花生粕的饲用资源开发及在养殖业的应用
2024-01-24崔艳红韩庆功刘长忠
■ 崔艳红 韩庆功 何 云 刘长忠
(河南科技学院,河南新乡 453003)
养殖业的发展使得饲料原料的需求量激增,我国 工业饲料总产量在2015 年突破了2 亿吨,至2022 年突破了3 亿吨,达到了30 223.4 万吨,比上年增长3.0%[1]。我国养殖者自行配制饲料也占有相当大的比例,因此每年养殖业消耗粮油饲料资源实际达到3.5 亿吨以上,但近五年我国粮食总产量每年稳定在6.5亿吨左右[2],造成我国饲料粮资源严重短缺,尤其是蛋白质饲料资源缺口严重。中国海关总署统计数据显示,2020 年我国进口大豆10 033 万吨,2021 年进口略有下降,为9 652万吨;2020年我国进口鱼粉142万吨,2021 年进口鱼粉182 万吨,同比增加28.34%,创历史新高。豆粕、鱼粉等优质蛋白饲料资源严重短缺,已成为限制畜牧业发展的主要因素,开发新的非常规蛋白质饲料资源、优化饲料配方设计、寻求豆粕减量替代产品等已成亟待解决的问题。为维持动物正常生产用饲料原料,确保动物源性产品的稳定供给,2021年中央一号文件提出要“稳定大豆生产,多措并举发展油菜、花生等油料作物”,以提高蛋白质原料资源的自给能力,减少进口依赖。同时2021年3月农业农村部畜牧兽医局发布了“玉米豆粕减量替代”工作的通知,制定了“饲料中玉米豆粕减量替代工作方案”,2023年4 月农业农村部办公厅再次制定印发了《饲用豆粕减量替代三年行动方案》[3],替代方案的发布为开辟新的饲料资源、节约粮食、缓解饲料资源紧缺提供了政策支持和技术指导。
花生粕中营养物质丰富,但优质蛋白质不足,跟鱼粉等动物蛋白相比,蛋白质的可消化吸收率较低。另外,花生粕中氨基酸组成不平衡,必需氨基酸中的赖氨酸含量只有豆粕的50%,精氨酸与赖氨酸比例严重失衡,比值为3~4,缺乏赖氨酸、蛋氨酸等[4],因此蛋白质品质欠佳,有效利用率低。花生粕极易感染黄曲霉、寄生曲霉等而产生黄曲霉毒素[5-6],在2017 年至2021 年的霉菌毒素调查中发现,抽查的69 个花生粕样本黄曲霉毒素检出率100%,正常情况下,黄曲霉毒素总量为2~50 μg/kg,但自然污染较严重的黄曲霉毒素平均水平达417.72 μg/kg[7]。王石等[8]通过建立的全自动免疫亲和柱净化-液相色谱串联质谱法检测9 个花生粕样本,黄曲霉毒毒素含量最高可达到412.5 μg/kg。花生粕中含有胰蛋白酶抑制因子、致甲状腺肿素和植酸等多种抗营养因子[9],其中以植酸为主,蛋白质、脂质、淀粉以及钙、锌等矿物质可能与植酸盐螯合,从而直接影响单胃动物对营养物质的消化率[10-11]。花生粕贮存期间易被病原微生物污染,尤其在潮湿的环境中,易被大肠杆菌、沙门氏菌、青霉、黄曲霉等病原菌污染[12],动物食用污染的花生粕会引起一系列肠道疾病,严重影响畜禽的生长发育,甚至畜禽的死亡。花生油提取主要采用热榨工艺,在高温条件下,蛋白质变性度高,有的甚至烧焦炭化,花生蛋白的持水性、乳化性、溶解度下降[13-14],导致蛋白质和氨基酸的利用率进一步降低。由于花生粕在营养学上的品质缺陷限制了其在动物生产中的应用。
改善花生粕品质的方法主要有物理、化学和生物发酵法,其中生物发酵法是最有应用前景的有效途径之一。花生粕发酵过程中微生物利用自身分泌的酶系降解其中的营养或抗营养成分,将产酶和酶解紧密连接,不仅可以改善发酵底物的质地和适口性,并可产生功能性多肽、多糖和积累有益代谢产物,可有效去除花生粕中的抗营养因子和毒素,从而提高原料的利用率,减少粪便中氮、磷、铜等元素在环境中排放。此外,发酵花生粕中的益生菌可以调节动物肠道微生物群落结构,抑制病原菌的繁殖或定植,提高肠黏膜免疫,减少腹泻和促进生长等。本文介绍了当前优质蛋白质饲料资源紧缺和替代资源花生粕营养学上的品质缺陷,然后对花生粕资源状况、营养特性、生物发酵花生粕研究现状等进行简述,最后对其在猪禽、反刍动物和水产动物养殖中的应用进行综述,以期为缓解蛋白质原料紧张、提高花生粕的附加值和安全高效可持续利用提供参考依据。
1 花生粕资源及营养特性
1.1 花生粕原料现状
中国是重要的花生生产国,花生是我国五大油料作物之一,2016年我国花生总产量为1 669万吨,2019年总产量为1 752万吨,2022年总产量达1 830万吨,花生产量保持稳步增长。我国花生种植面积居世界第二位,花生产量居世界第一位。当前我国花生产量仅次于大豆,居第二位。我国花生主要用于榨油,占比约在60%左右[15],花生粕是花生经压榨制油后的副产物,因此我国花生饼粕原料资源丰富,年产总量在550 万吨以上。河南省是我国花生的主产区,2020 年河南省花生种植面积增加3.873 万公顷,达到了126.184万公顷,花生产量594.93万吨,增加18.21万吨,增长3.2%。2022年河南省花生种植面积128.712万公顷,花生产量615.41 万吨,种植面积和产量均居全国第一[16],因此河南省做好花生副产物的利用、避免出现处置困难和相关环境问题、充分发掘花生副产物尤其是花生粕的潜在功能和经济价值意义重大。
1.2 花生粕营养价值
花生粕富含多种营养成分和活性物质,花生粕粗蛋白含量为40%~50%[17],氨基酸含量高达39.65%,其中精氨酸含量5.2%[18],可溶性总糖含量在30%左右[19]。研究表明,花生多肽和多糖具有抗氧化、护肝、调节免疫、降血脂、血糖和抗菌抗病毒等生物活性[20]。并且花生粕中含有丰富的黄酮、多酚、三萜、鞣质或甾体类化合物等成分[21],但对其活性物质利用的研究还处在起步阶段。花生粕主要营养组成及含量见表1。
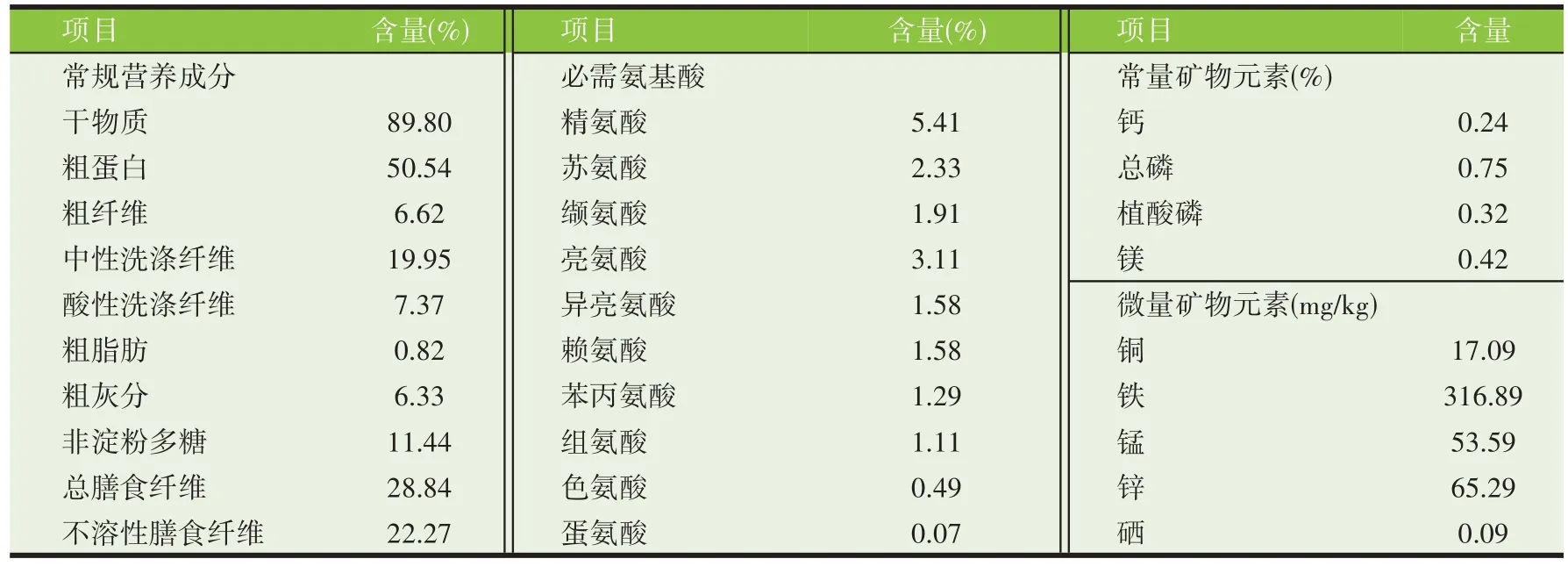
表1 花生粕主要营养成分
2 生物发酵花生粕研究现状
花生粕生物发酵是利用单一菌[23-28]、复合菌[29-31]或利用菌酶协同[4,32-33]发酵技术,实现花生粕的体外预消化,提高可溶性蛋白、多肽、游离氨基酸[4,22-24,30]和挥发性脂肪酸(VFA)含量[25],消除或降低花生粕中的抗营养因子[22],促进代谢产物的生物转化[26-27],降解霉菌毒素[28-29,32],从而获得新的高品质蛋白质饲料原料。同时发酵花生粕中的益生菌[34-35]、有机酸[23]、小肽[30]、黄酮类[22,27]和酶类[26,31]等生物活性产物,可抑制肠道有害病菌,提高微生物的多样性或有益微生物丰度,调节肠道微生态平衡[36],提高动物免疫力[37-38],预防肠道疾病和减少腹泻等[36],从而替代或减少抗生素在养殖过程中的使用。生物发酵技术的运用,可提高花生粕的资源化利用率,缓解养殖业蛋白质饲料原料紧缺,提高饲料消化率,降低动物粪便中氮、磷和气体的排放,减少粪便臭味和环境污染[39-40],进而实现节粮增效和环境生态效益。花生粕生物发酵对花生产业链延伸、畜牧业的安全高效持续发展具有重要意义。生物发酵花生粕在改善营养特性、提高抗氧化活性、酶活性、降解霉菌毒素等方面的效果明显,生物发酵花生粕相关的研究报道见表2。
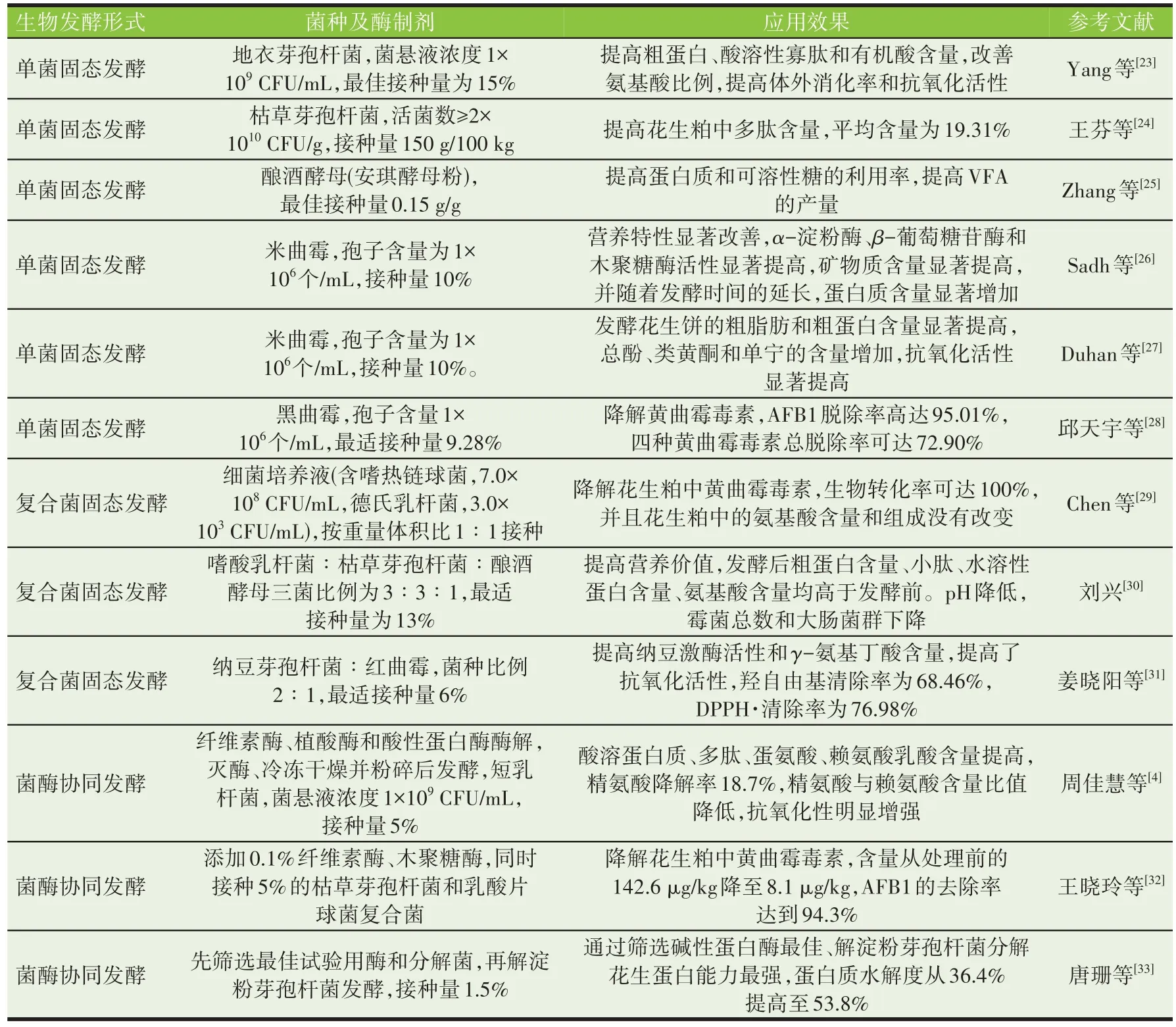
表2 生物发酵改善花生粕营养特性和生物功能的相关研究报道
3 花生粕在动物生产中的应用
3.1 花生粕在猪禽养殖中的应用
目前,对花生粕的研究更多的是聚焦在改善花生粕营养、脱毒、活性物质提取等方面,在猪饲粮中的应用研究鲜见报道,且在实际生猪养殖中使用甚少。2021年全国动物营养指导委员会在《猪饲料豆粕减量替代技术方案》中指出,花生饼粕的用量一般不超过10%。花生粕在家禽养殖,尤其是蛋鸡、鸭饲粮中有一定的使用。当前,迫切需要开发和评估的是在猪养殖中不可持续的传统蛋白原料替代品,缓解原料紧张和价格居高的状况,另一方面花生粕和菜籽粕等杂粕也迫切需要提高其资源化利用率和附加值。
解佑志等[39]报道,用8%的发酵花生粕替代生长猪基础日粮中的豆粕,结果蛋白质、钙、磷的消化率显著提高,并显著提高了日增重,降低了料重比,表明花生粕经生物发酵可改善蛋白质的消化率和促进猪的生长。张思轩等[40]报道,饲喂肉鸡花生粕杂粕型日粮,并且蛋白质水平降低2%,粪中吲哚、粪臭素含量可显著降低,粪中氮的含量及NH3浓度也显著降低,表明饲喂花生粕可以提高氮的利用率。梁海超[41]报道,育成鸡日粮中用花生粕替代30%的豆粕,与基础日粮组相比,替代组对育成鸡生长发育无不良影响,蛋鸡产蛋率也无显著差异,但蛋壳强度和蛋壳比重显著增加。Xia 等[42]的研究结果显示,随着花生粕替代豆粕水平的增加,蛋鸭产蛋量提高,蛋重和饲料消耗减少。蛋黄中ω-6 系列多不饱和脂肪酸的含量以及多不饱和脂酸的总含量增加;但在花生粕完全替代豆粕时,饲料转化率降低,产蛋量和抗氧化能力降低,表明花生粕取代产蛋鸭日粮中75%的豆粕,对产蛋量或蛋质量不会产生负面影响。
3.2 花生粕在反刍动物养殖中的应用
由于反刍动物自身存在庞大的瘤胃微生物发酵系统,使得花生粕在替代豆粕时其生产性能基本不受影响,在反刍动物养殖中的使用相对较多。
Oliveira 等[43]报道,用不同比例的花生饼粕替代浓缩料中的豆粕饲喂肉牛,结果显示肉牛的采食行为和生理反应无显著变化。Santos 等[44]研究发现,利用花生粕替代豆粕,泌乳奶牛的乳产量、总固形物含量、饲料转化效率、脂肪酸组成不受替代水平的影响,但牛奶蛋白质和酪蛋白含量随花生粕添加量的增加而降低,表明在不影响产奶量、总固形物含量和乳蛋白含量的前提下,可采用部分花生粕替代饲粮中的豆粕来降低饲料成本。Silva 等[45]报道,花生饼替代浓缩料中的豆粕,随替代比例的增加,波尔×本地山羊杂交后代的采食量、总增重和日均增重减少,但营养物质的消化率除粗脂肪增加外,其他指标没有受到影响。Valenca 等[46]报道,花生粕替代豆粕,羔羊肉中油酸、十五烷酸、十七烷酸和二十碳五烯酸的比例上升,棕榈酸的比例下降,高胆固醇脂肪酸的总和下降,表明花生粕改善了羔羊肉的脂质特征。靳亚星等[47]的研究发现,育肥绵羊饲喂花生粕替代50%、100%豆粕和棉粕的试验日粮,总能表观消化率显著提高,各处理间绵羊生长性能、屠宰性能、肉色、滴水损失、氨态氮均无显著差异,熟肉率提高,乙酸含量显著下降,丙酸、正丁酸、戊酸含量均无显著差异,且50%的替代水平获得了最佳的经济效益。
3.3 花生粕在水产动物养殖中的应用
随着水产养殖规模的不断扩大,水产饲料的需求大幅增加,2022年水产饲料产量达2 525.7万吨,比上年增长10.2%[1],致使鱼粉等优质蛋白质原料供应更加紧张。加速推进植物源低成本蛋白质原料的开发,可助力水产养殖的健康可持续发展。
李百安等[48]报道,发酵花生粕替代鱼粉,替代1/3和2/3 的用量时,对罗非鱼体增重率和饲料系数影响不显著。当完全替代鱼粉时,体增重率显著下降,饲料系数显著上升,但对消化道蛋白酶活性无显著影响。各组罗非鱼肌肉中常规营养指标含量及必需氨基酸总量等均无显著差异,表明在罗非鱼日粮中用发酵花生粕替代2/3 的鱼粉是可行的。Zhu 等[49]利用花生饼替代鱼粉,低比例的替代时可促进杂交石斑鱼的生长;当替代水平高于22%时,增重率下降;当高于44%时,会破坏肠道菌群的丰度,肠道有害细菌数量显著增加;当高于55%时,肝脏沉积脂肪的增加达到显著水平。因此,饲粮中适宜的花生饼替代水平为15%。Wang等[50]研究发现,饲料中添加混合植物蛋白(油菜籽粕、棉籽粕和花生粕)替代600 g/kg 豆粕对黄河鲤生长性能无不良影响。但当替代800 g/kg 豆粕时,抑制了黄河鲤的生长性能,血清超氧化物歧化酶(SOD)和溶菌酶活性降低,谷丙转氨酶(ALT)和谷草转氨酶(AST)活性升高,丙二醛(MDA)含量增加,胰岛素样生长因子-1(IGF-1)水平降低,肝脏生长激素(GH)和IGF-1基因的表达下调,表明用混合植物蛋白质替代一定量的豆粕不会影响黄河鲤的生长、免疫、肝脏功能等。刘毅等[51]利用花生粕替代豆粕,在替代用量低于50%时,对津新鲤2 号的体增重率、饲料系数和蛋白质效率无显著影响。各组鱼肌肉中必需和非必需氨基酸组成和含量均无显著变化,但随着替代比例的增加多不饱和脂肪酸呈下降趋势。
综合上述研究,花生粕替代部分豆粕、鱼粉对动物生产性能和产品品质无显著负面影响,表明在配合饲料中使用花生粕的替代方案是可行的,但适宜的替代比例不同报道之间存在明显的差异,需进一步的验证。花生粕发酵后营养特性和产品功能发生变化,但发酵后花生粕在不同动物配合饲料中替代豆粕或鱼粉的比例,在不同来源混合蛋白质饲粮中的添加量,替代或添加后对动物机体生产性能、营养物质消化酶和转运载体的分泌表达、肠道健康、免疫功能的影响,目前在动物试验中的研究报道较少。
4 小结
我国的花生粕资源十分丰富,并且价格低于豆粕和鱼粉,但由于花生粕蛋白品质欠佳,含有植酸等多种抗营养因子,易污染黄曲霉毒素,经高温压榨后花生蛋白的功能性质发生改变,导致蛋白质消化利用率不稳定,这些因素限制了花生粕的使用,因此迫切需要一种安全有效的方法来降解花生粕中的黄曲霉毒素,改善其营养特性和产品功能,提高其利用价值。生物发酵被认为是将农业和食品工业加工后产生的副产物转化为营养成分和生物活性物质的有效工具,因此,在迫切需要开发非常规蛋白质饲料资源时,生物发酵花生粕等杂粕越来越受到行业的重视。
目前,对花生粕的研究主要集中在以花生粕为发酵底物,优化发酵工艺参数,进行营养特性的改善、功能性食品或添加剂的开发,但对其发酵过程微生物群落的动态变化规律和改善花生粕品质的机理研究鲜见报道。在配合饲料中的应用研究,还主要是未发酵花生粕对动物生长性能、血清生化和肉蛋奶品质影响的报道,发酵花生粕在动物生产中的应用研究鲜见报道,其促进动物生长发育和调节免疫功能的作用机制尚不十分清楚。现有的研究初步表明,生物发酵花生粕,对提升原料品质、提高其养分消化率、节粮增效和减少环境污染等方面具有重要的作用,发酵花生粕饲用原料的开发将为缓解蛋白质饲料资源紧缺,实现安全、高效、健康养殖业的发展提供一条新选择。
