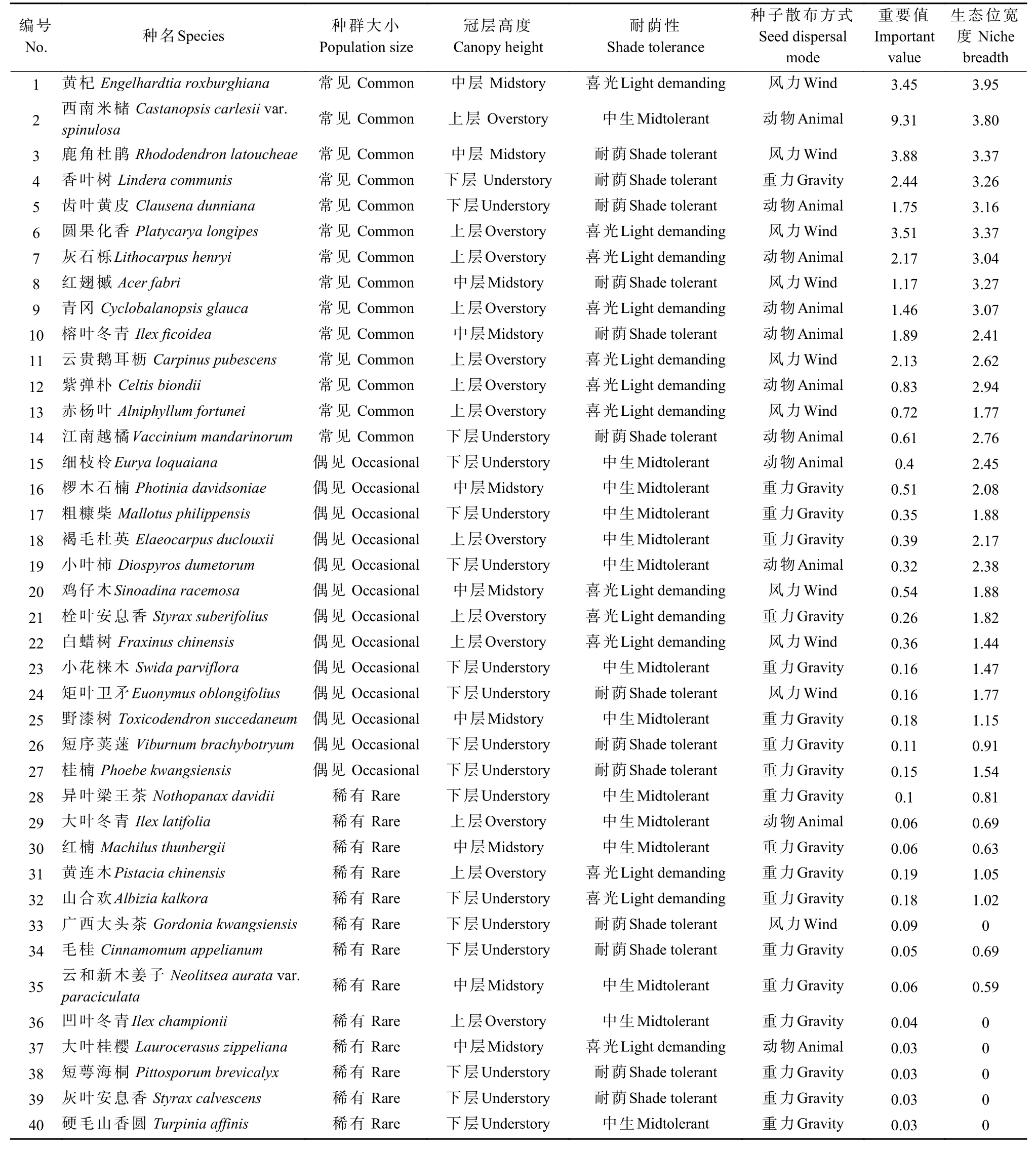
表1 喀斯特森林40 种树木的功能型划分、重要值和生态位宽度Tab. 1 Classification of tree functional types,important values, and niche breadth of 40 tree species in a karst forest
2.3 生态位宽度与重叠的计测
基于10 m×10 m 样方调查数据,计测40 种树木的重要值(IV),计算方法(张金屯,2018)为:
式中:RA 为相对多度,指某种的个体数占全部种个体总数的比例;RF 为相对频度,指某种的频度占全部种频度之和的比例;RD 为相对优势度,指某种的胸高断面积之和占全部种胸高断面积之和的比例。
采用Shannon-Wiener 指数(Shannonet al.,1949)计测生态位宽度,其公式为:
式中:Bi为种i的生态位宽度;Pij=nij/Ni,为种i在第j种资源状态下的重要值占该种所有重要值的比例;r为资源状态数。Bi越大,说明物种生态位越宽,利用的资源总量越多,竞争力越强。
采用Pianka 指数(Pianka,1973)表征生态位重叠指数,其公式为:
式中:Oik为2 个种群的生态位重叠指数,Oik越大,说明2 个种群的生态位重叠程度越大;Pkj为种k在第j种资源状态下的重要值占该种所有重要值的比例。
上述2 种指数计算在R 语言环境下运行SPAA软件包完成。
2.4 统计分析
采用生态位宽度和生态位重叠指数的平均值量化树木功能型的生态位特征,如喜光种生态位重叠指数为所有喜光种种对间重叠指数的平均值,喜光种与耐荫种间的生态位重叠指数为所有喜光种与耐荫种种对间重叠指数的平均值。采用单因素方差分析(one-way ANOVA)或非参数检验(nonparametric test)分析40 种树木的生态位宽度和生态位重叠指数在同种功能型内和不同功能型间的差异显著性。比较前,对数据进行描述性统计和Shapro-Wilk 检验,判断其是否符合正态分布,服从正态分布的数据采用单因素方差分析比较数据间的差异,不服从正态分布采用Kruskal-Wallis H 法进行非参数检验。统计数据用平均值±标准差表示。采用软件SPSS 22.0(SPSS Inc.,Chicago, USA)进行上述分析。
3 结果与分析
3.1 不同树木功能型的生态位宽度
由表1 可知,40 种树木中,生态位宽度在0~1 区间的占35.0%,1.1~2 和2.1~3 区间的均占22.5%,3.1~4区间的占20.0%,大部分树木的生态位宽度较小。生态位宽度最大的是黄杞(3.95),其次是西南米槠(3.80)、圆果化香(3.37)和鹿角杜鹃(3.36),这几个种均为群落优势种。
图1 表明,生态位宽度表现为常见种>偶见种>稀有种,且三者间差异显著(P<0.05);重力传播种的生态位宽度显著低于动物和风力传播种(P<0.05),动物和风力传播树种间的生态位宽度差异不显著;生态位宽度在不同冠层高度以及不同耐荫性树种间差异不显著。

图1 喀斯特森林树木各功能型生态位宽度Fig. 1 Niche breadth of various tree functional types in a karst forest
3.2 不同树木功能型的生态位重叠指数
由图2 和3 可知,40 种树木的种对间生态位重叠指数为0~0.8,其中0~0.2 的有695 对,占种对总数的89.1%,而0.2~0.4、0.4~0.6 和0.6~0.8 的种对数分别只有67、15 和3 对,合计占种对总数的10.9%。生态位重叠指数大于0.6 的仅3 对,分别为云和新木姜子与小花梾木(0.72)、大叶冬青与硬毛山香圆(0.67)、紫弹朴与黄连木(0.65)。
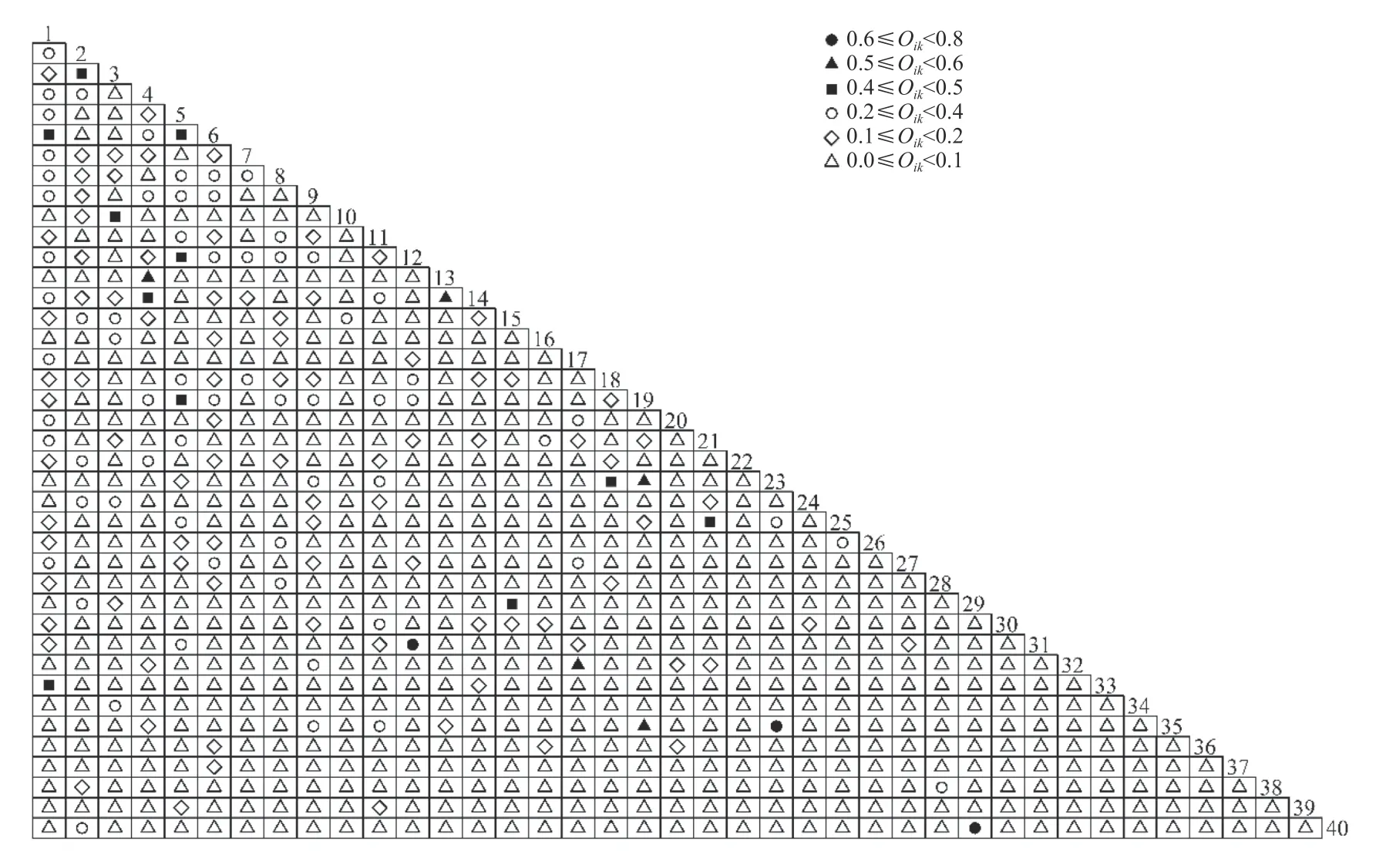
图2 喀斯特森林40 种树木间生态位重叠指数半矩阵图(物种编号同表1)Fig. 2 Semi-matrix graph show niche overlap index of 40 tree species in a karst forest (species number is the same as Tab. 1)
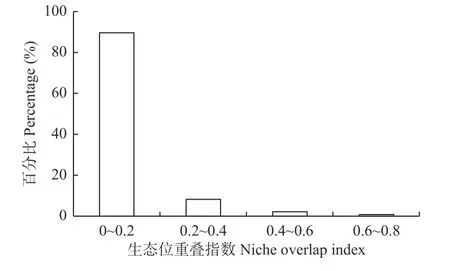
图3 喀斯特森林40 个树种生态位重叠指数的分级比例Fig. 3 Graded proportion of niche overlap index for 40 tree species in a karst forest
生态位重叠指数表现为常见种>偶见种>稀有种,且三者间存在显著差异(P<0.05,图4A);从冠层高度看,生态位重叠指数表现为上层种>中层种>下层种,其中上层种内的生态位重叠指数显著高于下层种(P<0.05,图4C);在耐荫性方面,生态位重叠指数表现为喜光种>中生种>耐荫种,中生种与喜光种和耐荫种间的差异均不显著,而喜光和耐荫树种间存在显著差异(P<0.05,图4E);重力传播种内的生态位重叠知识显著低于动物和风力传播种(P<0.05),而动物和风力传播种间则无显著差异(P>0.05,图4G)。

图4 喀斯特森林树木功能型内和功能型间生态位重叠指数的比较Fig. 4 Change in values of niche overlap index within and between tree functional types in a karst forest
常见种与偶见种间的生态位重叠指数最高,其次为常见种与稀有种间以及偶见种与稀有种间,3 组间存在显著差异(P<0.05,图4B);不同冠层高度树种之间的生态位重叠指数差异不显著(图4D);中生种与耐荫种间的生态位重叠指数显著低于喜光种与中生种及喜光种与耐荫树种(P<0.05,图4F);动物传播种与风力传播种间的生态位重叠指数显著高于动物传播种与重力传播种间以及风力传播种与重力传播种间(P<0.05,图4H)。
4 讨论
4.1 喀斯特森林树木功能型的生态位宽度
喀斯特生境以碳酸盐岩为主要基质,地表土壤稀薄,地形复杂,易形成各种小生境,不同树种适应不同小生境从而出现生态位分化,导致多数树种对生境的资源利用能力较低,种间竞争相对较低(Zhanget al.,2013;黄甫昭等,2018)。一项基于Meta分析的研究证明,环境异质性可广泛促进不同空间尺度、不同生物群落中物种的丰富度(Steinet al.,2014)。本研究表明,中亚热带喀斯特常绿落叶阔叶混交林中40 种树木的生态位宽度普遍较小(0~4),喀斯特生境的高度异质性可能是导致树种生态位宽度较小的原因之一。同时,普遍较低的生态位宽度和异质的生境有利于多物种共存以及形成复杂多样的群落结构。本研究中,仅黄杞、西南米槠、圆果化香和鹿角杜鹃等少数树种的生态位宽度较大,这些种往往是资源竞争中占据优势地位的树种,其对生态系统功能(如生产力)具有重要影响。
常见种的生态位宽度最大,其次是偶见种和稀有种。常见种的种群数量多,且部分为优势种,其对资源的综合利用能力较强,具有较大的生态位宽度,而稀有种数量较少且出现频率较低,适应性和对资源的利用能力较弱,占据的生态位较小。该结果同时表明,多度可度量种群生态位宽度变化,是体现种群或植物功能型资源利用和占据能力的有效指标(Carscaddenet al.,2020)。
本研究发现,重力传播种的生态位宽度显著低于动物和风力传播种,种子传播方式决定种群的空间分布格局与幼苗的增补方式(Mendeset al., 2021)。一般而言,重力传播种往往以母树为中心进行种群更新,扩散距离有限(Nathanet al.,2000);同时,喀斯特生境岩石裸露致使土壤分布不均匀和不连续,在一定程度上阻碍种子遇到适合萌发的微生境。此外,受病原菌或食草动物引起的距离和密度制约性死亡效应的影响,母树周围可能不利于同种幼苗的增补(Janzen,1970;Connell,1971)。因而,扩散限制、生境制约等多种因素影响下,重力传播种占据空间资源的能力较弱(Wiegandet al., 2021)。然而,依靠动物和风力传播的树种可以避开上述不利因素的影响,扩散至有利于定植的生境,从而维持其在群落中的相对优势地位。因此,重力、动物和风力传播种的生态位宽度变化体现了不同功能型树木的生活史策略差异。
研究发现,不同冠层高度和耐荫性树种间生态位宽度的差异并不显著。树木的高度决定其在群落垂直空间中的位置,影响种子的传播距离,能够反映植物对光、空间等资源的竞争能力(Hiroseet al., 1987;Poorteret al., 2006; Guoet al., 2013)。喀斯特森林上层往往为高大建群种,对垂直空间资源的竞争能力强,而下层多为小乔木,虽然资源竞争能力低于上层树种,但其种群数量较多,在亚乔木层的水平空间上具有较强竞争性。总的来看,虽然上层、中层和下层树种间生态位宽度逐渐下降,但各功能型对空间资源竞争的分化可能使其未表现出显著差异。树种的耐荫性与冠层高度存在联系,群落上层树种往往为喜光的阳性树种,下层往往为耐荫树种,而中层可能多为中生性树种,因此,不同耐荫性功能型间的生态位宽度也未表现出显著差异。
4.2 喀斯特森林树木功能型的生态位重叠
本研究发现,喀斯特森林树种间的生态位重叠程度普遍较低,重叠值为0~0.2 的占总数的89.10%,表明40 种树木对环境资源的利用和要求差异较大,生态位分化较明显,物种间竞争不激烈,种间关系较稳定。黄甫昭等(2018)对喀斯特季节性雨林优势种群生态位特征的研究同样发现喀斯特森林树种间的生态位重叠指数普遍不高(重叠指数0~0.5 占95.4%)。在喀斯特生境的筛选下,植物种间生态位分化以避免产生激烈竞争(Zhanget al.,2021),此外岩石裸露而形成的空间阻隔可能进一步降低了种间的竞争作用。因此,喀斯特高度异质的生境不仅降低了树种的生态位宽度,同时也明显降低了种间的生态位重叠程度。成熟林中树种的生态位宽度与重叠度普遍较低,这一结果与Fu 等(2015)的研究结果基本一致,表明成熟林中物种生态位趋向高度分化或互补以达到多物种的稳定共存。
生态位重叠体现了物种对同等级资源的利用程度以及空间配置关系(Geangeet al.,2011;董雪等,2020)。有研究认为,生态位宽度大的物种,其资源利用能力较强,分布范围较广,因而与其他物种间的生态位重叠程度也较大,反之亦然(龚雪伟等,2017;Huet al.,2022)。本研究中,常见种、上层种、喜光种以及动物传播种的生态位宽度以及生态位重叠指数均最高,而稀有种、下层种、耐荫种和重力传播种则相反,这一结果进一步证实了上述观点。一般而言,森林乔木上层种多为喜光树种,其对光照、水分、土壤养分等环境资源的需求相似,这种相似的资源利用策略和生境偏好导致树种间生态位重叠较高,而下层种多为耐荫种,在群落中不占优势,其生态位宽度和重叠指数均较低。本研究发现,重力传播树种的生态位重叠指数显著低于动物和风力传播树种。重力传播种往往以母树为中心进行有限距离的扩散,而在喀斯特生境中,短距离扩散不利于种子散布到适合萌发的小生境中,导致其种群更新受阻,而动物和风力传播树种则相反。因而,种子传播方式可能在较大程度上决定了树木对空间资源的占据和利用能力以及种间竞争格局,从而影响树木功能型间的生态位重叠程度。资源量、资源的供求比、资源满足生物需求的程度决定着种间生态位重叠程度,在资源丰富的情况下,即使生态位相似性较高,也不会出现高的生态位重叠(Fuet al.,2015;白晓航等,2017)。本研究中,树木功能型内的生态位重叠指数大多为0~0.2,因此对于成熟的喀斯特森林,同一功能型的树种即使具有相似的资源位,其种间竞争排斥作用也维持在较低水平,这可能是中亚热带喀斯特森林树种高度丰富的潜在原因。
本研究表明,不同树木功能型间的生态位重叠指数存在明显差异。由生态位越宽的树种组成的功能型,其功能型间的生态位重叠指数也越高,即越是空间分布广、资源占据能力强的功能型,其之间的生态位重叠概率越大。因此,当资源受限情况下,越是生活史功能和策略相似的功能型,二者间竞争作用可能越强。如依靠动物和风力传播树种,其种群扩散能力均较强,功能型间的生态位重叠指数较高,因为动物和风力传播树种的种群扩散能力较强,在群落中往往为常见种或偶见种(如黄杞、圆果化香等),同样常见种和偶见种间的生态位重叠指数也较高。
4.3 喀斯特森林的物种共存机制与恢复重建
植物个体差异和生境异质性的相互作用促进了物种共存,在种群扩散率较低时尤为明显,而在异质生境中,斑块质量差异和(或)空间生境自相关的增加,减少了物种间竞争能力的差异,从而允许物种在局域和区域共存(Chenet al.,2020)。群落发生、发展和演替过程既是环境对物种自然选择的过程,也是物种对环境的适应过程,在环境过滤和种间交互作用的多重作用下,通过生态位分化减弱(如生态位重叠度降低)或者避免竞争是实现多物种稳定共存的重要途径(Silvertown,2004;HilleRisLamberset al.,2012;陈磊等,2014)。在喀斯特常绿落叶阔叶混交林中,树种的生态位宽度和重叠程度均较小,表明该群落在局域尺度上树木的生态位分化较为明显,种间竞争强度小。经过长期的生境筛选(如气候、土壤和地形条件等)和种间交互作用(如化感作用、护理效应等),成熟林中多数树种占据着相对独立的资源,种间关系处于协调和平衡的状态,群落处于相对稳定状态。由于岩石裸露而形成的物理空间隔离以及微生境的环境因子差异(Zhanget al.,2013;Wanget al.,2016;Shenet al.,2019),使得分布在特定环境中的物种与其他物种重叠的机会降低,实现多物种的稳定共存。本研究认为,高度异质的喀斯特生境拥有高度丰富的树种组成,生态位分化可能是驱动喀斯特森林群落构建的主要力量,而扩散限制和更新限制也会发挥潜在作用。
近几十年来,通过封山育林和人工造林等措施,我国喀斯特石漠化趋势已得到有效抑制,石漠化面积持续减少(王克林等,2019;Qiaoet al.,2021)。现有的西南喀斯特植被以灌丛、灌草丛等为主,人工介入自然演替进程,可加快形成物种多样的森林群落、提升喀斯特生态系统的功能(王克林等, 2019)。喀斯特森林树种的生态位特征研究成果可为制定适应性的植被恢复与重建措施提供依据(黄甫昭等,2018;李坤等,2020)。以区域性天然林作为种源和植被恢复模板,依据适地适树原则选取合适的造林树种,需要充分考虑树种形态、结构和生理等特征。可依据功能型选取树种,在选取树种时考虑到不同功能型的搭配组合。由于喀斯特生境的高度异质性,在建立人工植被时,要严格根据生境类型筛选适应性强的树种。在植被恢复早期,可选取常见种中的喜光树种,在上层乔木建植后,选取中生或耐荫树种中生态位较宽的树种构建亚乔木层,避开生态位高度重叠的物种,从而建立近自然的空间结构合理的复层林分。此外,尽可能选取动物和风力传播树种,因为这些树种可能具有更好的传播和更新能力。总之,人工干预植被恢复进程中,合理选取和配置不同功能型的树种,有利于充分利用空间资源以及降低种间竞争作用,从而促进喀斯特森林植被快速恢复。
5 结论
中亚热带喀斯特常绿落叶阔叶混交林中,不同树木功能型的生态位宽度和重叠指数普遍较低,表明绝大多数树种占据的生态位空间较小,种间的竞争排斥作用可能较弱,群落处于相对稳定状态。不同树木功能型生态位宽度和重叠指数的差异与生境异质性、树木性状差异和种间关系密切相关,体现了不同功能型对空间资源的占据和利用能力以及竞争作用的差异,这种差异利于局域尺度上喀斯特森林多物种的稳定共存。在进行植被恢复与重建时,需要充分考虑不同植物功能型的生态位差异,合理配置不同功能型树种可能有助于森林的恢复。
