针阔人工混交林及其纯林对土壤微生物碳循环功能基因丰度的影响
2024-01-22秦佳琪肖指柔明安刚朱豪滕金倩梁泽丽陶怡覃林
秦佳琪,肖指柔,明安刚,朱豪,滕金倩,梁泽丽,陶怡,覃林*
1. 广西大学林学院/广西森林生态与保育重点实验室,广西 南宁 530004;2. 中国林业科学研究院热带林业实验中心,广西 凭祥 532600;3. 广西友谊关森林生态系统定位观测研究站,广西 凭祥 532600;4. 崇左凭祥友谊关森林生态系统广西野外科学观测站,广西 凭祥 532600
土壤碳循环是生物地球化学循环中最重要和复杂的代谢过程之一(Lynn et al.,2017;Hu et al.,2019),调节着全球气候变化(Deng et al.,2017)。微生物是调控土壤碳循环的关键驱动因素,包括参与土壤有机碳固定、降解及甲烷代谢等一系列相互关联的碳迁移转化过程(Kuypers et al.,2018),推动着生态系统的能量流动和物质循环。土壤微生物具有编码碳循环相关酶系统的整套基因,其基因丰度可以反映相应酶的活性,基因丰度高则相应的微生物酶活性较强(Trivedi et al.,2016)。同时,土壤微生物碳循环功能基因丰度能够预测土壤微生物的物质循环潜力和代谢特征,可表征土壤有机碳的稳定性(Bani et al.,2018)。
森林土壤碳储量约占全球陆地土壤总碳库的73%(Pan et al.,2011)。近年来,随着宏基因组测序技术的发展,森林土壤微生物碳循环功能基因已逐渐成为生态学研究的热点。胡明慧等(2022)认为自然增温通过改变土壤微生物群落结构、碳代谢相关功能基因丰度来影响南亚热带常绿阔叶林土壤有机碳代谢过程;Wang et al.(2019)发现天然次生林土壤的乙二醛氧化酶、木质素过氧化物酶和锰过氧化物酶等木质素碳降解基因丰度均高于乡土和外来树种人工林,并且土壤养分、树种丰富度、凋落物储量、有效氮、粘土和粉砂含量等显著影响木质素降解基因的组成。还有研究表明,天然阔叶林转化为人工林和次生林后导致土壤碳固定、碳降解和甲烷代谢等相关基因丰度下降,其原因是森林转化通过降低土壤微生物生物量碳(MBC)和可溶性有机碳(DOC)含量及土壤pH、土壤湿度和土壤粘土含量间接降低土壤微生物碳循环基因丰度(Luo et al.,2022)。目前关于森林土壤碳循环功能基因的研究基本上限于天然林、次生林以及人工林土壤表层,而对于针阔人工混交林及其纯林在不同土壤深度的微生物碳循环功能基因丰度还知之甚少。
随着全球变暖加剧,碳中和已成为全球可持续发展的目标(Tan et al.,2021;Cui et al.,2022;陈科屹等,2022)。营造人工林是实现碳中和战略的重要举措。然而,长期的人工林经营发现,低质单一的针叶林会带来物种多样性丧失、生产力下降以及土壤营养元素亏缺等生态问题(刘世荣等,2018),营建乡土阔叶林和针阔混交林将成为世界上最有前景的森林经营模式(刘世荣等,2015)。本研究以南亚热带乡土针叶马尾松(Pinus massoniana)和乡土阔叶格木(Erythrophleumfordii)人工纯林及其混交林为研究对象,依据各林分不同土层(0—20、20—40、40—60 cm)土壤样品的宏基因组测序数据,旨在阐明土壤微生物碳循环功能基因丰度对人工混交林以及纯林的响应特征及其调控的土壤环境因子,以期提升对南亚热带人工林土壤微生物碳循环机制的理解,为评价该地区人工林土壤碳循环潜力提供科学参考。
1 材料与方法
1.1 研究区域概况
研究区位于广西区凭祥市的中国林业科学研究院热带林业试验中心青山实验场(22°10′N,106°42′E),属南亚热带湿润半湿润季风气候,4—10 月降雨量最高,年均降雨量高达1 500 mm,年蒸发量高达1 388 mm,相对湿度和年平均温度分别为80%—84%和21.1 ℃;地貌类型主要以低山丘陵为主,土壤类型主要是花岗岩发育形成的山地红壤,坡度25°左右。为探究更优的人工林培育模式,2006年在马尾松林皆伐迹地上分别种植了马尾松-格木混交林及马尾松纯林和格木纯林(密度均为2 500 plant·hm-2),其中混交林中马尾松与格木的株数比为3∶1(罗达等,2014)。
1.2 土壤样品采集
2021 年1 月,分别在上述3 个人工林中随机设置3 块20 m×20 m 的样地,且样地之间至少间隔20 m 以上(避免空间自相关)。马尾松-格木混交林、马尾松林和格木林的保留密度分别为750、1 700 和1 125 plant·hm-2,平均胸径分别为15.6、14.7、13.7 cm,平均树高分别为13.2、11.6、13.7 m。在每个样地采用5 点“S”法确定取样点,清除土壤表面凋落物,使用土钻(内径5 cm)分别采集(0—20、20—40 和40—60 cm)土层土样,将同层土样混合并除去动植物残体与石块等杂质后装入无菌采样袋并用生物冰袋保存带回实验室。将每个土壤样品分为3 份,一份贮存于-80 ℃冰箱用于宏基因组测序;一份贮存于4 ℃冰箱用于测定土壤硝态氮、铵态氮、土壤微生物量碳(MBC)和可溶性有机碳(DOC)含量;一份在室温下风干用于测定土壤pH 值、土壤有机碳(SOC)、全氮(TN)、总磷(TP)、速效磷(AP)以及有机碳组分等含量。
1.3 土壤理化性质与有机碳组分测定
土壤pH 值采用电位法(水土=2.5∶1,V∶m)进行测定(鲁如坤,2000);土壤含水量(SWC)用烘干法测定;土壤有机碳(SOC)采用K2Cr2O7氧化外加热法测定(鲁如坤,2000);全氮(TN)用H2SO4消煮后,用连续流动分析仪(AutoAnalyzer3,SEAL,Germany)测定(鲍士旦,2000);硝态氮(NO3--N)和铵态氮(NH4+-N)使用KCI 浸提(土水比为 1∶5,m∶V),并用连续流动分析仪(AutoAnalyzer3,SEAL,Germany)测定(Wu et al.,2019);总磷(TP)采用H2SO4酸溶-钼锑抗比色法测定(鲍士旦,2000);速效磷(AP)采用双酸(HCl-H2SO4)浸提,用酶标仪(INFINITE M200 PRO,TECAN,Switzerland)测定(鲍士旦,2000)。
微生物生物量碳(MBC)采用氯仿熏蒸法测定(吴金水等,2006);可溶性有机碳(DOC)采用去离子水振荡浸提新鲜土样,用总有机碳分析仪(TOC-VCPH/CPN,Shimadzu,Kyoto,Japan)测定(Huang et al.,2019);易氧化有机碳(EOC)采用高锰酸钾氧化比色法测定(刘合明等,2008);颗粒有机碳(POC)采用(NaPO3)6提取法测定(Zhang et al.,2004);惰性有机碳(ROC)采用酸水解法测定(Rovira et al.,2000)。
1.4 土壤微生物宏基因组测序及碳循环功能基因确定
用FastDNA®Spin Kit for Soil 试剂盒抽提DNA,然后利用1%琼脂糖凝胶电泳检测基因组DNA 质量。检测合格的DNA 样品采用Covaris M220 超声仪将DNA 提取物分割成约400 bp 的小片段,使用NEXTFLEX™ Rapid DNA-Seq Kit 试剂盒构建PE文库,具体步骤及试剂参考试剂盒说明书。宏基因组测序由上海美吉生物技术有限公司 Illumina NovaSeq 测序平台完成。
在测序中采用多个样品平行混合测序,各样品中的序列均引入了一段标示其样本来源信息的Index 标签序列。根据Index 序列区分各个样品的数据,提取出的原始测序数据以fastq 格式保存。基于原始数据,使用Fastp(v0.20.0)对其进行数据质控,剪切掉数据中低质量及含N 的reads,获得后续分析需要的优化序列。将所得优化序列用使用Megahit(v1.1.2)软件进行拼接组装,过滤结果中300 bp 以下的片段。使用Prodigal(v2.6.3)(https://github.com/hyattpd/Prodigal)对拼接结果中的序列重叠群(contig)进行开放阅读框(ORF)预测。将所有样品预测出来的基因序列,用CD-HIT(v4.6.1)软件(http://www.bioinformatics.org/cd-hit/)进行聚类(相似度≥90%、覆盖率≥90%),每个类取最长的基因作为代表序列,构建非冗余基因集。利用SOAPaligner(v2.21)将测序数据与非冗余基因集进行比对(相似度≥95%),统计基因在对应样品中的丰度信息。
使用Diamond(v0.8.35)(https://github.com/bbuchfink/diamond)将非冗余基因集序列与KEGG数据库(v94.2)(http://www.genome.jp/kegg/)进行比对,采用KEGG orthology 对应的碳固定、碳降解和甲烷代谢统计土壤碳循环功能基因丰度。全部土壤样品非冗余基因集的基因数(Catalog gene number)、基因总序列长度(Catalog total length)和基因平均序列长度(Catalog average length)分别为5 518 361 个、2 241 669 993 bp 和406.22 bp,获取微生物碳固定、碳降解和甲烷代谢等功能基因674 个。选取各碳循环途径基因丰度大于10 000 的功能基因进行分析(表1)。
1.5 统计分析
采用双因素方差分析方法(Two-way ANOVA)检测不同林分不同土层的土壤理化性质、土壤有机碳组分含量及碳循环功能基因丰度的差异,并用Duncan 法进行多重比较,计算由SPSS 26.0(IBM SPSS Inc,Chicago,IL,USA)完成。使用R 软件psych 包中“corrlot ( )”函数对土壤环境因子与碳循环功能基因丰度之间进行Pearson 相关性分析。
2 结果与分析
2.1 不同林分不同土层的土壤理化性质
由图1 可知,相同土层不同林分相比较,0—20 cm 土层马尾松林TP 含量显著高于格木林和马尾松-格木混交林;20—40 cm 土层马尾松林和马尾松-格木混交林土壤TP 含量无显著差异,且均显著高于格木林(P<0.05)。
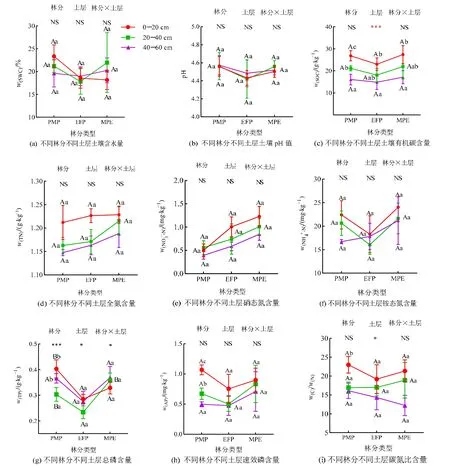
图1 不同林分不同土层之间土壤理化性质的比较Figure 1 Comparison of soil physicochemical properties between different soil layers and forest stands
相同林分不同土层相比较,马尾松林SOC、AP含量和C/N 随土壤深度增加而显著减少(P<0.05)。马尾松林0—20 cm 和40—60 cm 土层TP 含量无显著差异,且均显著高于20—40 cm 土层。格木林与马尾松-格木混交林0—20 cm 土层的SOC 含量与20—40 cm 土层差异不显著,但显著高于40—60 cm土层。双因素方差分析表明,林分和林分与土层的交互作用对土壤TP 含量具有显著影响,土层对土壤SOC、TP 含量和C/N 有显著作用(P<0.05)。
2.2 不同林分不同土层的土壤有机碳组分
从表2 可知,不同林分土壤有机碳组分含量存在显著差异(P<0.05)。0—20 cm 土层格木林的MBC含量显著高于马尾松林,而POC 含量显著高于马尾松-格木混交林;马尾松-格木混交林的EOC 含量显著高于马尾松林,而显著低于格木林。20—40 cm土层中格木林的POC 含量显著高于另外两个林分,而土壤ROC 含量从马尾松林到格木林再到混交林显著增加。40—60 cm 土层中格木林的MBC 和POC含量显著高于马尾松林和混交林。

表2 不同林分不同土层土壤有机碳组分的比较Table 2 Comparison of soil organic carbon components between different soil layers and forest stands
比较相同林分不同土层土壤有机碳组分发现,马尾松林中0—40 cm 土层的MBC 含量显著高于40—60 cm 土层,而POC 和ROC 含量在0—20 cm土层显著最高。格木林中0—20 cm 土层MBC 含量显著高于40—60 cm 土层,与20—40 cm 土层差异不显著,且0—20 cm 土层EOC 和ROC 含量显著高于20—60 cm 土层;同时,0—40 cm 土层格木林的POC 含量和混交林的MBC 含量显著高于40—60 cm 土层。另外,混交林中0—20 cm 土层EOC和POC 含量显著高于20—60 cm 土层,而20—40 cm 土层ROC 含量显著高于40—60 cm 土层,与0—20 cm 土层差异不显著。双因素方差分析表明,林分对MBC、EOC、POC 和ROC 含量有显著影响,土层对土壤有机碳组分均具有显著影响,而林分与土层对EOC、POC 和ROC 含量有显著交互效应(P<0.05)。
2.3 不同林分不同土层的土壤微生物碳循环功能基因丰度
2.3.1 碳固定功能基因丰度
从图2 可知,马尾松林各土层的MUT基因丰度均显著高于格木林(P<0.05),而20—40 cm 土层马尾松林的rbcL、PCCA基因丰度分别显著高于马尾松-格木混交林及格木林。

图2 不同林分不同土层碳固定功能基因丰度Figure 2 Abundance of carbon fixation genes in different stands and soil layers
相同林分不同土层碳固定功能基因丰度的结果表明,马尾松林rbcL和accA基因丰度随土壤深度增加而显著增加(P<0.05);同时,马尾松林20—60 cm 土层的acs和korA显著高于0—20 cm 土层,而40—60 cm 土层korB显著高于0—20 cm 土层。格木林20—60 cm 土层的rbcL显著高于0—20 cm 土层,而40—60 cm 土层的acsA显著高于0—20 cm 土层,以及40—60 cm 土层accA显著高于0—40 cm 土层。混交林各土层功能基因丰度差异不显著。总之,3 个林分碳固定功能基因丰度对土壤深度的响应趋势相似,深层土壤碳固定功能基因丰度相对较高。双因素方差分析表明,林分显著影响rbcL、MUT和PCCA,土层显著影响rbcL、acs、acsA、korA、korB和accA,而林分与土层对碳固定功能基因丰度无显著交互作用(图2)
2.3.2 碳降解功能基因丰度
双因素方差分析表明,林分对amyA、MAN2C1、bglB和nagA有显著作用,土层对amyA、pulA、malQ、xylB、MAN2C1、xylA、endoglucanase、bglB、bglX、nagA和catE有显著影响,林分和土层无显著交互作用(图3)。相同土层不同林分微生物碳降解功能基因丰度的比较发现(图3),0—20 cm土层马尾松林与格木林的MAN2C1无显著差异,且都显著高于混交林(P<0.05);而马尾松林bglB显著高于混交林,与格木林差异不显著。20—40 cm 土层各林分间碳降解功能基因丰度无显著差异。40—60 cm 土层混交林amyA显著高于格木林,而马尾松林nagA显著高于格木林。
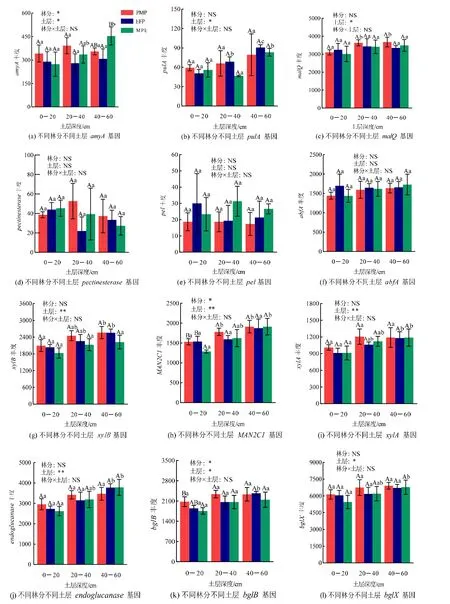
图3 不同林分不同土层碳降解功能基因丰度Figure 3 Abundance of carbon degradation functional genes in different forest stands and soil layers
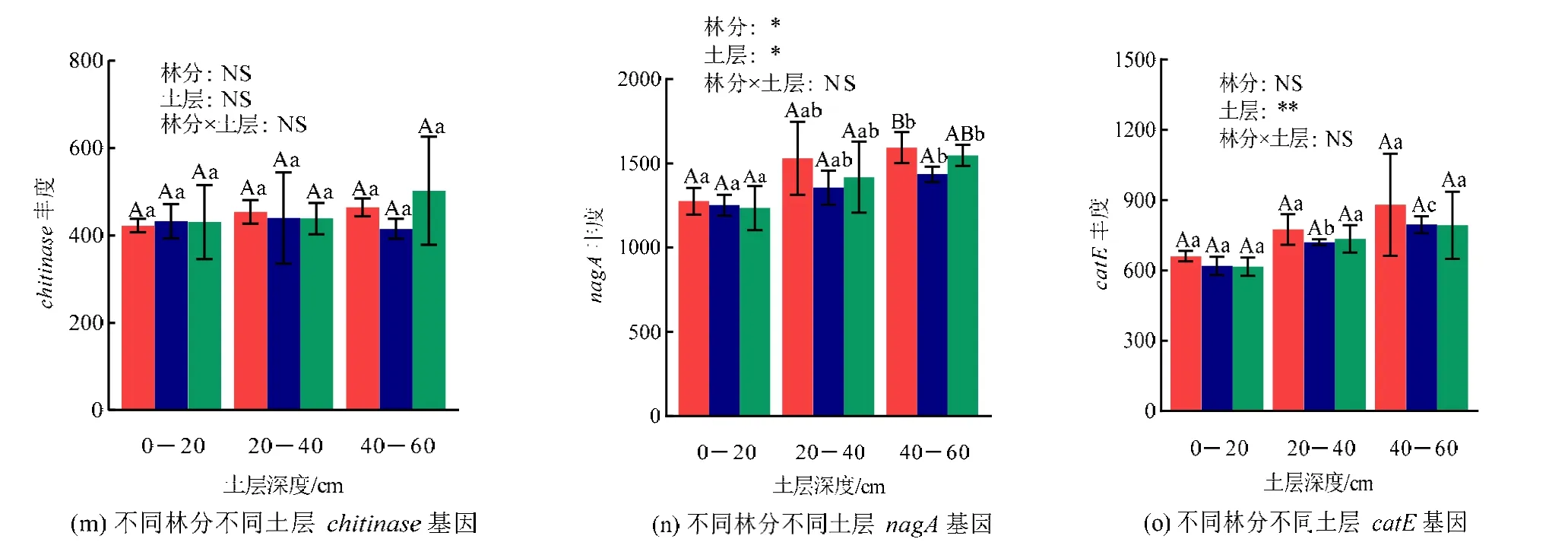
续图3 不同林分不同土层碳降解功能基因丰度Continued figure 3 Abundance of carbon degradation functional genes in different forest stands and soil layers
从图3 可知,就相同林分不同土层而言,马尾松林20—60 cm 土层malQ和MAN2C1基因丰度显著高于0—20 cm 土层,而40—60 cm 土层xylB和nagA显著高于0—20 cm 土层。格木林pulA和catE基因丰度随土壤深度加深而显著增加,而40—60 cm 土层xylB、xylA和nagA显著高于0—20 cm 土层;另外,格木林40—60 cm 土层endoglucanase和bglB显著高于0—40 cm 土层。混交林40—60 cm土层amyA、MAN2C1、xylA、endoglucanase、bglX和nagA显著高于0—20 cm 土层,且0—40 cm 土层的pulA显著低于40—60 cm 土层。
2.3.3 甲烷代谢功能基因丰度
从图4 可知,对于相同土层不同林分来说,0—20 cm 土层马尾松林pomA-amoA基因丰度显著高于格木林和马尾松-格木混交林(P<0.05);20—40 cm土层格木林mtrA显著高于另外两个林分;40—60 cm 土层马尾松林pomB-amoB和pomC-amoC显著高于混交林,与格木林差异不显著。比较相同林分不同土层发现,马尾松林中40—60 cm 土层pomAamoA显著高于20—40 cm 土层,而pomB-amoB随土壤深度增加而显著增加;此外,马尾松林40—60 cm 土层pomC-amoC显著高于0—20 cm 土层。格木林0—40 cm 土层pomC-amoC显著低于40—60 cm 土层。混交林20—60 cm 土层mtrA显著高于0—20 cm 土层。双因素方差分析结果显示,林分和土层显著影响pomA-amoA、pomB-amoB、pomCamoC和mtrA,而林分与土层交互作用对甲烷代谢功能基因丰度无显著影响。
2.4 土壤碳循环微生物功能基因与土壤环境因子的相关性
碳固定功能基因rbcL、acs、acsA、korA和accA与C/N、SOC、MBC、DOC、EOC 和ROC 显著负相关(P<0.05);MUT与SWC 和TP 显著正相关,与NO3--N、MBC、EOC、POC 和ROC 显著负相关,PCCA与TP 显著正相关(图5a)。同时,碳降解功能基因amyA、abfA和xylA与SWC 显著正相关,xylB、MAN2C1、xylA、endoglucanase、bglB、bglX、nagA和catE则与土壤SOC、MBC、DOC、EOC 和ROC 显著负相关(P<0.05)(图5b)。另外,甲烷代谢功能基因pmoA-amoA、pmoB-amoB和pmoC-amoC与土壤MBC、DOC 和EOC 显著负相关,而mtrA与土壤NH4+-N、C/N、SOC 和ROC 显著负相关(P<0.05)(图5c)。
3 讨论
3.1 林分类型及土壤深度对微生物碳固定功能基因丰度的影响
土壤微生物碳循环相关功能基因丰度的不同,会影响微生物群落的碳代谢功能,进而影响土壤碳循环(Ding et al.,2015)。森林生态系统中,土壤微生物碳循环基因丰度与植被类型和土壤性质密切相关(Yue et al.,2015;Deng et al.,2019;李毳等,2020)。土壤微生物碳固定是在多种固定途径下,微生物将碳源转化为自身物质的过程(碳同化),能够促进土壤碳增汇和碳固持(袁红朝等,2011;陈晓娟等,2014)。微生物碳固定基因丰度指示微生物的固碳潜力,基因丰度越高相应的固碳潜力越强,对土壤碳汇贡献越大(刘茗等,2021)。本研究发现,马尾松林各土层的MUT丰度均显著高于格木林,以及20—40 cm 土层的PCCA显著高于格木林,这意味着马尾松林土壤(0—60 cm)微生物碳固定潜力强于格木林。相关性分析显示,研究林分MUT丰度与SWC 和TP 显著正相关,与NO3--N、MBC、EOC、POC 和ROC 显著负相关,而PCCA与TP 显著正相关(图5a),说明微生物碳固定功能基因丰度与土壤养分和有机碳组分密切相关。因此,格木林土壤MUT和PCCA丰度显著低于马尾松林可能是因TP 含量较低所致;同时,格木林MUT丰度低还与土壤中丰富的NO3--N、MBC、EOC、DOC 和ROC 抑制有关。已有研究指出,在贫瘠、低碳土壤环境中,仍有微生物具有较强的固碳潜力,促进土壤碳固定,从而导致土壤SOC、DOC、MBC 和ROC 对固碳细菌丰度的负影响(Wang et al.,2021)。
深层土壤(30 cm 以下)碳储量占土壤总碳储量的一半以上,具有较高的固碳潜力和稳定性(Liao et al.,2020)。本研究各林分微生物碳固定功能基因丰度基本上随土层深度增加而增加(图2),表明深层土壤对森林生态系统固碳潜力较大,有助于土壤碳积累。同时,本研究各林分土壤C/N、SOC、MBC、POC 和EOC 随土层深度加深而减少,这与前人的研究结果一致(Peng et al.,2016;梁艳等,2021;宋瑞朋等,2022);而研究林分的微生物碳固定功能基因丰度与上述土壤环境因子显著负相关(图5a),因此,当土壤养分与有机碳组分含量随土层深度增加而减少时,导致碳固定功能基因丰度增加。另外,也有研究认为表层土壤因氧浓度较高而导致一些固碳细菌的生长受到抑制,引起表层土壤微生物固碳潜力较弱(Wang et al.,2021)。
3.2 林分类型及土壤深度对微生物碳降解功能基因丰度的影响
土壤微生物碳降解是指微生物对易降解(淀粉)和难降解(果胶、半纤维素、纤维素、几丁质、木质素等)等碳源化合物的分解,是碳素释放和迁移转化的过程,推动碳的生物地化循环(Sokol et al.,2019)。土壤微生物碳降解基因相对丰度随植被类型变化而异(Yan et al.,2020)。Wang et al.(2020)研究发现,中国南方天然次生林碳降解基因丰度高于马尾松和湿地松(Pinuselliotti)人工林,以及稳定性有机碳组分、有效氮和硝态氮是重要的影响因子。本研究中马尾松林0—20 cm 土层半纤维素降解功能基因MAN2C1和纤维素降解功能基因bglB的丰度显著高于马尾松-格木混交林(图3),这是由于马尾松林土壤SOC、MBC、EOC 含量低,而MAN2C1和bglB基因丰度与SOC、MBC、DOC、EOC 和ROC 显著负相关(P<0.05)(图5b)。土壤微生物在有机碳充足和养分丰富的条件下,因自身生长速率加快而加速对碳源的利用,导致碳的降解潜力降低(Ma et al.,2022)。已有研究也表明土壤MBC 和DOC 等有机碳组分与土壤碳降解基因丰度呈负相关(Coolen et al.,2015;Xue et al.,2016),较低的SOC 促进了微生物碳降解酶活性(Liao et al.,2023)。此外,本研究3 个林分20—40 cm 土层的碳降解功能基因丰度差异不显著,可能与林分间土壤SOC、MBC、DOC 和EOC 含量无显著差别有关。混交林40—60 cm 土层的amyA丰度显著高于格木林(图3),可能是该土层格木林有较高的MBC 和POC 含量以及较低的SWC 含量,这是由于amyA与SWC 和TP 显著正相关,而与MBC、EOC 和POC 显著负相关(图5b)。Li et al.(2017)发现水分增加通过增加微生物碳降解功能基因丰度,加速半干旱地区的碳循环过程。
碳降解是土壤碳流失的主要过程(Yang et al.,2023)。本研究发现,3 个林分土壤微生物碳降解功能基因丰度与碳固定功能基因丰度具有相似的土壤垂直变化趋势。由于碳降解功能基因丰度基本上与土壤有机碳组分含量显著负相关(图5b),因而当碳降解功能基因丰度随土层深度增加而升高后,土壤中有机质的分解速率加快,可造成土壤碳损失。
3.3 林分类型及土壤深度对微生物甲烷代谢功能基因丰度的影响
甲烷代谢包括甲烷生成和甲烷氧化过程,在产甲烷菌和甲烷氧化菌共同参与下进行,控制着甲烷产生、氧化和陆地生态系统的甲烷平衡(刘洋荧等,2017)。本研究中马尾松林0—20 cm 土层的甲烷氧化功能基因pmoA-amoA丰度以及40—60 cm 土层的pmoB-amoB和pmoC-amoC丰度显著高于混交林(图4),说明马尾松林土壤甲烷氧化速率高于混交林。相关性分析表明,pmoA-amoA、pmoB-amoB和pmoC-amoC都与土壤MBC、DOC 和EOC 显著负相关(图5c),意味较高的土壤有机碳组分含量对甲烷氧化功能基因丰度具有一定的抑制作用。另外,格木林20—40 cm 土层的甲烷生成功能基因mtrA丰度最高,且与SOC、NH4+-N、C/N 和ROC显著负相关(图5c),说明格木林土壤SOC、NH4+-N、C/N 和ROC 含量较高时会使mtrA丰度降低。Luo et al.(2022)研究发现土壤DOC 对pmoA丰度具有负影响;赵姣等(2023)研究指出,黄土高原植被恢复土壤甲烷生成功能基因(mxa、emGDH)丰度与SOC、EOC 显著负相关,而甲烷氧化功能基因(pmoA、mmoX)丰度与EOC、TN 和NO3--N 含量显著负相关;Kosugi et al.(2020)认为土壤SOC含量与产甲烷菌的甲烷生产速率具有负相关关系。可见,本研究对林分土壤甲烷生成与氧化微生物功能基因丰度产生负效应的主导因素与上述研究结果类似。
本研究3 个林分土壤微生物甲烷代谢功能基因丰度与SOC、NH4+-N、C/N、MBC、DOC、EOC 和ROC 显著负相关(图5c),主要是因为甲烷代谢微生物种群复杂,一些生活在贫营养(低氧和低氮)环境中的优势菌氧化潜力较强,相应的基因丰度也较高(Zheng et al.,2010)。因此,当土壤养分和有机碳组分含量随土壤深度加深而减少时,甲烷代谢功能基因丰度增加。此外,甲烷作为有机质厌氧分解的最终产物,通常在厌氧条件下产生,深层土壤因氧浓度较低产甲烷菌代谢作用更强(王芸等,2012),从而造成土壤微生物甲烷代谢功能基因丰度与有机碳组分及养分含量负相关。
4 结论
南亚热带马尾松林土壤微生物碳固定(rcbL、MUT和PCCA)、碳降解(MAN2C1和bglB)和甲烷代谢(pmoA-amoA、pmoB-amoB和pmoC-amoC)功能基因丰度均高于马尾松-格木混交林和格木林,表征马尾松林土壤微生物碳循环潜力较强,这主要与马尾松林土壤MBC、EOC、DOC 和ROC 含量较低密切相关。3 个人工林土壤微生物碳循环功能基因丰度基本随土壤深度增加而增加,是因为林分土壤SOC、C/N、MBC、DOC、EOC 和ROC 含量随土壤深度加深而降低的缘故。因此,土壤有机碳组分对该地区马尾松-格木人工混交林及其纯林土壤微生物碳循环功能基因丰度具有负效应。
