黄芪甲苷调控JAK2/STAT3/CXCL12信号通路抑制炎症细胞浸润改善小鼠胃炎的机制研究
2024-01-22江晓涛杨泽虹王姝烨安金琪黄远程邹雨杉文艺李培武
江晓涛, 杨泽虹, 王姝烨, 安金琪, 黄远程, 邹雨杉, 文艺, 李培武
(1.广州中医药大学第一临床医学院,广东广州 510006;2.广州中医药大学第一附属医院,广东广州 510405)
胃炎患者的胃黏膜中存在着多种炎症细胞的浸润[1]。过量浸润的炎症细胞通过分泌多种炎症介质诱发胃黏膜炎症反应[2-3],从而引起胃黏膜的损伤[4-5],导致症状反复。因此,抑制炎症细胞浸润是修复胃黏膜损伤、减少症状反复的关键。微环境中脂多糖(LPS)持续存在是导致炎症持续存在的主要原因,其可活化Janus 激酶2/信号传导和转录激活因子3(JAK2/STAT3)蛋白、转录因子κB(NF-κB)等经典炎症信号通路合成和释放多种细胞因子诱导免疫细胞浸润至免疫微环境[6]。JAK2/STAT3 通路作为重要的信号传导途径,广泛参与机体免疫调节、炎症反应、凋亡等过程[7-8]。STAT3 的磷酸化水平常被用于评估JAK2/STAT3信号通路的活化程度,磷酸化的STAT3 通常入核促进下游细胞因子的转录表达[9],进而诱导炎症细胞浸润。CXCL12作为关键的趋化因子,通常由促炎性刺激剂[LPS或肿瘤坏死因子(TNF)]激活STAT3、NF-κB、Sp1、AP1、PARP1等转录因子转录而来[10],其通过与免疫细胞膜上的受体趋化因子受体(CXCR)4/CXCR7相互作用诱导免疫细胞迁移至免疫微环境发挥功能[11]。因此,抑制CXCL12 上游转录因子激活是减少炎症细胞向微环境浸润的关键靶点。
多项临床研究[12-14]已经证实,黄芪及其相关中药方剂对于慢性胃炎有确切的疗效。黄芪甲苷(astragaloside Ⅳ)作为黄芪[为豆科植物蒙古黄芪Astragalus membranaceus(Fisch.)Bge.var.mongholicus(Bge.)Hsiao 或膜荚黄芪Astragalus membranaceus(Fisch.)Bge.的干燥根]的主要药理成分,已被报道具有一定的抑炎作用。本研究采用黄芪甲苷对LPS诱导的人胃黏膜上皮细胞GES-1 炎症模型和小鼠胃炎模型进行干预,通过检测JAK2/STAT3/CXCL12通路上相关分子的表达,探讨黄芪甲苷改善胃黏膜炎症的作用机理,以期为临床应用黄芪甲苷及黄芪治疗胃炎提供循证依据,现将研究结果报道如下。
1 材料与方法
1.1 细胞人胃黏膜上皮细胞株GES-1,购自中国科学院上海细胞库。
1. 2 动物SPF 级C57BL/6 小鼠24 只,5 周龄,体质量16~20 g,购自广东省医学实验动物中心,动物生产许可证号:SCXK(粤)2022-0002。动物饲养于广州中医药大学SPF级实验动物中心,动物使用许可证号:SYXK(粤)2018-0001。饲养环境温度(22±2)℃,湿度(50±5)%,5只/笼,自由饮水与摄食。动物实验经广州中医药大学实验动物伦理委员会批准,批准号:20230315013。
1. 3 药物、试剂与仪器黄芪甲苷(分子式C41H68O14,分子量784.97,纯度≥98%,生产厂家:上海源叶生物科技公司,批号:C14J9Q65866);p-STAT3抑制剂BP-1-102(MCE公司,批号:HY-100493)。幽门螺杆菌脂多糖(Hp-LPS)(卡迈舒公司,批号:22E031201);RPMI-1640 细胞培养液、胎牛血清(FBS)(美国Gibco 公司);细胞计数试剂盒8(CCK-8)(美国MCE 公司);RNA 提取试剂盒、RNA 反转录试剂盒(EZB 公司);兔源磷酸化STAT3(p-STAT3)抗体、兔源CXCL12 抗体、辣根过氧化物酶(HRP)标记的IgG 二抗(Abmart 公司);人源趋化因子CXCL12 酶联免疫吸附分析(ELISA)试剂盒、鼠源趋化因子CXCL12 ELISA 试剂盒(江苏酶免实业股份有限公司)。Nanodrop 核酸定量仪(美国Thermo 公司);荧光定量PCR 仪(瑞士罗氏公司);电泳仪(美国Bio-Rad 公司);多用途酶标仪(美国Biotek 公司);化学发光凝胶成像设备(美国ProteinSimple 公司);病理切片机(上海徕卡设备股份有限公司)。
1.4 体外实验
1.4.1 细胞培养 GES-1细胞用加入10%胎牛血清(FBS)、1% 青霉素的RPMI-1640 培养液在37 ℃和5%CO2条件下进行体外培养。定期更换培养液,直到超过80%进行传代培养。
1.4.2 LPS诱导GES-1炎症模型最佳浓度筛选将GES-1 细胞分成不同浓度(1、2.5、5、10、20 μg·mL-1)LPS 模型组及空白对照组(等体积PBS)。干预24 h 后,采用定量聚合酶链反应(qPCR)法检测不同浓度LPS 诱导的GSE-1 细胞CXCL12 mRNA 表达水平,ELISA 法检测上清液CXCL12因子含量,以此确定LPS的最佳浓度。
1.4.3 CCK-8法测定黄芪甲苷干预后GES-1细胞存活率 GES-1 细胞生长至对数生长期时,接种于96 孔板中,细胞密度为4 ×103个/孔,每孔100 μL培养液。细胞贴壁附着后,更换新的培养液,对照组加入100 μL完全培养液,黄芪甲苷3个浓度组分别加入黄芪甲苷终浓度为0.1、1、10 μg·mL-1的培养液100 μL(设置3 个复孔)。同时设置空白孔(加入不含细胞和黄芪甲苷的培养液)。24 h后,吸出培养液。每孔用100 μL 检测试剂(10 μL CCK-8+90 μL 无血清培养液)在37 ℃避光孵育1 h。应用酶标仪测定光密度(OD)值(波长设置为490 nm)。公式:细胞存活率=(OD给药-OD空白)/(OD对照-OD空白)。
1.4.4 黄芪甲苷抑制CXCL12 表达的最佳浓度筛选 GES-1 在6 孔板中按1 × 105个/孔的数量进行培养,并分成5 组,即空白对照组、模型组和黄芪甲苷0.1、1、10 μg·mL-1组。空白对照组不做干预,其余各组均加入最佳浓度的LPS,孵育24 h。黄芪甲苷3 个浓度组分别加入终浓度为0.1、1、10 μg·mL-1的黄芪甲苷培养液,黄芪甲苷使用0.05% DMSO 作为溶剂,处理24 h。采用qPCR法、ELISA法分别对应检测不同浓度黄芪甲苷干预下GSE-1 细胞CXCL12 mRNA 表达水平、上清液中CXCL12 因子含量,以此确定最佳的黄芪甲苷浓度。
1.4.5 黄芪甲苷与STAT3分子对接 在PDB数据库(https://www.rcsb.org/)获取STAT3 的PDB 格式,选择原则为:人源蛋白、分辨率高(小于等于2.5Å)、具有原始配体以及结晶pH值应尽量接近人体正常生理范围。从PubChem 数据库(https://pubchem.ncbi.nlm.nih.gov)将STAT3 分子结构下载至本地计算机,使用chendraw 绘制小分子的3D立体结构,使用AutoDock vina 对接蛋白和小分子,在PyMOL中实现可视化。
1.4.6 细胞分组 GES-1 细胞在6 孔板中,按照每孔1 × 105个的数量进行培养,培养液为2 mL。培养24 h,细胞贴壁后弃去培养液。将GES-1 细胞分成空白对照组、模型组、黄芪甲苷组和p-STAT3 抑制剂组。空白对照组不做干预,其余3 组给予最佳浓度(10 μg·mL-1)的LPS 处理24 h。黄芪甲苷组给予最佳浓度(10 μg·mL-1)的黄芪甲苷(用0.05%DMSO作为溶剂)最佳浓度处理24 h,阳性药物组即p-STAT3 抑制剂组,给予5.0 μmol·L-1p-STAT3 抑制剂BP-1-102(使用0.05%DMSO 作为溶剂)处理24 h。采用Western Blot法检测p-STAT3和CXCL12蛋白表达水平。
1.4.7 qPCR方法测定CXCL12 mRNA 表达量 从细胞中提取RNA 后,测定其浓度和纯度并转录成cDNA。反应体系为20 μL(包括Premix、上游引物、下游引物、模板cDNA 和MilliQ 水)。单个循环设置为95 ℃热启动10 min、95 ℃变性15 s、60 ℃退火延伸1 min,重复40 个循环。设定GAPDH 为内参基因,采用2-△△CT法测定mRNA 的相对表达量。人源CXCL12 基因的引物序列:正向,5’-ATTCTCAACACTCCAAACTGTGC-3’;反向,5’-ACTTTAGCTTCGGGTCAATGC-3’。 鼠源CXCL12 基因引物序列:正向,5’-GGAGGATAG ATGTGCTCTGGAAC-3’; 反向,5’-AGTGAGGA TGGAGACCGTGGTG-3’。引物由上海捷瑞生物工程有限公司设计与合成。
1. 4. 8 ELISA 法检测细胞上清液中CXCL12含量 对于细胞样本,以300×g离心10 min,去除沉渣,然后将上部的澄清液收集。用ELISA 法测定CXCL12在细胞上清液中的浓度。
1. 4. 9 Western Blot 法检测p-STAT3 和CXCL12蛋白表达水平 提取细胞总蛋白、胞浆和胞核蛋白,二喹啉甲酸(BCA)法测定细胞总蛋白、胞浆和胞核蛋白浓度,进行十二烷基硫酸钠(SDS)-聚丙烯酰胺凝胶电泳(PAGE)。每孔上样量为50 μg,电泳、转膜、封闭。后根据检测需要,添加针对目标蛋白的p-STAT3、CXCL12 一抗(稀释比均为1∶1 000),4 ℃孵育过夜。洗膜后,加入二抗室温孵育2 h。洗膜后,再添加曝光溶液进行曝光,利用凝胶成像系统进行图像处理。使用ImageJ 软件对得到的条带进行灰度处理,然后通过计算目的蛋白/内参蛋白(GAPDH)比值,表示目的蛋白的表达水平。
1.5 体内实验
1.5.1 小鼠分组、模型复制与给药 小鼠适应性喂养1周后,随机分成正常组、模型组、黄芪甲苷组和抑制剂组,每组6只。除正常组外,其他3组每日给予LPS 160 μg 灌胃和100 μg·mL-1LPS 自由饮用。持续1周后停止LPS灌胃,继续保持LPS自由饮用,同时,黄芪甲苷组给予黄芪甲苷(用1%的羧甲基纤维素钠溶液溶解)20 mg·kg-1·d-1[15-16]灌胃,p-STAT3 抑制剂组给予BP-1-102[用0.05%DMSO溶解]10 mg·kg-1·d-1[17]灌胃,干预1周后取材。
1.5.2 小鼠体质量测量及胃黏膜组织病理检测记录各组小鼠干预前后体质量。取材后,使用4%多聚甲醛溶液固定胃黏膜标本,再用乙醇梯度脱水、石蜡包埋。切片后,采用苏木素-伊红(HE)染色法观察组织学变化。
1.5.3 Western Blot法检测胃黏膜p-STAT3、CXCL12蛋白表达水平 取胃黏膜组织提取总蛋白、胞浆和胞核蛋白后,余操作同“1.4.9”项内容。
1. 5. 4 免疫组织化学法检测胃黏膜p-STAT3、CXCL12蛋白阳性表达水平 取小鼠胃黏膜组织经4%多聚甲醛溶液中固定、乙醇梯度脱水、包埋、切片后,脱蜡和水化,柠檬酸抗原修复缓冲液(pH6.0)抗原修复。3%过氧化氢溶液室温避光孵育25 min,3%牛血清白蛋白室温封闭30 min。加入一抗p-STAT3、CXCL12(稀释比例分别为1∶100、1∶300)4 ℃孵育过夜。加入HRP 标记的IgG二抗(稀释比例为1∶800)室温孵育50 min,3,3’-二氨基联苯胺(DAB)显色。苏木素复染、脱水、透明和封片,于显微镜下观察并拍照。使用Quant Center 2.3分析软件中PatternQuant模块将棕色区域(呈阳性)和蓝色部分区分,两部分Mask Area 作为组织面积,在棕色区域下一层级添加HistoQunant以统计阳性部分的面积,将阳性面积与组织面积的比值作为阳性率。
1.5.5 ELISA法用于检测血浆中CXCL12含量 采用EDTA 抗凝法采集血标本后,将其离心,收集上部的血浆。用ELISA法测定CXCL12在血浆中的浓度。
2 结果
2. 1 LPS 诱导GES-1 细胞炎症模型的浓度图1结果显示:10 μg·mL-1LPS 诱导GES-1 细胞24 h后,CXCL12 mRNA 表达水平和细胞上清液中CXCL12因子含量相比空白对照组上调最显著(P<
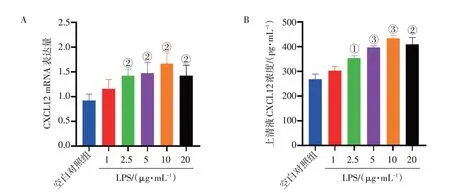
图1 不同浓度的脂多糖(LPS)对GES-1细胞CXCL12 mRNA和蛋白表达的影响(±s,n=3)Figure 1 Effects of different concentrations of lipopolysaccharide(LPS)on mRNA and protein expression of CXCL12 in GES-1 cells(±s,n=3)
0.01 或P<0.001),当LPS 浓度在10 μg·mL-1以上时,CXCL12 mRNA 表达水平和细胞上清液中CXCL12因子含量反而下降。因此,选择10 μg·mL-1作为诱导GES-1 细胞发生炎症模型的LPS 的最合适浓度,并应用于后续实验。
2.2 黄芪甲苷对GES-1 细胞存活率的影响图2结果显示,与空白对照组比较,黄芪甲苷0.1、1、10 μg·mL-1组GES-1 细胞存活率无显著性差异(P>0.05)。表明0.1、1、10 μg·mL-1黄芪甲苷对GES-1 细胞活力没有明显影响,故选择对0.1、1、10 μg·mL-1黄芪甲苷浓度开展后续的干预实验。
2.3 黄芪甲苷干预LPS 诱导的GES-1 细胞的最佳浓度图3结果显示:与空白对照组比较,10 μg·mL-1LPS组GES-1 细胞诱导24 h 后CXCL12 mRNA表达水平和上清液中CXCL12因子含量显著升高,差异有统计学意义(P<0.001);与模型组比较,黄芪甲苷0.1、1、10 μg·mL-1组CXCL12 mRNA 表达水平和细胞上清液中CXCL12因子含量均降低,且随黄芪甲苷浓度逐渐升高而逐渐下调,差异有统计学意义(P<0.05 或P<0.01)。当黄芪甲苷浓度为10 μg·mL-1时,CXCL12 mRNA 水平(P<0.01)和CXCL12 因子水平(P<0.05)下调最为显著,故选用本浓度开展后续实验。
2.4 黄芪甲苷与STAT3 分子对接图4 分子对接计算结果表明,黄芪甲苷能与STAT3 靶点发生结合,其结合能皆<5 kcal/mol,提示黄芪甲苷与STAT3 具有较强的结合力。STAT3 的氨基酸ARG-1127、ASN-986、GLN-955 与黄芪甲苷之间的氢键结合能较小,形成的活性口袋稳定,此为黄芪甲苷与STAT3结合的潜在位点。

图4 黄芪甲苷与STAT3对接结果Figure 4 Docking results of astragaloside Ⅳwith STAT3
2.5 黄芪甲苷对LPS 诱导的GES-1 细胞炎症模型p-STAT3、CXCL12 蛋白表达水平的影响图5 结果显示:模型组细胞中p-STAT3、CXCL12 蛋白表达水平均高于正常组(P<0.001);与模型组比较,黄芪甲苷组和p-STAT3 抑制剂组GES-1 细胞p-STAT3、CXCL12 蛋白表达水平显著降低(P<0.01 或P<0.001)。表明黄芪甲苷能抑制LPS 诱导的GES-1 细胞p-STAT3、CXCL12蛋白的表达。
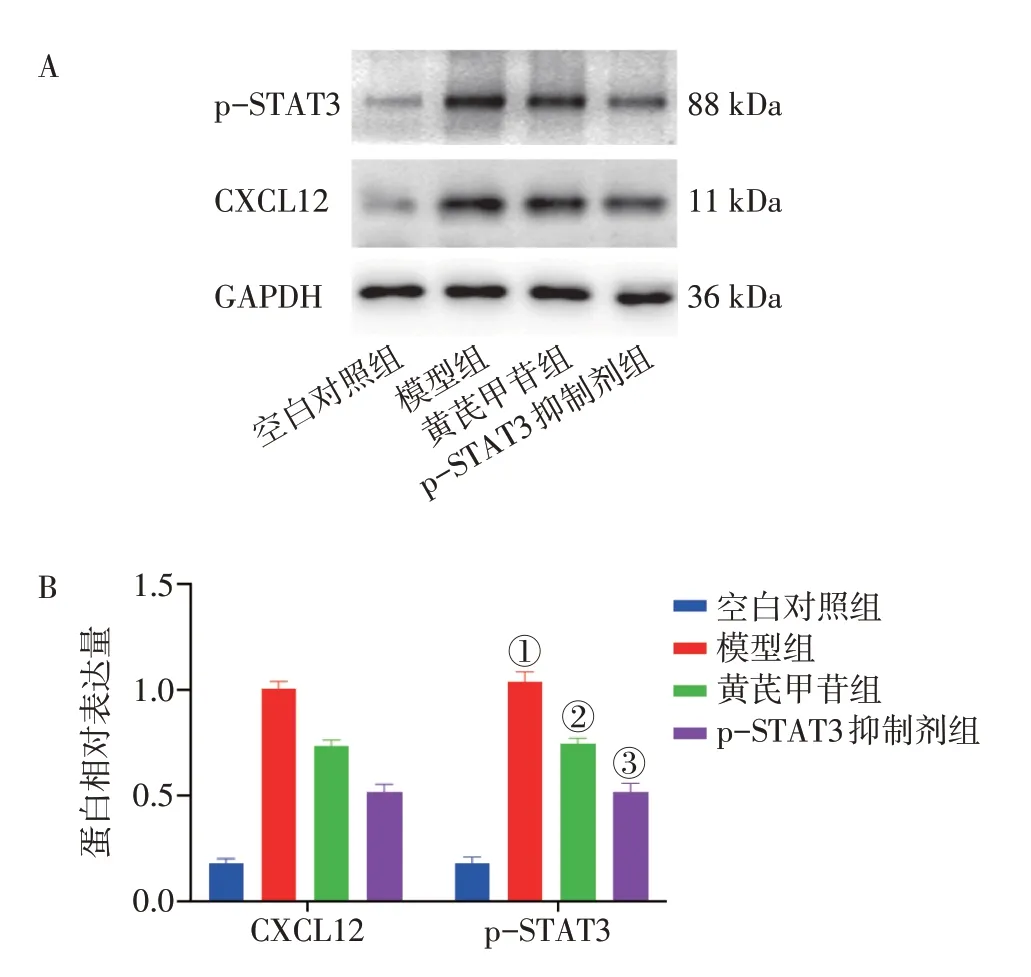
图5 黄芪甲苷对脂多糖(LPS)诱导的GES-1 细胞炎症模型p-STAT3、CXCL12蛋白表达水平的影响(±s,n=3)Figure 5 Effect of astragaloside Ⅳon protein expressions of p-STAT3 and CXCL12 in GES-1 inflammation model stimulated by LPS(±s,n=3)
2.6 黄芪甲苷对胃炎小鼠体质量和胃黏膜病理的影响图6-A 结果显示:与空白对照组比较,模型组小鼠的体质量显著降低(P<0.001);黄芪甲苷组和p-STAT3 抑制剂组小鼠体质量较模型组显著升高(P<0.01)。图6-B 组织病理学结果显示:空白对照组胃黏膜腺管排列整齐,壁细胞和主细胞之间界限清晰,固有层内未见炎症细胞浸润;模型组胃腺管紊乱,黏膜固有层变薄、紊乱,可见炎症细胞渗入;黄芪甲苷组和p-STAT3抑制剂组小鼠胃黏膜固有层炎症细胞数目减少,胃腺排列略显规整。提示黄芪甲苷可以改善胃炎小鼠的一般情况和病理损伤。
进入十二五以来,胜利油田低渗透区块的开发大幅提速,暴露出常规压裂增产效果差、稳产时间短、钻井数量多等问题,导致经济效益不高。针对这些问题,提出直井长缝大规模压裂开发整体区块的技术思路,采用大井距、小排距的井网部署,减少钻井数量、提高单井压裂增产效果和控制储量[1-2],达到增产和改善经济效益的目的。
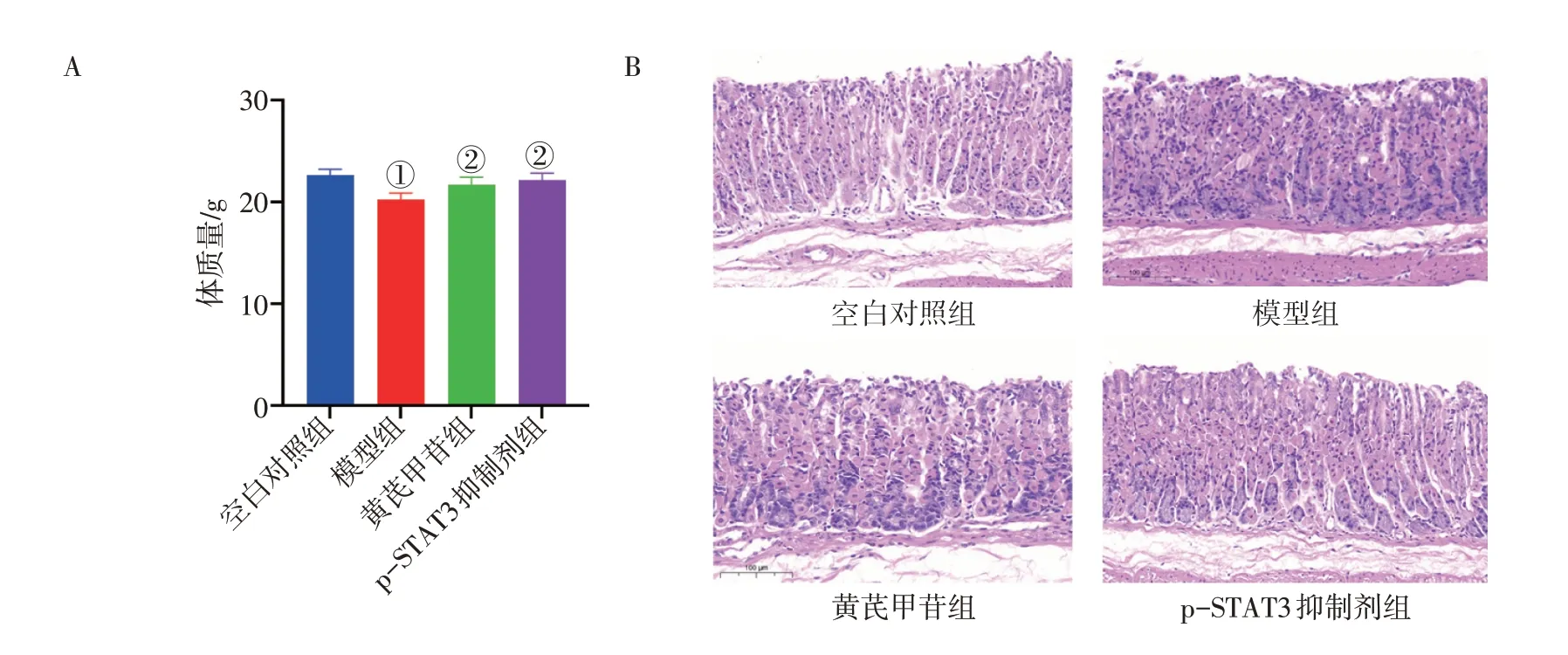
图6 黄芪甲苷对胃炎小鼠体质量和胃黏膜病理变化的影响(±s,n=6)Figure 6 Effect of astragaloside Ⅳon body mass and stomach mucosa histopathology of mice with gastritis(±s,n=6)
2. 7 黄芪甲苷对胃炎小鼠胃黏膜p-STAT3、CXCL12 蛋白表达水平的影响图7 结果显示:模型组胃黏膜p-STAT3、CXCL12 蛋白表达水平较空白对照组显著升高(P<0.01 或P<0.001);与模型组比较,黄芪甲苷组和p-STAT3 抑制剂组胃黏膜中p-STAT3、CXCL12 蛋白表达水平显著降低(P<0.05 或P<0.01 或P<0.001)。提示黄芪甲苷可以抑制LPS 诱导的胃炎小鼠胃黏膜中p-STAT3、CXCL12蛋白表达水平。
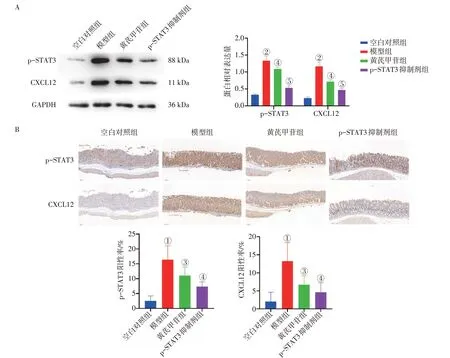
图7 黄芪甲苷对胃炎小鼠胃黏膜p-STAT3、CXCL12蛋白表达水平的影响(±s,n=6)Figure 7 Effect of astragaloside Ⅳon protein expressions of p-STAT3 and CXCL12 in gastric mucosa of mice with gastritis(±s,n=6)
2.8 黄芪甲苷对胃炎小鼠血浆中CXCL12 含量的影响图8结果显示:模型组血浆中CXCL12含量较空白对照组显著升高(P<0.01);黄芪甲苷组和p-STAT3抑制剂组血浆中CXCL12含量较模型组显著降低(P<0.05 或P<0.01)。提示黄芪甲苷可以下调LPS 诱导的胃炎小鼠血浆中CXCL12 因子水平。

图8 黄芪甲苷对胃炎小鼠血浆中CXCL12含量的影响(±s,n=6)Figure 8 Effect of astragaloside Ⅳon the content of CXCL12 in plasma of mice with gastritis(±s,n=6)
3 讨论
胃炎是消化系统的常见病和难治病,目前,我国幽门螺杆菌感染率为40.6%~55.8%,感染者几乎均存在慢性活动性胃炎[18]。大多数患者有不同程度的上腹部疼痛、腹胀、早饱等轻微症状,严重情况下可能会出现贫血、腹泻、呕血、黑便等,这对患者的生活质量和生命健康产生较大影响[19]。胃炎若得不到积极有效的干预,病程迁延,会伴有一定的癌变风险[19]。胃炎通常与胃黏膜上的炎症细胞浸润有关[20],炎症细胞不断释放炎症介质,导致非可控性炎症发生。持续的炎症会损伤胃黏膜,进一步加剧炎症反应[21]。因此,抑制黏膜微环境中炎症细胞的浸润是改善胃黏膜损伤的关键。
胃炎归属于中医学“胃脘痛”“胃痞”范畴,本病的基本病机为脾胃虚弱,补益脾胃是基本治法。黄芪是治疗脾胃消化系统疾病的常用药物,具有补中益气、利水消肿、敛疮生肌的功效[22]。有研究[23]报道,穴位注射黄芪注射液能改善慢性胃炎患者黏膜病理状态。黄芪建中汤、补中益气汤等以黄芪为君药的经典名方也被证实有助于纠正慢性胃炎患者的胃黏膜损伤[24-26]。黄芪甲苷是黄芪的主要活性物质,发挥调节免疫、抗炎的作用[27]。研究表明,黄芪甲苷可以通过升高大鼠血清中一氧化氮(NO)水平、提高超氧化物歧化酶(SOD)活性以及降低丙二醛(MAD)含量,改善酒精导致的急性胃黏膜损害[28],另外,还能调节Hedgehog 信号途径,进而对慢性萎缩性胃炎大鼠胃黏膜损伤具有一定的改善作用[29]。以上结果表明,黄芪甲苷在胃炎防治上有潜在的临床意义。
本研究使用LPS 诱导构建体外胃上皮细胞炎症模型和体内胃炎小鼠模型,发现p-STAT3、CXCL12表达水平上调,小鼠胃黏膜炎症细胞浸润增加。STAT3 磷酸化是JAK2/STAT3 信号通路活化的关键标志之一,磷酸化的STAT3 通常入核促进下游一系列细胞因子的转录表达[9-10],进而趋化炎症细胞浸润。CXCL12 又称基质细胞衍生因子1(SDF-1),编码序列位于10q11.1,其与CXCR4/CXCR7 受体结合对淋巴细胞有强烈的趋化作用[11]。故本研究结果提示,LPS 可激活JAK2/STAT3信号通路,促进CXCL12的表达并介导炎症细胞浸润及炎症发生。
分子对接是一种计算生物学和药物设计中的重要技术,常用于研究确定潜在药物与生物分子(通常是蛋白质)的结合方式,有助于理解药物与生物分子的相互作用[30]。分子对接结果显示,黄芪甲苷与STAT3 具有较高的亲和力,提示黄芪甲苷具有潜在抑制STAT3 的活性。进一步使用黄芪甲苷干预LPS 诱导的GES-1 炎症模型后,细胞中CXCL12 mRNA 表达水平和上清液中CXCL12 含量显著下调,STAT3磷酸化水平明显降低。体内研究结果显示,黄芪甲苷能改善LPS诱导的胃炎小鼠胃黏膜病理情况,抑制胃炎小鼠胃黏膜中p-STAT3蛋白磷酸化水平,下调胃黏膜和血浆中CXCL12因子的含量。上述研究结果提示,黄芪甲苷可能是通过抑制STAT3 的磷酸化,进而抑制JAK2/STAT3通路的活化及下游CXCL12 的转录,CXCL12-CXCR4/CXCR7 趋化信号进一步受到抑制,使炎症细胞浸润减少,黏膜炎症改善。
综上所述,黄芪甲苷可能是通过抑制JAK2/STAT3 信号通路活化,抑制CXCL12 表达,进而减少炎症细胞浸润,达到改善胃黏膜炎症水平的作用。
