固态共培养木霉和芽孢杆菌抗烟草黑胫病及其活性成分研究
2024-01-30彭治鑫赵栋霖管志坤王耀斌张成省
彭治鑫,赵栋霖,林 伟,高 贵,管志坤,王耀斌,张成省,李 伟
固态共培养木霉和芽孢杆菌抗烟草黑胫病及其活性成分研究
彭治鑫1,2,赵栋霖2,林 伟3*,高 贵4,管志坤5,王耀斌5,张成省2,李 伟1*
(1.长江大学生命科学学院,湖北 荆州 434000;2.中国农业科学院烟草研究所,青岛 266101;3;福建省烟草公司南平市公司,福建 南平 353000;4.贵州省烟草公司黔西南州公司,贵州 兴义 562400;5.山东青岛烟草有限公司,青岛 266000)
为提高棘孢木霉HG1和枯草芽孢杆菌tpb55共培养菌的生物量和抗烟草疫霉活性,构建了两株菌固态共培养体系,并探究了其活性成分。采用菌丝生长速率法评价发酵提取物的抑菌活性,结合发酵后菌量,确定固态共培养的条件;运用正/反硅胶柱层析对活性成分进行追踪分离,使用高效液相色谱(HPLC)和核磁共振(NMR)及ESI质谱,纯化和鉴定活性化合物;通过盆栽试验验证共培养发酵对烟草黑胫病的防治效果。结果表明,两株生防菌固态共培养发酵的初始条件为:枯草芽孢杆菌(106CFU/mL,接种量3%)接种12 h后再接种棘孢木霉(106CFU/mL,接种量1.5%);发酵14 d时,枯草芽孢杆菌菌量达到1.58×1010CFU/g,棘孢木霉菌量达到6.75×1010CFU/g,并且提取物的抗菌活性最强,抑制率达到73.25%;从活性组分中分离得到一个单体化合物,鉴定为harziandione。盆栽防病效果表明,采用固态培养基共培养发酵的产物对烟草黑胫病的防效优于单培养,并能够改善土壤理化性质。该研究为烟草黑胫病生防菌剂的研发提供了新的思路和方法。
固态发酵;共培养;抗卵菌活性;harziandione
烟草疫霉是引起烟草黑胫病的主要病原菌,对烟草生产危害严重[1]。目前,烟草黑胫病的防治依然是以化学防治为主[2],亟需加强生物防治方法和产品的研究与开发。生防微生物通过产生抑菌物质、与病原菌竞争营养和空间、寄生在病原菌内或使植物产生诱抗作用来发挥防病和抗病作用,具有无毒害、无残留、安全环保等优点[3],已经成为烟草病害生物防治的重要方法和手段。
木霉和芽孢杆菌作为应用最为广泛、效果最为显著的生防菌,对多种植物病原菌具有明显的抑制作用。然而,单一菌种的生防效果不稳定,采用不同生防菌株进行共培养发酵,可以提高优势菌株生存及生防效果[4]。本实验室前期构建了棘孢木霉HG1和枯草芽孢杆菌tpb55液体共培养发酵体系[5],对烟草黑胫病具有较好防治效果,然而发酵后菌量和代谢产物产量较低,抗菌活性有待提升。
目前文献报道的共培养菌剂的发酵主要为液态发酵,但液体发酵保存期短,不利于储存和长距离运输,在生产中会排放大量的污水。固态发酵具有产量高、产物稳定性好、生产成本低、后期处理简单、污染少等优点[6]。与液体发酵相比,固态发酵含水量较少,养分和代谢产物呈现明显的分层现象,从而能够更好刺激微生物持续发酵。另外,在固态发酵中,代谢产物对菌株的抑制效果较小,发酵产物的终浓度高,能够产生更多的次级代谢产物[7-8]。Ola等[9]将植物内生尖孢镰刀菌和枯草芽孢杆菌在大米培养基上共培养,产生了4种单培养时没有的化合物,并且共培养后,代谢物产量与单培养相比提高了78倍。段金柱等[10]在绿色木霉产纤维素酶的发酵研究中发现固体发酵产率(742~827 U/g)比液体发酵产率(640~663 U/g)高25%,并且能够产生更多的活性酶组分。
为了建立高效和抑菌活性更强的共培养发酵体系,本研究采用固态发酵方法对棘孢木霉HG1和枯草芽孢杆菌tpb55进行共培养,以发酵后两株生防菌的菌量和提取物对烟草疫霉的抑制率为衡量标准,确定最优固态发酵条件;在此基础上,开展抑菌组分及活性成分的追踪分离;最后,通过菌株发酵的固体培养基防治烟草黑胫病的盆栽试验,评估土壤理化性质,验证其防病效果。
1 材料与方法
1.1 菌株来源
棘孢木霉HG1分离自海南红树林样品[5],在马铃薯葡萄糖琼脂(PDA)培养基上28 ℃培养;枯草芽孢杆菌tpb55分离自烟草叶面[11],在营养琼脂(NA)培养基上28 ℃培养。病原菌为烟草疫霉JM01[12],分离自烟草黑胫病株,在燕麦琼脂(OA)培养基上28 ℃培养。以上菌株采用甘油–80 ℃保存于中国农业科学院烟草研究所海洋农业研究中心。
1.2 主要试剂和仪器
1.2.1 主要培养基和试剂 马铃薯葡萄糖琼脂培养基(PDA)、营养肉汤培养基(NB)、营养琼脂培养基(NA)、燕麦培养基(OA),购自北京索莱宝科技有限公司。甲醇为分析纯和色谱纯,二氯甲烷、乙酸乙酯和石油醚均为分析纯,购自中国国药集团化学试剂公司。硅胶(100~200目),购自中国青岛海洋化工厂;反向硅胶,购自德国EMD Millipore公司;Sephadex LH-20凝胶,购自美国GE Healthcare公司;GF254硅胶板,购自中国烟台江友硅胶开发有限公司。
1.2.2 主要仪器 CX23显微镜(日本Olympus);LRH-250生化培养箱(上海一恒);ZQZY-CF9.9振荡培养箱(上海知楚);Hei-VAP Value旋转蒸发仪(德国Heidolph);Waters e2695高效液相色谱仪(美国Waters);DD2 500 MHz NMR核磁共振波谱仪(美国Agilent);ESI-MS Micromass Q-TOF质谱仪(美国Waters)。
1.3 棘孢木霉HG1和枯草芽孢杆菌tpb55共培养接种比例的优化
1.3.1 固态培养基及种子液的制备 固态培养基制备:1000 mL锥形瓶中加入90 g大米和110 mL蒸馏水,121 ℃高温灭菌20 min。
棘孢木霉HG1种子液制备:棘孢木霉在PDA平板上28 ℃培养10 d。无菌水冲洗,使用纱布过滤菌丝,吸取上清,用血细胞计数板在显微镜下确定棘孢木霉孢子液浓度。无菌水稀释成1×106CFU/mL孢子悬浮液备用。
枯草芽孢杆菌tpb55种子液制备:在NA平板上划线,于28 ℃培养12 h,挑取单菌落于NB液体培养基里,180 r/min,28 ℃培养12 h。测菌液600值,无菌水稀释成1×106CFU/mL菌悬液备用。
1.3.2 棘孢木霉HG1和枯草芽孢杆菌tpb55共培养接种比例的确定 设置枯草芽孢杆菌tpb55单培养接种1.5 mL种子液,棘孢木霉HG1单培养接种1.5 mL种子液,枯草芽孢杆菌tpb55和棘孢木霉HG1按1∶1,1∶2,2∶1,1∶10,10∶1比例共培养接种,加入到大米培养基中后搅拌使其混合。接种完成后,于25 ℃静置培养30 d。
取1 g培养基,用1 mL无菌水稀释成10-3、10-4、10-5、10-6不同梯度的菌液。取100 μL的菌液分别在含链霉素和氯霉素(10 mg/mL)的PDA固体平板,含咪酰胺(10 mg/mL)的NA固体平板上涂板,在28 ℃培养1~2 d,统计单菌落的数量,计算菌量。
静置培养30 d后,大米培养基先使用乙酸乙酯超声浸取两遍,浸取的乙酸乙酯旋蒸浓缩至干。再使用甲醇和二氯甲烷(1∶1)超声浸取两遍,浸取液先旋蒸浓缩后用水转出,在水相中加入等体积的乙酸乙酯萃取3次,旋蒸至干。两次蒸干的乙酸乙酯相,合并后得到提取物。
1.3.3 共培养提取物对烟草疫霉的抑制率分析 采用菌丝生长速率法[5]测定提取物对烟草疫霉的抑菌率:取30 mg提取物加入150 μL二甲亚砜(DMSO)溶解,配置成200 mg/mL的溶液,取50 μL溶液加入到10 mL的燕麦培养基中,配成终浓度1 mg/mL的含药培养基,倒入6 cm的培养皿中制成平板培养基。用打孔器从活化7 d的烟草疫霉平板上取5 mm的菌碟,倒置在平板中央,28 ℃培养36 h。采用十字交叉法测量菌丝生长的直径,以只加入DMSO的燕麦培养基作空白对照。每处理重复3次。
菌落直径(cm)=测量菌落直径−0.5 (1)
抑菌率(%)=(对照菌落直径−处理菌落直径)/对照菌落直径×100 (2)
1.4 棘孢木霉HG1和枯草芽孢杆菌tpb55共培养接种的顺序和时间优化
按照优化后的接种比例,设置枯草芽孢杆菌tpb55和棘孢木霉HG1不同接种顺序和时间:枯草芽孢杆菌tpb55单培养(t);棘孢木霉HG1单培养(h);枯草芽孢杆菌tpb55和棘孢木霉HG1同时接种(th);其他共培养分别在先接种枯草芽孢杆菌tpb55 12 h(th12)、24 h(th24)、36 h(th36)后再接棘孢木霉HG1;先接棘孢木霉HG1,24 h(ht24)、36 h(ht36)、48 h(ht48)后再接枯草芽孢杆菌tpb55。在发酵14 d时涂板,统计菌量,提取代谢产物。将各个提取物经过减压硅胶柱色谱,乙酸乙酯/石油醚(10%),乙酸乙酯(100%),甲醇/水(10%)3个梯度进行洗脱,采用菌丝生长速率法测定各个组分对烟草疫霉的抑制率。
1.5 共培养固态发酵活性次级代谢产物的分离
根据优化后的发酵条件,发酵10瓶,提取代谢产物。将提取物经过减压硅胶柱层析,以乙酸乙酯/石油醚(5%~100%)和甲醇/乙酸乙酯(10%~50%)进行梯度洗脱。通过薄层层析分析,合并成5个组分,采用菌丝生长速率法测定各组分在1.0 mg/mL浓度下对烟草疫霉的抑菌率。将抑菌率最高的组分经过反向硅胶柱层析分离,以甲醇/水(30%~100%)进行梯度洗脱,再通过高效液相色谱分离纯化,采用1H和13C NMR及ESI质谱鉴定化合物结构。采用菌丝生长速率法测定化合物在0.1 mg/mL浓度下对烟草疫霉的抑菌率。
1.6 盆栽试验
供试土壤来自中国农业科学院烟草研究所即墨试验基地(田间自然土∶灭菌基质土=7∶3)[13]。将土壤与疫霉菌谷(1.2 g菌谷∶180 g土壤)混匀作为防病盆栽试验的病土,将不同发酵的大米固体培养基与病土(3 g∶180 g)混匀作为处理。盆栽试验总共设置3个处理:枯草芽孢杆菌tpb55单培养(t)、棘孢木霉HG1单培养(h)、枯草芽孢杆菌tpb55接种12 h后接种棘孢木霉HG1(th12),以只加入疫霉菌谷的病土作对照。取长势一致的烟苗移入土中,设置3个重复,每个处理30株烟苗。移栽7 d后统计发病率和病情指数,计算防病效果。烟草黑胫病的病情分级参照中华人民共和国烟草行业标准(GB/T 23222—2008)进行统计[14]。
1.7 盆栽土壤理化性质测定
盆栽试验第14天,取盆栽土。自然风干后,通过20 mm筛,测定土壤理化性质,包括pH、电导率、有机质含量(重铬酸钾容量法)、速效钾含量(NH4OAc浸提-火焰光度计法)、铵态氮和磷酸根含量(SKALAR SAN+连续流动注射分析)[15]。
1.8 数据分析
使用Microsoft Office Excel 2019和SPSS 18.0进行数据统计分析,Adobe Photoshop 2020作图。
2 结 果
2.1 共培养接种比例的确定
由表1可知,当枯草芽孢杆菌tpb55和棘孢木霉HG1接种比例为2∶1时,枯草芽孢杆菌tpb55(5.5×106CFU/g)和棘孢木霉HG1(2.65×107CFU/g)菌量高于其他接种比例。但值得注意的是,共培养后枯草芽孢杆菌菌量显著低于单培养,原因可能是为了得到更丰富的次级代谢产物,发酵时间过长,部分枯草芽孢杆菌已死亡,并且木霉的代谢产物对枯草芽孢杆菌产生了抑制作用。因此,后续将发酵时间缩短,但考虑到真菌产生次级代谢产物需要的时间较长,发酵时间确定为14 d[16]。

表1 不同接种比例HG1和tpb55在发酵30 d的菌量
注:表中t,h分别表示枯草芽孢杆菌tpb55、棘孢木霉HG1的单培养,1∶1、1∶2、2∶1、1∶10、10∶1是枯草芽孢杆菌tpb55与棘孢木霉HG1的接种比例。不同小写字母代表处理间<0.05水平差异显著,下同。
Note: In the table, t and h represent the single culture oftpb55 andHG1, respectively.1∶1, 1∶2, 2∶1, 1∶10 and 10∶1 are inoculation ratios oftpb55 toHG1. There was significant difference in the level of<0.05 between different lowercase letters, the same below.
不同接种比例下的提取物对烟草疫霉抑菌率,如表2所示,当枯草芽孢杆菌tpb55和棘孢木霉HG1接种比例为2∶1时,提取物的抑菌率(46.17%)最高。因此,综合发酵后两株菌菌量和提取物的抑菌活性,选用枯草芽孢杆菌tpb55和棘孢木霉HG1的接种比例为2∶1(枯草芽孢杆菌tpb55接种量为培养基体积的3%,棘孢木霉HG1接种量为培养基体积的1.5%),接种种子液浓度为1×106CFU/mL。
2.2 共培养接种顺序和时间的确定
在枯草芽孢杆菌tpb55和棘孢木霉HG1接种比例2∶1下,对枯草芽孢杆菌tpb55和棘孢木霉HG1的接种顺序和时间进行优化。如表3所示,在枯草芽孢杆菌tpb55先接种12 h后再接种棘孢木霉HG1,枯草芽孢杆菌tpb55(1.58×1010CFU/g)和棘孢木霉HG1(6.75×1010CFU/g)的菌量最多。且棘孢木霉HG1菌量显著高于单培养。与发酵30 d相比,两株生防菌菌量均得到了明显的提升。如表4所示,100%乙酸乙酯提取的组分抑菌活性最高,而10%乙酸乙酯组分和10%甲醇组分活性较低,说明抑菌活性成分主要集中在100%乙酸乙酯组分中。枯草芽孢杆菌tpb55先接种12 h后再接种棘孢木霉HG1时,提取物的抑菌率(73.25%)显著高于其他处理。因此,共培养选用的接种顺序和时间为先接种枯草芽孢杆菌tpb55,12 h后再接种棘孢木霉HG1。
综上所述,固态共培养发酵条件:在枯草芽孢杆菌tpb55接种12 h后接种棘孢木霉HG1,枯草芽孢杆菌tpb55(106CFU/mL)接种量为培养基体积的3%,棘孢木霉HG1(106CFU/mL)接种量为培养基体积的1.5%。
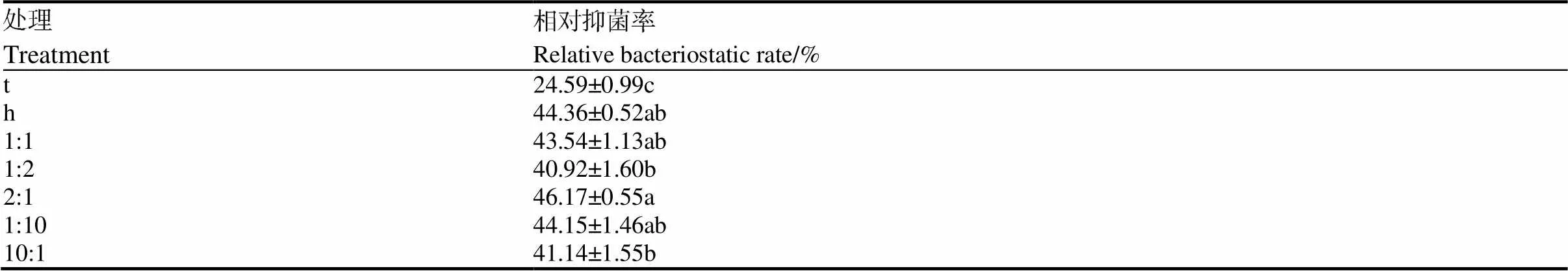
表2 不同接种比例HG1和tpb55提取物对烟草疫霉的抑菌率

表3 HG1和tpb55在不同接种时间发酵14 d的菌量

表4 不同接种时间的提取物组分的减压组分对烟草疫霉的抑制率
2.3 抑菌活性成分的分离及结构鉴定
静置培养14 d后提取发酵产物,共得到提取物2.3 g。测定5个减压组分对烟草疫霉的抑菌率,如表5和图1所示,ht2的抑菌活性(74.69%)显著高于其他组分,ht2组分经过反向硅胶柱色谱和高效液相色谱分离纯化得到的化合物1,鉴定为萜类化合物harziandione,结构如图2所示。在100 μg/mL浓度下,harziandione对烟草疫霉的抑制率为19.8%。
harziandione的核磁数据:1H-NMR(500 MHz,CDCl3):H2.91(2H,overlapped,H-4b,5),2.56(1H,d,=13.5 Hz,H-12b),2.46(3H,overlapped,H-8b,12a,14),2.26(1H,d,=7.0 Hz,H-2),2.12(3H,s,H-20),2.05(3H,overlapped,H-4a,8a,15b),1.90(1H,dd,=10.5,5.0 Hz,H-7b),1.53(1H,t,=7.5 Hz,H-15a),1.51(3H,s,H-19),1.41(1H,t,=11.0 Hz,H-7a),1.11(3H,d,=6.0 Hz,H-18),1.00(3H,s,H-17),0.98(3H,s,H-16);13C-NMR(125 MHz,CDCl3):C214.4(C-3),197.9(C-11),149.4(C-9),146.4(C-10),59.9(C-12),59.3(C-2),53.0(C-14),51.6(C-6),49.5(C-1),42.6(C-4),39.9(C-13),29.9(C-5),29.8(C-7),29.5(C-8),26.6(C-15),25.0(C-16),23.2(C-17),22.5(C-20),20.9(C-18),20.7(C-19);ESIMS:/302.31[M+2H]+。
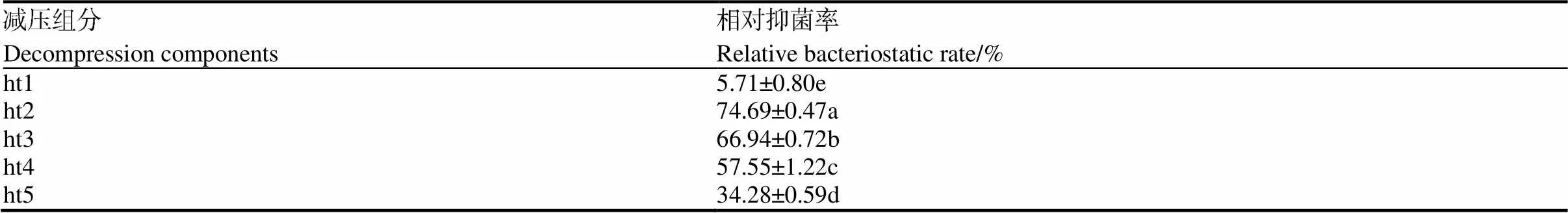
表5 提取物减压组分对烟草疫霉的抑制率

图1 提取物减压组分对烟草疫霉的抑制活性

图2 harziandione结构及对烟草疫霉的抑制率
2.4 盆栽试验及土壤理化性质
如表6所示,在处理后第7天,处理组的病情指数较对照组相比均显著降低。80%的对照组烟苗在7 d时茎部发软变黑、枯萎和死亡,而70%~80%的处理组烟苗依旧健康。其中棘孢木霉HG1与枯草芽孢杆菌tpb55共培养的防治效果优于棘孢木霉HG1、枯草芽孢杆菌tpb55单培养,其病情指数为27.78,防治效果为61.94%。
土壤理化试验表明(表7),施加固态共培养棘孢木霉HG1与枯草芽孢杆菌tpb55后,土壤理化性质有所改善。处理14 d后,处理组的盆栽土壤pH相较对照组显著提升,而电导率则比对照组土样有所下降。此外,与对照组相比,处理组的土壤中硝态氮和磷酸根含量得到提升,说明生防菌的施加可提升土壤中微量元素的含量,有利于植物的生长和对病原抵抗力的增加。
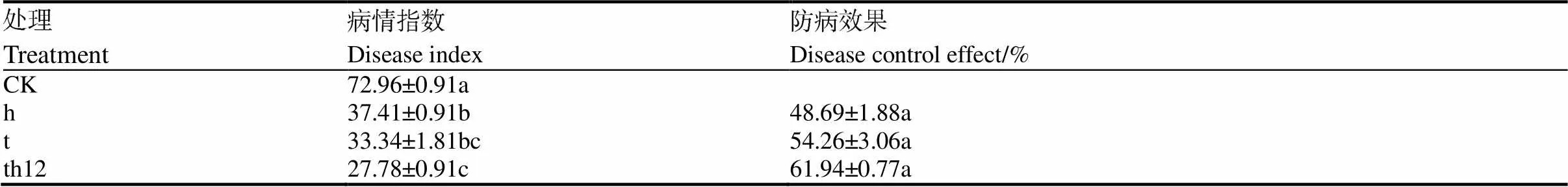
表6 盆栽试验7 d时的病情指数和防治效果

表7 土壤理化性质
3 讨 论
本实验室前期采用液体培养基对两株菌进行发酵,但发酵后两株菌菌量较低,在发酵14 d时,枯草芽孢杆菌tpb55最大菌量为1.2×108CFU/mL、棘孢木霉HG1最大菌量为2.34×106CFU/mL。在木霉和芽孢杆菌共培养中,木霉一般起主导作用[17]。目前菌株的共培养发酵主要是液体发酵,但在液体培养基中真菌产孢困难。与液体培养基相比,固体培养基更接近木霉生长环境,木霉能够在固体培养基表面和内部产孢,提高孢子产量[18]。段婉冬[19]采用甜叶菊渣固体培养基发酵木霉,培养4~6 d,木霉的孢子数量能达到7×109CFU/g。本研究采用大米培养基进行固态发酵,增加了两株生防菌共培养后的菌量,枯草芽孢杆菌tpb55达到1.58×1010CFU/g,棘孢木霉HG1达到6.75×1010CFU/g。
目前尚未有枯草芽孢杆菌和棘孢木霉固态共培养发酵的文献报道。不同菌株的最佳固体发酵条件不同,使用不同菌株进行共培养时,需优化明确最佳共培养发酵条件。CHEN等[20]确定了当使用黄孢原毛平革菌与绿色木霉对秸秆进行固态发酵降解时,两株菌的最佳接种比例为1∶1。杨壮等[21]在对乳杆菌进行固态发酵时确定其最佳的接种量为4.80%,最佳的发酵时间为51.30 h,随发酵时间延长黄精多糖的得率反而降低。王小兵等[22]发现短小芽胞杆菌和解淀粉芽胞杆菌共培养发酵16 h时,菌量达到峰值。本研究发现,当枯草芽孢杆菌tpb55和棘孢木霉HG1的接种比例为2∶1时,提取物具有更高的抑菌活性,能够促进更多的抗烟草疫霉物质的产生;枯草芽孢杆菌tpb55接种12 h后接种棘孢木霉HG1发酵14 d时菌量最多,tpb55和HG1的菌量分别为1.58×1010、6.75×1010CFU/g,效果优于共培养发酵30 d。
Harziane型二萜化合物是由GHISALBERTI等[23]于1992年从哈茨木霉中发现的一类化学分子,它含有独特的四环6/5/4/7碳骨架和5~6个相邻的立体中心。目前已知的harziane型二萜化合物几乎都是从木霉中分离发现的,但其对病原卵菌的抑制作用尚无相关报道。Li等[24]从深海木霉属真菌的发酵产物中分离鉴定出7种harziane型二萜,发现在C-8,C-18位点上有羟基的二萜能够抑制NO的生成,具有抗炎症的作用;而这7种二萜类化合物对常见的植物病原真菌均没有明显的抑菌活性。本研究采用活性追踪的方法从枯草芽孢杆菌tpb55和棘孢木霉HG1固态共培养的提取物中分离得到一个二萜类化合物harziandione,在100 μg/mL浓度下,对烟草疫霉的抑菌活性为19.8%,具有一定的抑菌活性。但与组分ht2的抗菌活性(74.69%)相比,harziandione的活性不强。原因可能是单体化合物产量较低,使用的测试浓度较低,因此活性没有达到预期,同时,活性组分中的不同化合物可能具有协同作用,这需要后续的深入研究。
4 结 论
本试验筛选出的棘孢木霉HG1与枯草枯草芽孢杆菌tpb55最佳的固态共发酵培养条件为枯草芽孢杆菌tpb55接种12 h后接种棘孢木霉HG1,枯草芽孢杆菌tpb55(106CFU/mL)接种量为3%,棘孢木霉HG1(106CFU/mL)接种量为1.5%。从共培养的发酵提取物中分离得到单体化合物harziandione,在100 μg/mL时对烟草疫霉的抑菌活性为19.8%。采用固态共培养发酵对烟草黑胫病的盆栽防治效果为61.94%,优于单培养。同时能够改善土壤pH,提高土壤中微量元素含量。本研究构建的枯草芽孢杆菌和棘孢木霉固态共培养体系,具有开发成为新型抗烟草疫霉生防菌剂的潜力。
[1] GUO D, YUAN C, LUO Y, et al. Biocontrol of tobacco black shank disease () byBa168[J]. Pestic Biochem Physiol, 2020, 165: 104523.
[2] 彭丽娟,丁海霞,陆铮铮. 贵州烟草黑胫病菌对甲霜灵的抗药性测定[J]. 贵州农业科学,2014,42(8):92-95.
PENG L J, DING H X, LU Z Z. Drug resistance ofagainst metalaxyi (WP) in Guizhou[J]. Guizhou Agricultural Sciences, 2014, 42(8): 92-95.
[3] LA SPADA F, STRACQUADANIO C, RIOLO M, et al. Trichoderma counteracts the challenge ofinfections on tomato by modulating plant defense mechanisms and the expression of crinkler, necrosis-inducingprotein 1, and cellulose-binding elicitor lectin pathogenic effectors[J]. Front Plant Sci, 2020, 11: 583539.
[4] MA Q, CONG Y, FENG L, et al. Effects of mixed culture fermentation ofandon its constituent strains and the biocontrol of tomato Fusarium wilt[J]. J Appl Microbiol, 2022, 132(1): 532-546.
[5] 张希芬,林伟,李清钰,等. 抗烟草疫霉活性木霉与芽孢杆菌共培养体系的构建与优化[J]. 中国烟草科学,2022,43(1):61-68.
ZHANG X F, LIN W, LI Q Y, et al. Construction and optimization of a co-culture system of anti-HG1 andTpb55[J]. Chinese Tobacco Science, 2022, 43(1): 61-68.
[6] 唐堂,王琪,周卫强,等. 固态发酵技术研究分析及应用[J]. 工业微生物,2020,50(5):55-59.
TANG T, WANG Q, ZHOU W Q, et al. Solid-state fermentation technology research analysis and application[J]. Industrial Microorganisms, 2020, 50(5): 55-59.
[7] LOUHASAKUI Y, WADO H, LATEH R, et al. Solid-state fermentation of Saba banana peel for pigment production by[J]. Braz J Microbiol, 2023, 54(1): 93-102.
[8] RAHARDIO YS, TRAMPER J, RINZEMA A. Modeling conversion and transport phenomena in solid-state fermentation: a review and perspectives[J]. Biotechnol Adv, 2006, 24(2): 161-179.
[9] OLA A R, THOMY D, LAI D, et al. Inducing secondary metabolite production by the endophytic fungusthrough coculture with[J]. J Nat Prod, 2013, 76(11):2094-2099.
[10] 段金柱,曹淡君. 固体发酵与液体发酵生产纤维素酶产率与催化性能比较[J]. 粮食与饲料工业,2000(3):24-26.
DUAN J Z, CAO D J. The compare of SSF and LSF with trichodema viride for cellulase production in yield and catalysis property[J]. Grain and Feed Industry, 2000(3): 24-26.
[11] 张成省,孔凡玉,关小红,等. 烟草叶围细菌Tpb55菌株的鉴定及其抑菌活性[J]. 中国生物防治,2008(1):63-68.
ZHANG C S, KONG F Y, GUAN X H, et al. Identification and antagonistic activity of tobacco phyllosphereTpb55[J]. Chinese Journal of Biological Control, 2008(1): 63-68.
[12] ZHANG C S, GAO J M, HAN T et al. Integrated control of tobacco black shank by combined use of riboflavin andstrain Tpb55[J]. Bio Control, 2017, 62(6): 835-845.
[13] 王耀晨. 花椒籽炭基有机肥防治烟草黑胫病研究[D]. 北京:中国农业科学院,2020.
WANG Y C. Study on control of tobacco black shank disease with biochar-based fertilizer of prickly ash seed[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020.
[14] 国家烟草专卖局. 烟草病虫害分级调查方法:GB/T 23222—2008[S]. 北京:中国标准出版社,2008.
State Tobacco Monopoly Administration. Grade and investigation method of tobacco diseases and insect pests: GB/T 23222—2008[S]. Beijing: Standards Press of China, 2008.
[15] 鲍士旦. 土壤农化分析[M]. 北京:中国农业出版社,2000.
BAO S D. Soil agrochemical analysis[M]. Beijing: China Agricultural Publishing House, 2000.
[16] 鲍泽安. 海洋真菌sp. S212B的次级代谢产物和生物活性研究[D]. 咸阳:西北农林科技大学,2022.
BAO Z A. Studies on chemical constituents and bioactivities of marine fungussp. S212B[D]. Xianyang: Northwest A&F University, 2022.
[17] KARUPPIAH V, LU Z X, LIU H, et al. Co-cultivation of T. asperellum GDFS1009 and B. amyloliquefaciens 1841: Strategy to regulate the production of ligno-cellulolytic enzymes for the lignocellulose biomass degradation[J]. J Environ Manage, 2022, 301: 113833.
[18] NEE, NIGAM P S, PANDEY A. Solid-state fermentation technology for bioconversion of biomass and agricultural residues[J]. Biotechnology for Agro-industrial Residues Utilisation, 2009, 197-221.
[19] 段婉冬. 甜叶菊渣固体发酵木霉NJAU 4742及发酵菌剂对玉米苗期生长的影响[D]. 南京:南京农业大学,2020.
DUAN W D. Solid state fermentation ofNJAU 4742 with stevia residue ang its effects on corn growth in seeding stage[D]. Nanjing: Nanjing Agricultural University, 2020.
[20] CHEN K J, TANG J C, XU B H, et al. Degradation enhancement of rice straw by co-culture ofand[J]. Sci Rep, 2019, 9(1): 19708.
[21] 杨壮,刘怡琳,李隆熙,等. 固态发酵制备黄精多糖的工艺优化、理化特性及抗氧化活性[J]. 食品与发酵工业,2023(7):1-9.
YANG Z, LIU Y L, LI L X, et al. Process optimization, physicochemical properties and antioxidant activity of polygonatum sibiricum polysaccharide prepared by solid-state fermentation[J]. Food and Fermentation Industry, 2023(7): 1-9.
[22] 王小兵,付宽宽,顾红亚,等. 复合芽胞杆菌共培养条件的响应面优化研究[J]. 江西农业学报,2021,33(12):1-7.
WANG X B, FU K K, GU H Y et al. Optimization of co-culture conditions of compoundby response surface methodology[J]. Acta Agriculture Jiangxi, 2021, 33(12): 1-7.
[23] GHISALBERTI E L, HOCKLESS D C R, ROWLAND C, et al. Harziandione, a new class of diterpene from[J]. J. Nat. Prod, 1992, 55: 1690-1694.
[24] LI H X, LIU X, LI X, et al. Novel Harziane diterpenes from deep-sea sediment fungussp. SCSIOW21 and their potential anti-inflammatory effects[J]. Mar Drugs, 2021, 19(12): 689.
Study on Co-culture ofandby Solid-state Fermentation against Tobacco Black Shank Disease and Their Bioactive Secondary Metabolites
PENG Zhixin1,2, ZHAO Donglin2, LIN Wei3*, GAO Gui4,GUAN Zhikun5, WANG Yaobin5, ZHANG Chengsheng2, LI Wei1*
(1. College of Life Science, Yangtze University, Jingzhou, Hubei 434000, China; 2. Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 3. Nanping Branch, Fujian Tobacco Company, Nanping, Fujian 353000, China; 4. Qianxinan Prefecture Branch, Guizhou Tobacco Company, Xingyi, Guizhou 562400, China; 5. Shandong Qingdao Tobacco Co., Ltd., Qingdao 266000, China)
In order to improve the biomass ofHG1 andtpb55 in the co-culture system, and increase their anti-oomycetes activities against, and to explore the bioactive ingredients, the solid-state fermentation system was constructed. The anti-oomycetes activities were evaluated by the mycelium growth rate method. The solid-state fermentation conditions were determined by the anti-oomycetes activities and biomass of HG1 and tpb55. The bioactivity guided separation and isolation of fermentation extracts by silica gel and octadecylsilyl silica gel (ODS) column chromatography (CC), as well as high performance liquid chromatography (HPLC), together with NMR and ESI mass spectrometry were applied to clarify and identify the bioactive ingredients. Finally, pot experiments were applied to verify the control effect of solid-state co-culture of HG1 and tpb55 on tobacco black shank. The results showed that the optimal solid-state fermentation condition was as follows: the inoculation sequence was first(106CFU/mL, 3%) 12 h and then inoculated with(106CFU/mL, 1.5%). After 14 d of fermentation, under the optimal condition, the bacterial amounts of tpb55 were 1.58×1010CFU/g and the spore amounts of HG1 were 6.75×1010CFU/g, with the strongest anti-oomycete activity towards(inhibition rate 73.25%). A pure compound was isolated from the bioactive fraction and identified as harziandione. The results of pot experiments showed that the control effect of solid co-culture strains on tobacco black shank was significantly better than that of single culture, reached 61.94%. The co-cultured strains could also improve soil physical and chemical properties. This study provided a new idea and method for the research and development of co-culture biocontrol agents for tobacco black shank disease.
solid state fermentation; co-culture; anti-oomycetes activity; harziandione
S435.72
A
1007-5119(2023)06-0036-08
福建省烟草公司南平市公司科技项目(NYK2021-01-03);中国烟草总公司黔西南州公司科技项目(2022-07);山东省烟草公司科技项目(201837020027209);青岛市科技惠民示范专项(23-3-8-xdny-2-nsh)
彭治鑫(1999-),男,在读硕士研究生,主要从事植物病害生物防治研究。E-mail:P18671073969@163.com*通信作者。E-mail:李 伟,wetli@yangtzeu.edu.cn;林 伟,lw160718@caas.cn
2023-07-25
2023-11-14
10.13496/j.issn.1007-5119.2023.06.006
