大渡河理县高原鳅体长体重关系、遗传多样性和分化水平
2024-01-22赖见生陈叶雨黄颖颖赵凤麒陈永胜周启红刘晓欢
赖见生,陈叶雨,颜 涛,林 珏,黄颖颖,赵凤麒,陈永胜,周启红,刘晓欢,何 斌
(1.四川省农业科学院水产研究所,成都 611731;2.宜宾市农业农村局,四川 宜宾 644000;3.宜宾市翠屏区农业农村局,四川 宜宾 644000)
【研究意义】理县高原鳅(Triplophysalixianensis)属鲤形目(Cypriniformes)、鳅科(Cobitidae)、条鳅亚科(Nemachilinae)、高原鳅属。高原鳅是一种重要的淡水鱼类,大多栖息于青藏高原及附近的湖泊、河流和溪流中。高源鳅属具有高度多样性,由大约140种组成[1],该类群的起源和演化与青藏高原的隆升有关[2]。作为青藏高原鱼类区系的重要组成部分,高原鳅对高原低氧、低压和寒冷的环境有独特的形态和生理适应。高原鳅对当地环境具有强烈适应性,表明它们的长期进化与高原生境有关。因此,高原鳅是研究历史气候和地质事件对该属生物多样性模式影响的理想物种。在中国的高原鳅种类中,目前在四川鉴定的有23种[3]。2008年何春林等[3]在四川省阿坝州杂谷瑙河流域采集到高原鳅标本,经形态学鉴定为高原鳅新种,并命名为理县高原鳅。目前,关于理县高原鳅的报道较少,仅见于其作为新种的描述及线粒体DNA标记的开发[3-4]。有关长江流域理县高原鳅种群结构和种群历史动态的信息仍然处于空白。因此,对理县高原鳅不同地理种群的遗传多样性和群体结构进行评价,对进一步利用理县高原鳅的遗传种质资源和野生群体保护具有重要意义。【前人研究进展】线粒体DNA(Mitochondrial DNA,mtDNA)具有进化速度快、缺少重组、分子小、易于测序及分析等特点,目前已成为鱼类物种鉴定、进化遗传学、分子生态学、群体遗传结构与系统地理学等多种研究领域的重要标记[5-9]。控制区(Control region,D-loop)作为线粒体DNA上最重要的一段非编码区,遗传变异程度高,碱基替换率比线粒体DNA上的其他部分高5~10倍,是适合种群水平遗传多样性研究特别是遗传多样性较低的群体遗传学研究的最优标记之一[10]。研究表明,理县高原鳅的分布非常局限,仅在长江上游岷江流域杂谷脑河发现[3-4]。【本研究切入点】通过对大渡河流域鱼类资源调查发现,在大渡河4个不同地段,包括金川县沙耳乡、石棉县安顺彝族乡、泸定县冷碛镇和丹巴县格宗镇均发现有理县高原鳅分布,这也是首次发现其出现于杂谷脑河以外的流域。【拟解决的关键问题】选取该4个不同地理种群的理县高原鳅群体为研究对象,分析其体长与体重分布特征及其关系,并通过线粒体标记对其进行遗传变异检测,为揭示大渡河理县高原鳅的遗传背景及今后的种质资源保护提供理论基础。
1 材料与方法
1.1 材料
于2021年在大渡河采集理县高原鳅样本,金川县沙耳乡采集30尾(31°30′50.3″ E, 102°38′5.4″ N,编号g1:1~30),大渡河石棉县安顺彝族乡采集34尾(29°15′8.3″ E, 102°19′6.7″ N,编号g 2:31~64),大渡河泸定县冷碛镇采集30尾(102°13′20.9″ E,29°46′57.3″ N,编号g 3:65~94),丹巴县格宗镇采集23尾(30°48′26.6″ E, 101°56′25.2″ N,编号g 4:95~117),共117尾,剪取鳍条保存在100%酒精管,并存于4 ℃冰箱直至提取组织基因组。采样点分布如图1所示,水系图来源于四川测绘地理信息局(http://scsm.mnr.gov.cn/index.htm)。丹巴县格宗镇23尾理县高原鳅年龄段为3~9龄,大渡河石棉县34尾年龄段为6~11龄,泸定县冷碛镇30尾年龄段为6~11龄,金川县沙耳乡30尾年龄段为6~11龄,4个种群理县高原鳅年龄段均集中在6~7龄。大渡河石棉县34尾,包括14尾雌鱼和20尾雄鱼,泸定县冷碛镇30尾,包括13尾雌鱼和17尾雄鱼,金川县沙耳乡30尾,包括13尾雌鱼和17尾雄鱼,丹巴县格宗镇23尾鱼,包含7尾雌鱼和10尾雄鱼(其中6尾幼龄鱼无法鉴定性别)。雌雄鱼性腺发育期集中在III~IV期。
1.2 理县高原鳅体长-体重的相关性及肥满度分析
理县高原鳅的体长-体重关系用幂函数公式拟合[11]:
W=aLb

图1 理县高原鳅调查采样站位Fig.1 Sampling stations of T. lixianensis
式中,W为体重(g);L为体长(cm);a为条件因子,b为异速生长因子[12]。测量理县高原鳅全长时,取吻端到尾鳍末端的最大长度;体长即标准长,为吻端到尾鳍基的直线长度,即全长减去尾鳍的长度。将体重和体长数据进行对数转化,利用SPSS 19.0软件,使用线性回归法进行拟合,估计参数a和b的95%置信区间(CI)及R2的显著性水平。
肥满度采用Fulton状态指数K计算:
K=(W/L3)×100
式中,W为体重(g);L为体长(cm)。数据以“平均数±标准差”表示。
1.3 理县高原鳅基因组 DNA制备
理县高原鳅基因组DNA提取采用CTAB高盐法,取1 μL样品通过电泳检测DNA质量,并用超微量分光光度计(Implen N50 touch)检测DNA的纯度和含量,保存于-20 ℃冰箱备用。
1.4 理县高原鳅线粒体D-loop序列引物设计、合成
根据理县高原鳅线粒体序列信息(GenBank accession number: KT966735.1),在其D-loop区上下设计特异性引物,基因引物序列F:5’-CTCACCCCTGGCTCCCAAAGC-3’,R:5’-AGGGCCCGTCTTAGCATCTTC-3’。
1.5 理县高原鳅线粒体DNA PCR扩增体系及程序
理县高原鳅线粒体DNA扩增体系为1 μL基因组DNA(50 ng),2×Premix LATaq聚合酶25 μL,正反向引物各1.5 μL(10 μmol/L),PCR级水21 μL,总扩增体系50 μL。PCR反应程序包括:94 ℃预变性5 min;94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,32个循环;72 ℃延伸8 min。PCR产物用1.2%的琼脂糖凝胶电泳检测,将片段大小一致且条带单一的扩增产物进行测序。
1.6 数据分析
测序结果通过BLAST程序与GenBank中理县高原鳅线粒体DNA序列进行比对分析,确认扩增产物为理县高原鳅线粒体DNA D-loop区序列。多样性相关参数,包括核苷酸多样度(Pi)、单倍型多样度(Hd)、多态位点数(S)、单倍型数(H)和平均核苷酸变异数(K)等,用DNAsp 5软件进行计算。群体遗传分化指数Fst值利用Arlequin 2.0软件统计。系统发育树利用MEGA 6软件中的邻接法(Neighbor-Joining)进行构建。使用Arlequin 2.0软件计算错配分布(Mismatch distribution)、Tajima’sD和Fu’sFs值来检验群体历史上是否经历扩张, 通过偏差平方和(Sum of squared deviation, SSD)检测核苷酸不配对分布与种群扩张模型下期望分布之间的拟合优度。
2 结果与分析
2.1 理县高原鳅的体重-体长关系
从图2可知,大渡河理县高原鳅全长7.5~18.5 cm,体长6.3~15.4 cm,平均体长11.5 cm,体重2.8~44.9 g,平均体重19.6 g。a值为0.0123,95%置信区间为0.0081~0.0187,b值为2.9734,95%置信区间为2.8030~3.1440,R2>0.9121(表1)。本研究采用Fulton状态指数K表示大渡河理县高原鳅的肥满度,理县高原鳅的肥满度为1.18±0.02(平均数±标准差)。
2.2 理县高原鳅线粒体DNA控制区碱基特征
117尾理县高原鳅线粒体DNA控制区序列PCR产物特异性较好。对117尾理县高原鳅个体的测序结果比对分析发现,去掉引物及部分端部序列,同源重排后的长度为878 bp。序列中T、C、A、G碱基所占比例分别为32.5%、21.4%、32.9%、13.2%,AT和CG所占比例为65.4%和34.6%,可见AT比例显著高于CG比例(表2),说明理县高原鳅线粒体DNA组成符合硬骨鱼线粒体DNA碱基组成的特征。
2.3 理县高原鳅不同地理种群的遗传结构
金川县、石棉县、泸定县和丹巴县4个理县高原鳅种群D-loop区共存在32个变异位点和28个单倍型(表3)。4个理县高原鳅群体的单倍型多样度(Hd)在0.637~0.921,丹巴县格宗镇样本单倍型多样度最高。理县高原鳅群体的核苷酸多样性在0.000 91~0.005 70,大渡河泸定县冷碛镇样本核苷酸多样性最低(表3)。金川县沙耳乡样本理县高原鳅有11个单倍型,大渡河石棉县安顺彝族乡样本有7个单倍型,大渡河泸定县冷碛镇样本有7个单倍型,丹巴县格宗镇理县高原鳅样本有11个单倍型。单倍型Hap1出现的频率最高,但4个理县高原鳅地理种群未有共享的单倍型。金川县和丹巴县2个理县高原鳅群体之间的个体存在共享的单倍型,石棉县和泸定县2个种群之间也存在共享单倍型,但金川县和石棉县或泸定县群体之间的个体未出现共享单倍型,丹巴县和石棉县或泸定县理县高原鳅群体之间的个体也未出现共享单倍型(表4)。
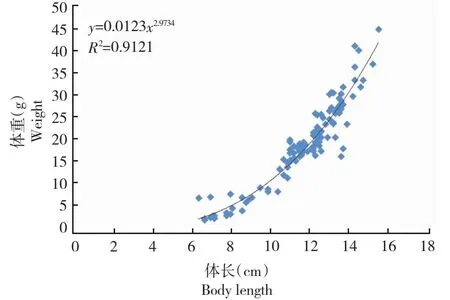
图2 理县高原鳅体长和体重的回归曲线Fig.2 Regression curves of relationship between body weight and body length of T. lixianensis
从表5可知,金川县沙耳乡和石棉及大渡河泸定县冷碛镇的理县高原鳅样本遗传距离分别为0.251 73和0.275 55,说明发生了中度遗传分化,与丹巴县格宗镇样本之间的遗传距离为-0.001 39,表明未出现遗传分化。石棉和大渡河泸定县及丹巴县格宗镇理县高原鳅样本之间的遗传距离为0.025 99和0.231 92,大渡河泸定县和丹巴县格宗镇样本之间的遗传距离为0.257 09,说明发生了遗传分化。

表1 理县高原鳅体重和体长关系函数的a和b参数

表2 不同地理种群理县高原鳅mtDNA控制区序列碱基组成
2.4 理县高原鳅不同地理种群的系统进化树
利用MEGA建立理县高原鳅4个地理种群个体的NJ分子系统树(图3),从系统树可以看出,117个理县高原鳅个体在进化树上形成2个独立的分支。其中,金川县沙耳乡和丹巴县格宗镇的个体形成一个独立分支,而大渡河石棉县安顺彝族乡和泸定县冷碛镇的个体形成另一个分支,与各地理种群的遗传距离结果一致。
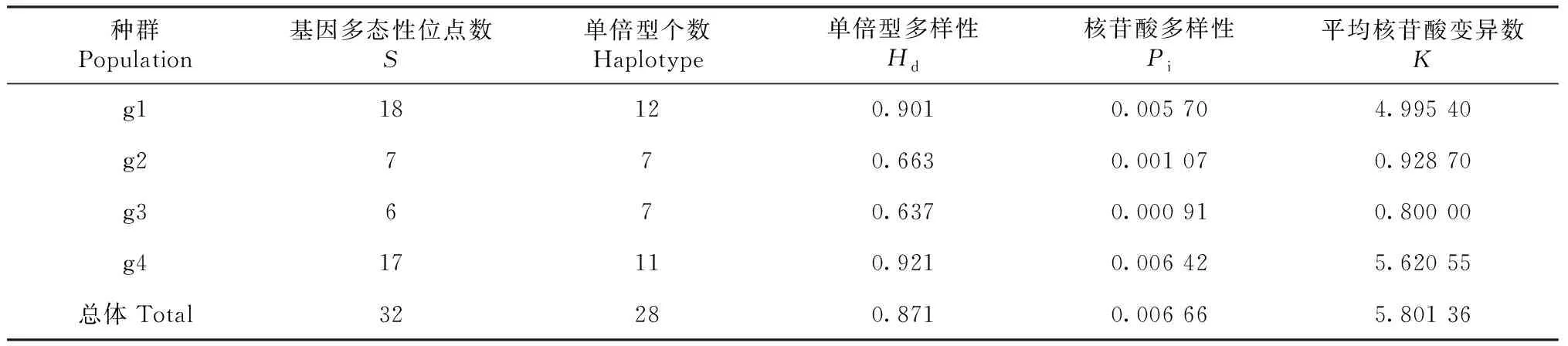
表3 理县高原鳅不同地理种群的遗传多样性参数
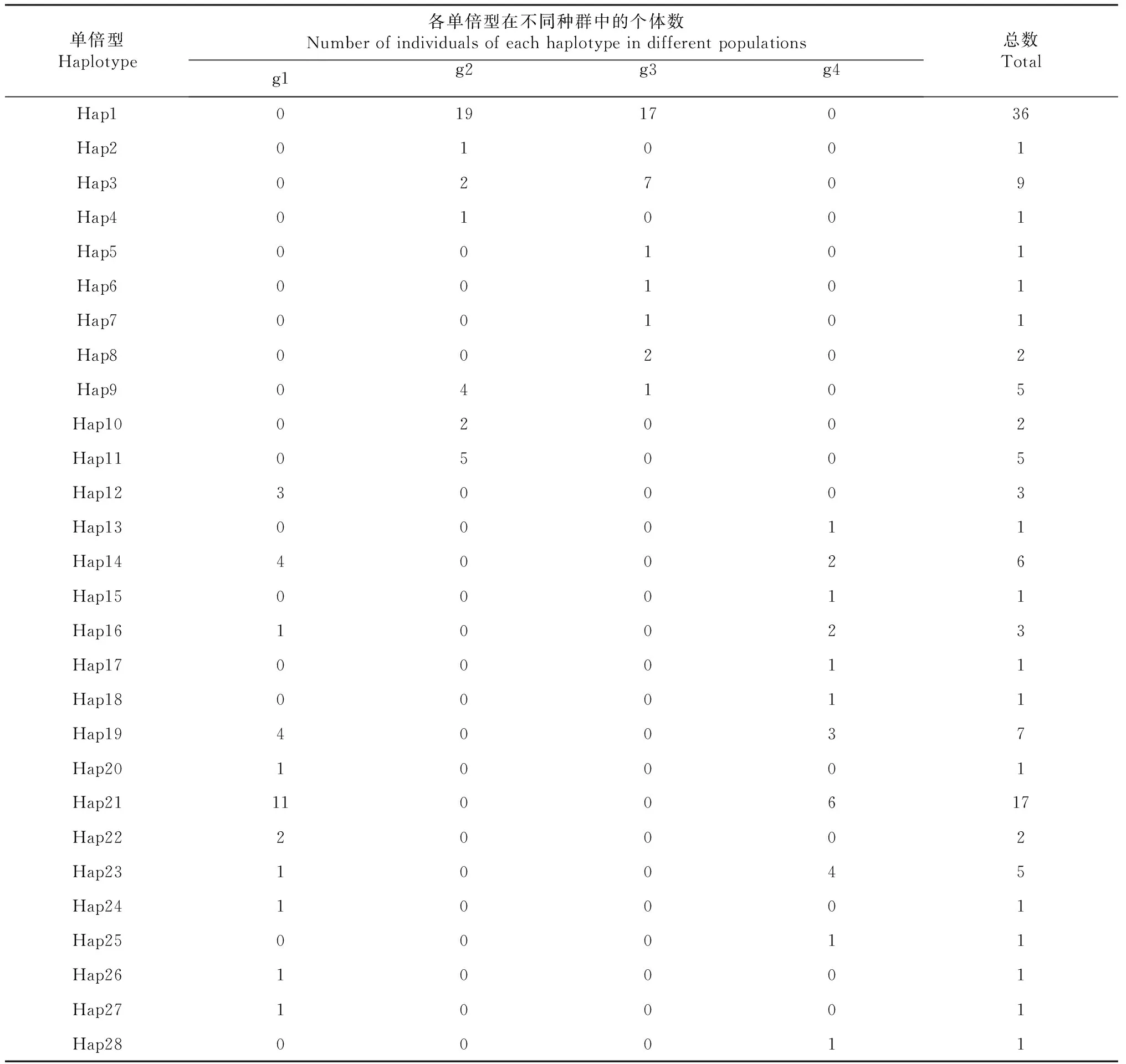
表4 理县高原鳅不同种群的28个单倍型分布
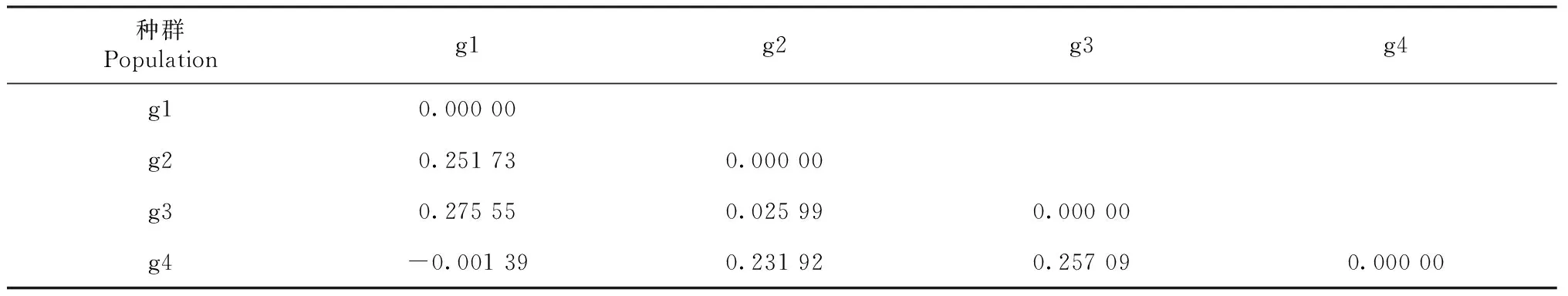
表5 理县高原鳅4个地理种群的遗传分化Fst值
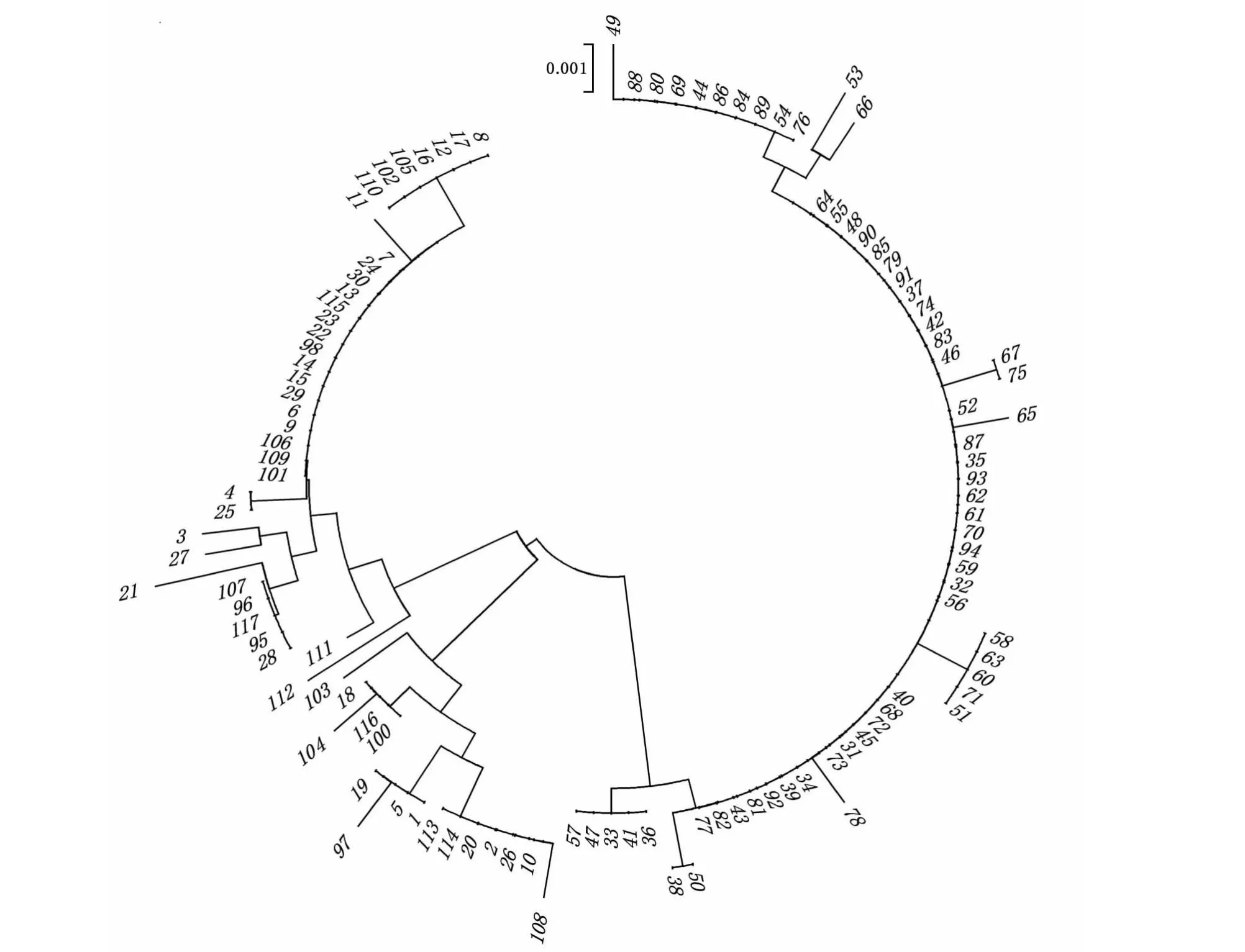
金川县:1~30;石棉县:31~64;泸定县:65~94;丹巴县:95~117。Jinchuan county: 1-30; Shimian county: 31-64; Luding county: 65-94; Danba county: 95-117.图3 基于线粒体D-loop区序列构建的NJ 系统树Fig.3 NJ tree based on D-loop region of mitochondrial DNA
2.5 理县高原鳅种群历史动态分析
中性检验结果显示, 金川县和丹巴县理县高原鳅地理群体的Tajima’sD值大于0,而石棉县和泸定县地理群体的Tajima’sD值小于0,且均未达到显著水平(P>0.05)。Fu’sFs检验结果显示, 4个理县高原鳅地理群体的Fu’sFs均小于0,其中,石棉县和泸定县群体均呈显著负值(P<0.05), 金川县和丹巴县群体均呈不显著负值(P>0.05,表6)。
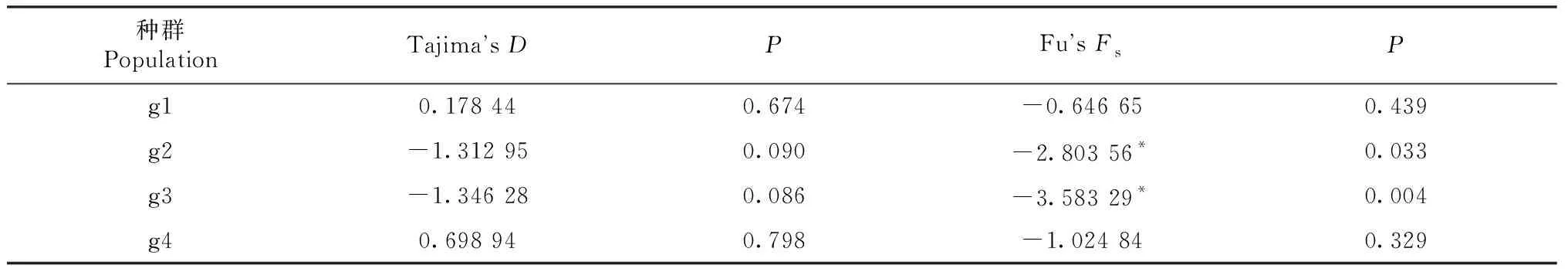
表6 理县高原鳅4个地理种群的中性检验

柱状图代表出现频率的观测值,曲线代表基于种群扩张事件模型的期望频率。A:金川县,B:石棉县,C:泸定县,D:丹巴县。The histograms indicate the observed frequencies, and the lines represent the expected frequencies based on the model of sudden population expansion. A: Jinchuan county, B: Shimian county, C: Luding county, D: Danba county.图4 理县高原鳅4个地理种群的单倍型错配分布 Fig.4 Mismatch distribution of haplotypes in four populations of T. lixianensis
对单倍型错配分布分析发现,金川县和丹巴县理县高原鳅地理群体的单倍型错配分布呈多峰分布(图4),同时,错配分析SSD不显著;而石棉县和泸定县理县高原鳅地理群体的单倍型错配分布呈单峰分布且SSD不显著,表明2个种群可能经历过群体扩张(图4-B、4-C)。
3 讨 论
本研究中,理县高原鳅的生长参数b值为2.9734,当b值小于3时,种群为负异速生长, 即体长的增速大于体重的增速,当b值等于3 时,种群为等速生长,当b值大于3 时,种群为正异速生长,体型表现为肥满而不是纤细[13]。本研究表明,大渡河理县高原鳅的b值保持在2.5~3.5的预期范围内,接近3,说明采样时间段(10—12月)理县高原鳅生长状况良好。由于缺乏理县高原鳅体重体长关系的文献报道,因此本研究结果无法进行比较。但研究显示,阿克苏河小鳔高原鳅b值为2.876[14],塔里木河巨头高原鳅b值为2.583[14],伊犁河斯氏高原鳅b值为1.22[15],伊犁河黑斑高原鳅b值为1.113[15],乌伦古河小体高原鳅b值为 2.52[16],湟水河似鲇高原鳅b值为2.67[17]。大多数高原鳅属鱼类b值小于3,处于负异速生长,而大渡河理县高原鳅b值高于其同属的其他高原鳅。b值的差异可以归因于多种因素,如检测的标本数量、捕获个体的长度范围、栖息地类型、季节性和地理区域[16,18-20]。生长环境是显著影响b值的因素之一[21]。Jisr等[22]发现,棕线石斑鱼等的生长参数b值在高温环境下明显高于低温环境。Sileesh等[23]对黑头鱼、深海鲱,蛇鲭等研究也表明b值水平显著受环境因素影响。
本研究采用线粒体基因组中进化速率最快的控制区序列分析方法,研究理县高原鳅在大渡河的4个不同地理种群共117个个体的碱基组成和变异情况,获得理县高原鳅野生群体的种质资源遗传变异基本数据。117个理县高原鳅个体的mt DNA 控制区序列中,4种碱基组成呈现出一定的偏向性,各群体均存在A+T所占比例高于C+G的现象,这与绝大多数鱼类的线粒体控制区组成特征相符[24]。
核苷酸多样性指数(Pi)和单倍型多样性指数(Hd)是线粒体DNA用来评估群体遗传多样性的2个重要参数。通过对理县高原鳅D-loop区序列分析发现,其总体的Hd在0.8以上,处于较高水平,而Pi偏低,仅为0.000 91~0.005 70。说明,目前理县高原鳅野生群体存在单倍型多样性较高而核苷酸多样性较低的情况,推测理县高原鳅种群可能正从一个更小的有效种群经历种群扩张,它们在较短的时间内,通过碱基突变积累其单倍型多态性,但不足以使核苷酸多态性也处于较高状态。由于核苷酸多样性指数考虑各单倍型在群体中所占的比例,因而较单倍型多样性指数更可靠[25]。类似结果在同属的其余高原鳅种群中也有发现。晁燕等[26]利用线粒体DNA cytb序列对大通河区域的似鲇高原鳅进行遗传多样性研究,结果显示似鲇高原鳅大通河种群的Hd为0.86,Pi为0.0036。颜渊等[27]利用D-loop区对西部黄河流域不同水域的黄河高原鳅种群研究发现,其Hd在0.9420~1.0000,Pi为0.0030~0.0043;杨成等[28]对拟硬刺高原鳅黄河种群和宝库河种群进行研究,发现其Hd为0.65~0.89,Pi为0.0010~0.0025;姚雁鸿等[29]利用线粒体D-loop序列对湖南乌龙山湘西盲高原鳅线粒进行分析,发现其Hd在0.083~0.282,其Pi范围为0.000 09~0.000 32。可见,以核苷酸多样性指数判断,目前理县高原鳅属的物种大部分都处于遗传多样性较低的状态,需要进行有效的遗传管理来增加其种群的遗传多样性。而从不同地理种群多样性差异分析,金川县和丹巴县理县高原鳅种群核苷酸多样度远远高于石棉县和泸定县种群。金川县和丹巴县属于藏区,人类活动较少,无人为干扰的生境保留了其遗传信息。前期实地调查也发现,石棉县和泸定县人为捕捞等活动更频繁,导致该地段鱼类种群的多样性偏低。
通过构建进化树可以发现,4个理县高原鳅地理种群的个体在NJ系统树上形成2大分支,推测这117个个体的单倍型可能来源于2个不同遗传背景的祖先。其中,金川县沙耳乡和丹巴县格宗镇的理县高原鳅个体形成一个独立分支,而大渡河石棉县安顺彝族乡和泸定县冷碛镇的个体形成另一个分支,说明金川县、丹巴县群体和石棉县、泸定县群体形成明显的分化。单倍型分布、遗传分化和聚类分析结果均显示大渡河金川县和丹巴县2个理县高原鳅种群之间存在广泛的基因交流,石棉县和泸定县之间的种群也存在基因交流。金川县和石棉县或泸定县之间,丹巴县和石棉县或泸定县之间的理县高原鳅种群均未发生基因交流,群体间存在显著的遗传分化,基于控制区序列的系统发育树也显示了两两种群之间形成独立分支,也支持上述结论。4个理县高原鳅群体的中性检验结果表明,金川县和丹巴县的Tajima’sD值大于0,Fu’sFs小于0,但均未达到显著水平,说明未检测到群体扩张迹象,可能未发生群体扩张事件。而石棉县和泸定县理县高原鳅地理群体的Tajima’sD和Fu’sFs均小于0,且后者呈现显著负值,表明这2个群体可能经历过群体快速扩张事件。石棉县和泸定县理县高原鳅地理群体的歧点分布呈单峰分布, 且SSD也不显著,进一步说明该2个群体动态未显著偏离原假设(种群扩张模型),近期都经历了快速的群体扩张。
目前,大渡河丹巴县和泸定县之间暂无已建电站,无人为因素阻碍该2个区域理县高原鳅的基因交流;而丹巴县和金川县及石棉县之间存在数个已建和在建电站,包括猴子岩水电站、长河坝水电站、黄金坪水电站、泸定水电站等,可能在一定程度上阻碍了坝上坝下鱼类的基因交流。对于物种遗传多样性和物种完整性的保护在于客观鉴别物种的关键遗传单元,然后采用合适的方式管理这些单元[30]。
4 结 论
目前大渡河流域石棉至金川段理县高原鳅生长状态良好。线粒体分子标记分析发现,大渡河理县高原鳅种群遗传多样性处于较低水平,并建议将大渡河金川县和丹巴县种群作为一个整体进行管理和保护,石棉县和泸定县也作为一个整体进行管理和保护,而金川县和丹巴县、石棉县和泸定县的种群分开管理和保护。
