Antipredatory call behavior of lapwing species in an Afrotropical environment
2024-01-22FtimJmesChiomOkforSmuelOsinubiShiiwuMnuSmuelIvndeTiwoOmotoriogun
Ftim R.Jmes, Chiom I.Okfor, Smuel T.Osinubi, Shiiwu A.Mnu,Smuel Ivnde, Tiwo C.Omotoriogun,e,f,*
aA.P.Leventis Ornithological Research Institute, University of Jos, P.O.Box 13404, Jos, Plateau State, Nigeria
bEden Creation Care Initiative Rennajj Fish Farm, P.O.Box 7173, Jos, Plateau State, Nigeria
cFitzPatrick Institute of African Ornithology, University of Cape Town, Cape Town, South Africa
dDepartment of Zoology, University of Jos, Jos, Plateau State, Nigeria
eBiotechnology Unit, Department of Biological Sciences, Elizade University, P.M.B.002, Ilara-Mokin, Ondo State, Nigeria
fNatural History Museum, University of Oslo, P.O.Box 1172 Blindern, 0318 Oslo, Norway
Keywords:Acoustic Alarm calls Lapwings Mobbing calls Vocal signals
ABSTRACTPredation is an important source of natural selection on prey species and has resulted in adaptations such as antipredator vocal signals, which can alert others to the presence of predators and solicit cooperative attack.Although vocal alarm signals of birds have been well studied, they are poorly known in tropical African species.To address this lack of information, the antipredatory signals and responses of two lapwings (Wattled Lapwing Vanellus senegallus and Spur-winged Lapwing Vanellus spinosus)to potential predators were investigated using data collected from focal observation,distance measurements,focal recordings,and playback experiment.The lapwing calls elicited to predators were classified as alarm or mobbing calls based on whether the calls elicited alert behavior or attack from other lapwings.Discriminant linear analysis (DLA) was used to compare the time and frequency parameters of the call types measured in Raven PRO.Also, lapwings’ responses to intruders, alert and start distance, time of day, and latency, as well as the effects of flock size and distance to cover were examined.About 48% of all calls was correctly classified by DLA.The best predictors of call type for the lapwings were maximum frequency and high frequency.Both alarm and mobbing calls were elicited by African Wattled Lapwings to dogs and humans.Mobbing calls were elicited to intruders by the Spur-winged Lapwings.Alert distance was positively associated with start distance, and differed between morning and evening in both lapwings.With scarce information from tropical Africa,this study put in perspective vocal and antipredator behavior of lapwing species in Africa.
1.Introduction
Predation is an important source of mortality in prey species (Palkovacs and Post,2008;Reidy et al.,2009;Johnson and Belk,2020)and has resulted in adaptations to reduce its impact (Elliot, 1985; Ruxton et al., 2004; Caro, 2005; Jeschke et al., 2008; Stevens and Merilaita,2008; Cooper et al., 2015).Among the myriad of adaptations are mechanical, chemical, physical, and behavioral antipredator defenses such as camouflage(e.g.,butterfly,Bohlin et al.,2008);masquerade(e.g.,fish,Buresch et al., 2011; plants, Lev-Yadum, 2014); alert behavior (e.g.,lizards, Cooper et al.,2010); and vocalizations(e.g., birds,Krams et al.,2006;Magrath et al.,2014).
Vocalizations are among the best studied antipredator adaptations(Klump, 2000; Hollen and Radford, 2009; Garcia and Favaro, 2017).Many species of prey call to alert others of the presence of a predator(Wheeler, 2008; Magrath et al., 2014).In response, alerted individuals can increase vigilance (Griesser, 2008), depart the area (Forsman and M€onkk€onen, 2001), or actively engage the predator (Krams and Krama,2002).There are reports that antipredator calls elicited reflect the category of predator (Van Buskirk, 2001; Templeton et al., 2005; Botham et al.,2006;Eccard and Yl€onen,2007;Seyfarth and Cheney,2010).
Vocal antipredator signals have been especially well studied in birds(Yorzinski and Platt, 2012).For example, these calls can elicit mobbing behavior in which prey individuals aggressively attack predators to drive them from the area (Arnold, 2000; Krams and Krama, 2002; Yorzinski and Vehrencamp, 2009).Mobbing calls are typically loud and easy to locate, and their frequencies may increase during the breeding season(Kelvin and Jennifer, 2011).Hypotheses for the functions of mobbing behavior include, the ‘lethal counterattack’; ‘move-on’; ‘silencing offspring’; ‘alerting others’; ‘selfish herd’; ‘attract the mightier’; ‘aiding the distressed relative’hypotheses(Krams and Krama,2002;Caro,2005;Griesser and Ekman, 2005).In contrast, antipredator vocalizations can serve to alert others about danger (Klump, 2000; Carl and Michael,2000).These alarm calls are high frequency, structurally simple sounds that are difficult to localize (Carl and Michael, 2000).Alarm calls also signal to predator of its detection (‘perception advertisement’ hypothesis), as well as expresses prey's ability for escape (‘condition advertisement’hypothesis)(Caro,2005).
Although antipredator vocal behavior has been studied in many species of birds (Elliot, 1985; Kis et al.,2000;Krams et al.,2006;Evans and Evans,2007;Brynychova et al.,2022),there is far more to be known about this behavior in bird species especially within the Afrotropical environment (Nichols and Yorzinski, 2016).There are some general life-history differences between birds from the tropics and those from other parts(e.g.,temperate zones)of the globe(Stutchbury and Morton,2008).Most tropical birds have characteristically low adult mortality and reduced annual reproductive rates small clutch sizes and slow pace of life(Peach et al., 2001; Ricklefs and Wikelski, 2002; Wiersma et al., 2007).Also, the tropical environment is highly diverse (Wiersma et al., 2007),with seasonal environmental conditions and relatively constant day length and warm temperature.Studies have shown increased evolutionary speed (Rohde, 1992; Allen et al., 2002), and faster speciation rates in the tropics (Schemske, 2002).Furthermore, variation in frequencies and call-based parameters of antipredatory vocal signals are poorly known in Afrotropical birds, despite considerable evidence of intraspecific differences in the accuracy of signalling behavior of some species (Ridley and Raihani, 2007; Flower et al., 2014; Silvestri et al.,2019).One exception is Walters (1990), who studied three species of lapwings in South America and Africa and showed that lapwings discriminate the various kinds of predators and non-predators they encounter and that responses to each correspond to the type and degree of danger each represents.In general, however, studies that investigate responses to predators or intruders(whether human or nonhuman),and the proximity to which prey can tolerate an advancing predator before producing vocal signals,fleeing,or engaging in other appropriate forms of defense measures,are severely lacking.In addition,information on the effects of time of day, group size, and nearness to cover on signalling behavior of birds is scarce.
To address the limited knowledge of antipredator behavior of Afrotropical birds,this study investigated variation in the antipredatory calls and responses of two species of lapwing.The specific objectives were to:1) classify antipredatory call types as mobbing and alarm calls; 2)discriminate between mobbing and alarm calls using their call parameters;3)compare the call rates of the two lapwings in response to human and nonhuman predators; 4) determine the relationship between alert and start distance and the effect of time of day on alert distance and latency on the call types in the lapwing species;5)determine the effects of flock size and distance to cover on the antipredatory calls of the lapwing species.
2.Materials and methods
2.1.Study area and site locations
The study was conducted in Jos Plateau State Nigeria, located at an altitude of ~1200 m above sea level.The area experiences wet (May-–October)and dry(November–April)seasons with average annual rainfall of~1400 mm and a temperature range between 10 and 35 C(<10 C in coldest months and 35 C during warm and dry months; Payne,1998).Ten sites were used for the study:Aningo River(839ʹN,858ʹE),Gwash Dam(990ʹN,895ʹE),Gwafan(953ʹN,856ʹE),Kwanga(993ʹN,888ʹE),Lamingo Dam(989ʹN,893ʹE),Laminga(953ʹN,859ʹE),Shen Dam(946ʹN,855ʹE),Zarazong Dam(955ʹN,854ʹE),Bayan Dutse(9 52'N, 857'E), and Rennajj Fish Farm (982ʹN, 889ʹE).The sites were within the open savannah habitats, characterized by farmlands, human settlements,rocky outcrop,and water bodies.
2.2.Study species
The study was focused on the African Wattled Lapwing (AFWLA,Vanellus senegallus)and Spur-winged Lapwing(SPWLA,Vanellus spinosus)(Family: Charadriidae; Subfamily: Vanellinae) due to easy access to the species, conspicuous and charismatic behavior.Both species are largesized, Afrotropical resident birds with partially migratory and erratic behavior within range (Borrow and Demey, 2014).They are sexually monomorphic (except the females of AFWLA has less black on its head and throat, and the SPWLA have shorter spurs) compared to their respective males.The two species are monogamous, solitary and sometimes in small parties of four pairs.Both species breed in the study area during the early rainy season (March–June) with nests constructed on bare ground,and containing 2–3 eggs that are highly prone to predation from humans and nonhuman species(del Hoyo et al.,1996;Borrow and Demey, 2014).The species maintain and defend territories aggressively against predators and other intruders; and occur mainly in open and lowland habitats, damp grassland, grassy edges of lakes and rivers,marshes, and lightly wooded grassland, including open grasslands and cultivated farmlands near water bodies.The species mostly feed on insects and larvae, but occasionally feed on crustaceans, molluscs, small lizards, tadpoles and adult frogs (del Hoyo et al., 1996; Borrow and Demey,2014).
2.3.Field survey and data collection
The field survey and data collection occurred from May to July 2016 and consisted of four major aspects: focal observations, distance measurements, focal recordings, and playback experiment which were carried out in the mornings(06:30–10:00 GMT)and evenings(15:00–18:00 GMT).All distance measurements were performed using laser range finder (Nikon Laser-800; Nikon, Tokyo, Japan); and coordinates were taken using a GPS device.
2.4.Focal observation
The study sites were visited for the presence, identification and observation of the AFWLA and SPWLA.Thirty minutes observation of the species was undertaken using a telescope and a pair of binoculars.The antipredatory call behaviors (vocal sound categorized as alarm or mobbing calls)elicited against the presence of non-human intruders such as cattle,dog and other bird species,were recorded.We distinguished the calls (alarm and mobbing) based on the activities of the birds during a call.If the birds elicit a call while fleeing from a predator/intruder,it was classified as alarm calls.Where the birds elicit a call while attacking an intruder or predator, it was classified as mobbing call.Birds were breeding during the survey and the activities of birds such a incubation,brooding, foraging and preening were recorded.Birds were not marked to avoid interference with breeding activities.However,we relied on the territorial behavior, and consistent availability of the pair at the site during the monitoring period.The sexes or individuals of the pair were not differentiated;hence,any individual of the pair was observed during the survey.
2.5.Alert distance measurement
After each focal observation, alert distance experiment was performed to determine the distance of intruders to lapwing before eliciting response call.The human subject used as the intruders wore the samecolor of clothes for all approaches to reduce bias that might be due to differences in intruders’to detect different colors.The subject advanced towards the birds from a start distance(i.e.,distance between the subject and the bird before the advancement towards the birds) at a steady walking pace.The call elicited and proximity of the focal birds from the human intruder when the call was elicited was recorded.Alert distance was calculated as the distance between the intruder and the focal bird when it detected the approaching intruder and elicited the first call.Alert distance is considered a better measurement than other measurements like flight initiation distance(the distance at which a bird initiates flight when approached by a potential predator)when measuring the minimum approaching distance of birds.This is because it allows the birds to adapt their responses to intruders(Fernandez-Juricic and Jimenez,2001).
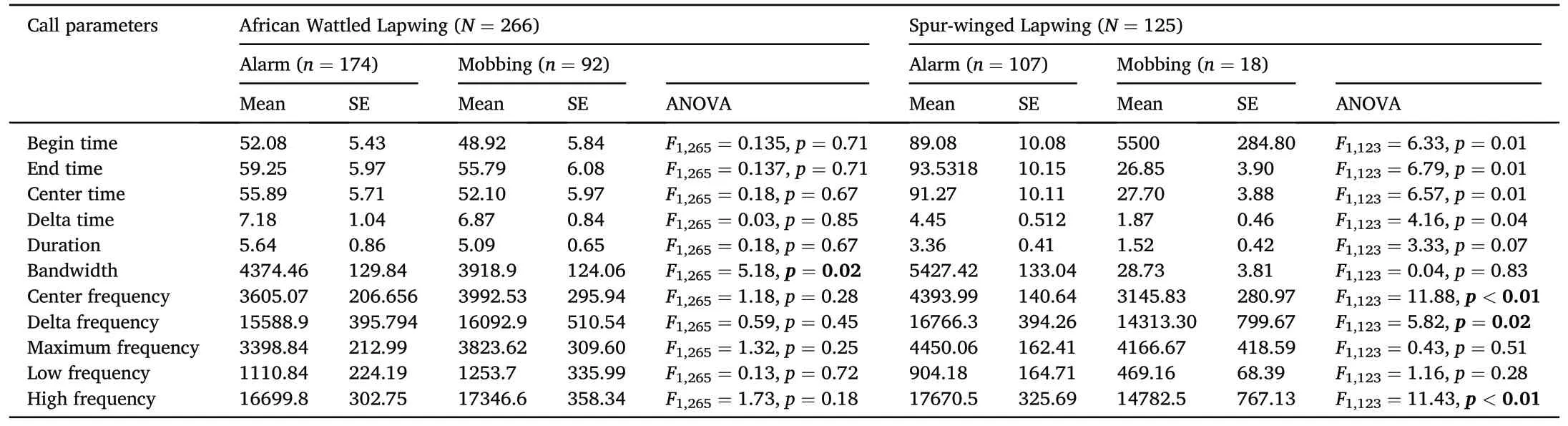
Table 1Means and standard error(SE)including ANOVA test of the time(s)and frequency(Hz)call parameters for the antipredatory call types(mobbing and alarm)of the two lapwing species (African Wattle Lapwing and Spur-winged Lapwing) in tropical African environment.
2.6.Focal recording experiment
The calls of the lapwings were recorded using a digital recorder(Marantz PMD661, D&M Holdings Inc., Kanagawa,Japan) fitted with a shot-gun microphone (ME67/K6, Sennheiser, Germany).The unidirectional shot-gun microphone was pointed in the direction of the birds identified either as AFWLA or SPWLA.
2.7.Playback experiment
Alarm and mobbing calls (from another individual of AFWLA or SPWLA, i.e., obtained from the African Bird Sounds Collections) were played to focal AFWLA or SPWLA using playback.The response (either alarm or mobbing calls)of the birds,the call latency(s),distance of the speaker from the bird,and group size during the playback were recorded.The playback was performed using a WAV player (PMD661MK II Handheld Solid-State Recorder) connected to a JBL GO portable Bluetooth speaker set to maximum volume.
2.8.Data analysis
Sound files from the digital recorder were transferred and interpreted in Raven Pro version 1.5 (www.birds.cornell.edu/raven; Charif et al.,2008).Call-based parameters which best described the characteristics of each call were extracted and used for comparative analysis.The parameters include the following: high and low frequencies–the lower and highest frequencies bound of the calls; center frequency–the frequency that divides the selection into two frequency intervals of equal energy or the smallest discrete frequency within the calls which satisfies the inequality; maximum frequency–the frequency at which maximum power, peak power occurs within the calls; delta frequency–the difference between the upper and lower frequency limits; bandwidth–the difference between the 1st and 3rd quartile frequencies; begin and end time–the start and end time of selection; Delta time–the difference between begin time and end time for the selection;center time–the point in time at which the selection is divided into two-time intervals of equal energy.
The extracted parameters were discriminated and classify into the antipredatory calls(mobbing and alarm)of the two species using a Linear Discriminant Analysis(LDA)implemented in the Software PAST version 3.13 (Hammer et al., 2016).The axes were linear combinations of the original variables with eigen values,to produce first maximal and second maximal separations between the call groups as the amount of variation explained by the axes.The clusters of the call types were visualized using a scattergram projected in two-dimensional discriminant axes space.The discriminant of the call type was also assessed for each species separately.The data was classified by assigning each point to the group that gives minimal Mahalanobis distance to the group mean.The Mahalanobis distance was calculated from the pooled within-group covariance matrix to give a linear discriminant classifier.The group assignment procedure was by a leave-one-out cross-validation (the Jackknifing procedure).
Variation in the frequency and time-based parameters of the calltypes (mobbing and alarm calls) of the species was tested using multivariate analysis in R version 2.15.2 (R development core team, 2018).The variation in the call parameters between and within call-types were compared using Mann–Whitney U (unpaired) and Wilcoxon Sign Rank(paired) respectively.The hypothesis that the frequencies of call-types were independent of the category of intruders was tested using a Chi-square test of dependence.
Next,a General Linear Model(‘lmer’function of the‘lme4’package;Bates et al.,2015)was used to determine the effects of flock size,distance to vegetation cover and time of day on the antipredatory call response of the lapwings.Further, alert distance was tested as a function of start distance;distance to cover,flock size,and species were used as response variables; and session and sites were used as random variables.The‘drop1’function(model 1,~,test¼“Chi”)was used to get the P-values by creating and comparing two models for each variable (one with the variable of interest,and the other without it)to choose the best model.
3.Results
In total, 130 lapwings (AFWLA, n ¼ 63; SPWLA, n ¼ 67) were observed as individuals or in a group (Appendix A); and 81 lapwings(AFWLA, n ¼ 35; and SPWLA, n ¼ 47) were approached during the antipredator call response experiment (Appendix B).The mean and standard error of the parameters (time and frequency) of the call type analyzed for 391 (Appendix C) extracted from Raven are shown in Table 1.The spectrogram of the call types from each species of lapwing is shown in Fig.1.
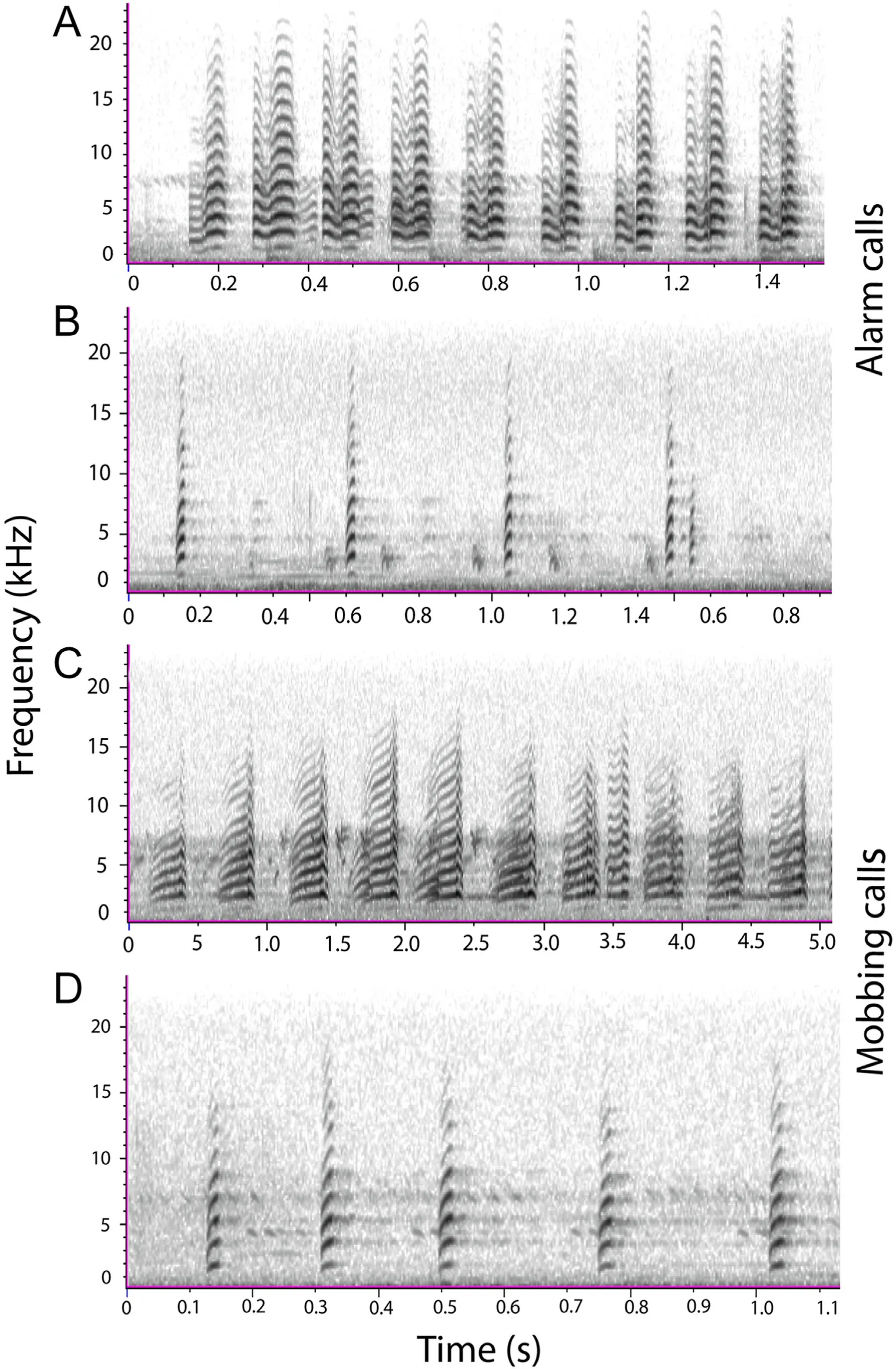
Fig.1.Spectrogram of antipredatory call signals from Raven Pro showing the frequency (kHz) of for the alarm (A) and mobbing (C) calls for the African Wattled Lapwing; and alarm (B) and mobbing (D) calls for the Spur-winged Lapwing.
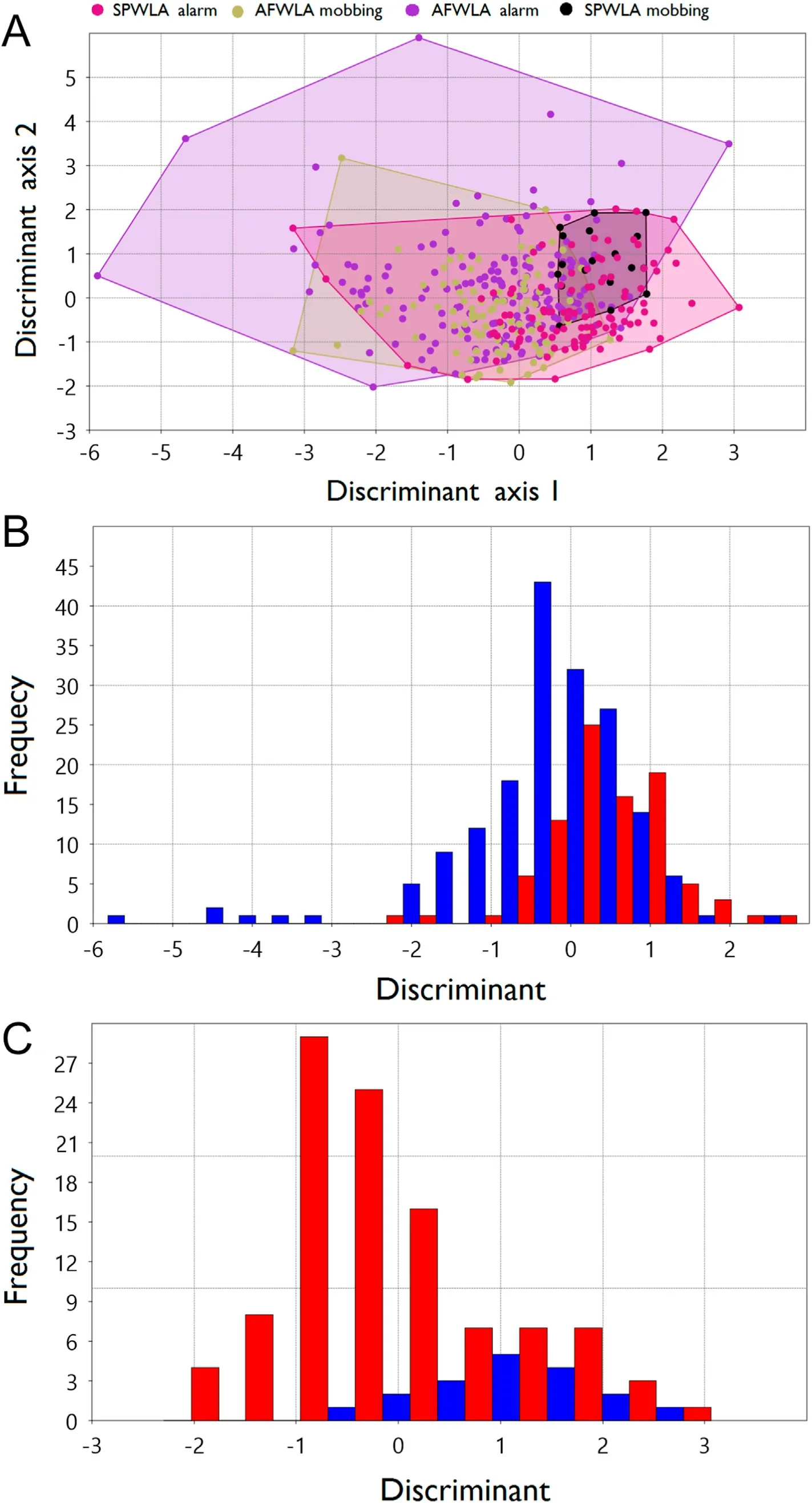
Fig.2.(A)The scattergram with convex hulls of the antipredatory call types(alarm and mobbing)with calls classified using Jacknife procedure and projected into a two-dimensional linear discriminant axis.Individual species discriminant of the call types for the African Wattled Lapwing (B) and Spur-winged Lapwing (C).

Table 2Variation (ANOVA) in the frequencies of the call parameters, and pairwise comparison for difference in the call types (alarm and mobbing) elicited by the lapwing species.

Fig.3.The rate of alarm calls (green) and mobbing (grey) in the African Wattled Lapwing (AFWALA) and the Spur-winged Lapwing (SPWLA); mobbing call was more frequently elicited than alarm call in both species of Lapwing.
Linear Discriminant Analysis of call parameters showed two significant DLA functions: axis 1 and axis 2 explaining 72%and 18% of the variance respectively between call types of the species of lapwing.Forty-eight percent of all calls was correctly discriminated between the call types.Overall,the clusters of the call types did not reveal any clearcut distinction in the antipredatory call type of the AFWLA and SPWLA as shown in Fig.2A.There was evidence that mobbing call between the two species can be distinguished except for an overlapping region between the calls(see Fig.2A).
The best predictors of the call types for the lapwings were maximum frequency (Wilks λ ¼ 0.00057), and high frequency (Wilks λ ¼ 0.0004)respectively (Table 2).There was significant variation in maximum frequency(one way ANOVA:F2,388¼208.9,p<0.001)and high frequency(one way ANOVA: F2,388¼ 208.9, p < 0.001).The first discriminant linear axis indicates calls located at the negative scores along the discriminant axis had higher values of the maximum and high frequencies than those of the positive axis.This also explains the similarity seen in the cluster of call locations along that axis (Fig.2A for the call clusters).There was also significant variation in high frequency(one way ANOVA:F2,388¼818.1,p<0.001).The second discriminant linear axis indicates that the calls located along the positive scores have higher frequency than those located at the negative scores.
The outcome of discrimination and classification of the call types showed 63.53%and 77.2%for AFWLA and SPWLA respectively(Fig.2B and C).There was significant variation in frequencies for the alarm and mobbing calls in bandwidth (F1,265¼ 5.182, p ¼ 0.02) but not center,delta, maximum, low or high frequencies for the AFWLA.On the other hand, center (F1,123¼ 11.88, p < 0.01), delta (F1,123¼ 5.82, p ¼ 0.02),high(F1,123¼11.43,p<0.01)frequencies showed significant variation for the SPWLA (Table 1).Pairwise comparison of frequency parameters between the call types for the two species was significant for bandwidth(F3,387¼ 20.85, p < 0.001), center (F3,387¼ 2.972, p ¼ 0.032), high(F3,387¼ 4.03, p ¼ 0.008) and (F3,387¼ 3.864, p ¼ 0.009) frequencies(Table 2).The AFWLA alarm and mobbing calls showed significant difference in bandwidth(ANOVA:F1,265¼5.182,p¼0.02)but not in other call parameters;Table 2).
Mobbing call was more frequently elicited than alarm call in each species of lapwing(AFWLA,χ2(2)¼36.8571,p<0.001;SPWLA,χ2(2)¼18.7463,p<0.001;Fig.3).The rate of alarm and mobbing calls directed to dogs and humans were higher than in cattle,pied crow and other birds for the AFWLA.Mobbing call was more frequently elicited against intruders in the SPWLA.There was significant variation in the antipredatory response call for the two species of lapwings based on category of intruders(AFWLA:χ2(8)¼32.92,p<0.001;SPWLA:χ2(8)¼45.44,p<0.001,Fig.4A and B).
Antipredatory response calls from the two species were elicited within a mean distance of 32.56 17.4 (range, 5–98 m).The alert distance was positively correlated with start distance in the lapwing species(GLMM: p < 0.001; Fig.5).There was significant difference in alert distance between morning and evening of the day in the lapwing species(GLMM:p<0.05;Fig.6).Longer alert distance was recorded for evening than morning.There was no effect of flock size on the call-types made by the two lapwing species.There was variation in call latency by the type of antipredator response call(GLMM:p <0.05).
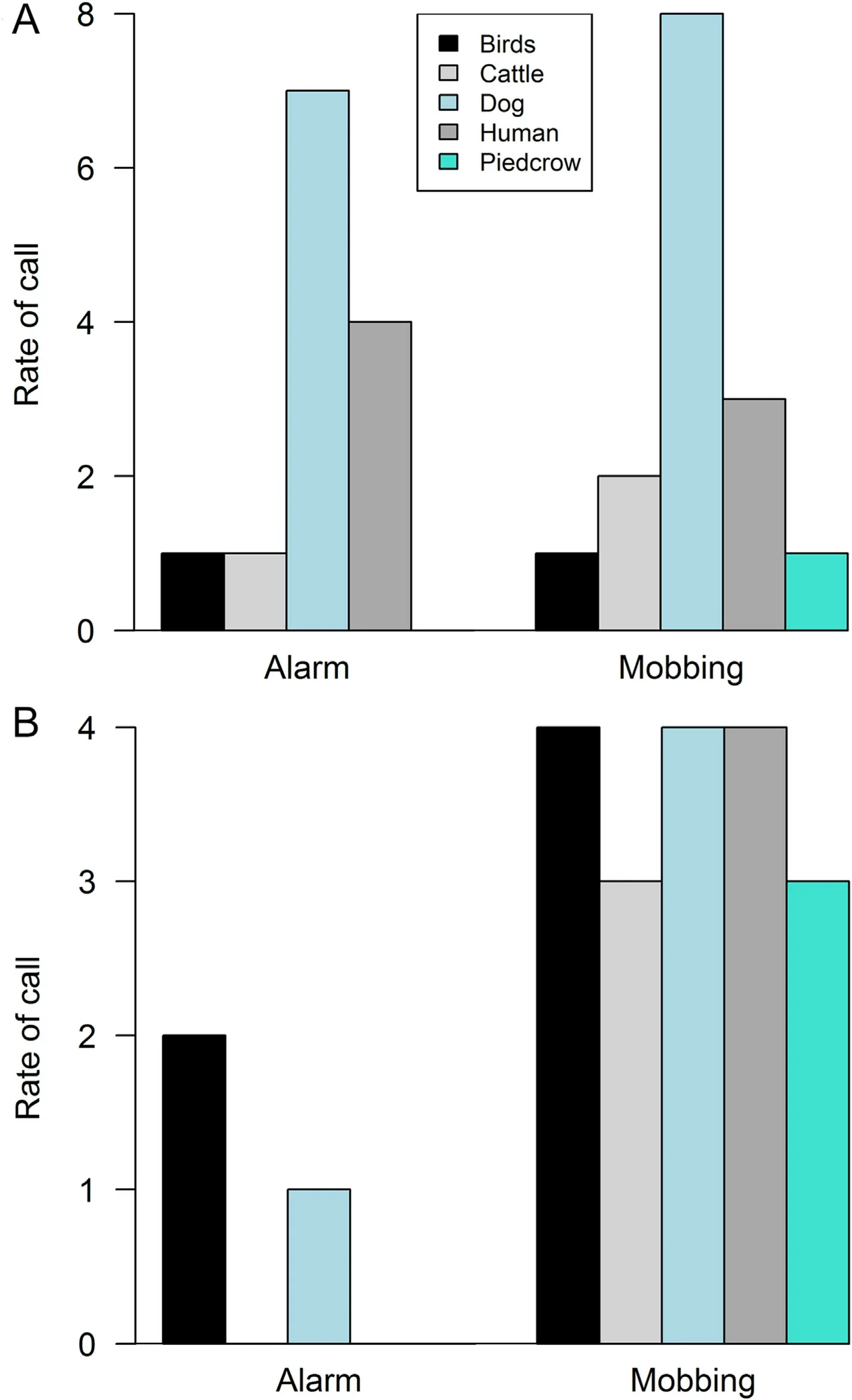
Fig.4.The rate of antipredator call-types(alarm and mobbing)elicited against intruders by the African Wattled Lapwing(A)and Spur-winged Lapwing(B).Dog and human received higher frequency of both alarm and mobbing from the Spur-winged Lapwing.Mobbing call was mainly elicited against intruder in the African Wattle Lapwing.
4.Discussion
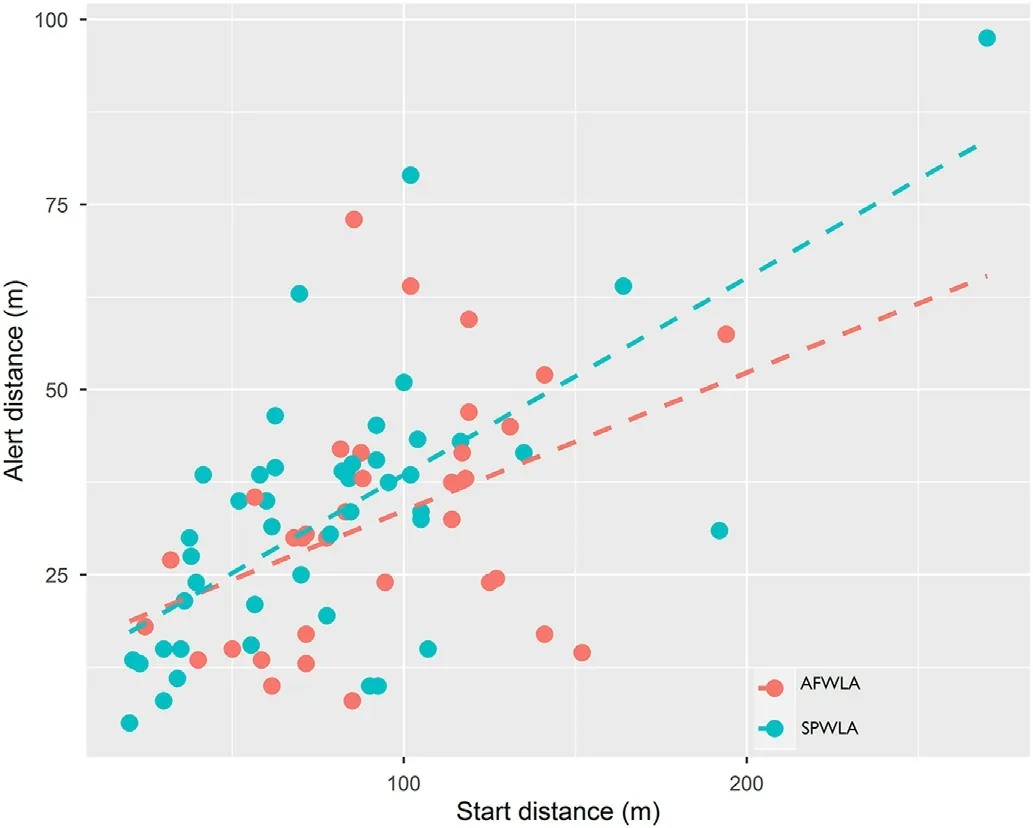
Fig.5.Positive relationship between alert and start distance (m)in the African Wattle Lapwing (red) and Spur-wing Lapwing (blue).
The antipredatory calls (alarm and mobbing) of the lapwing species showed similarity, no clear-cut difference was found in the calls of the species (Fig.2A).Forty-eight percent of the calls were correctly discriminated to call types.Similarities appear stronger within the mobbing and alarm calls of conspecifics.A comparison of the call-based parameters indicates the calls of the species can be differentiated on the basis of maximum and high frequencies(Tables 1 and 2;Figs.1 and 2).AFWLA had longer call duration of lower maximum and high frequencies than SPWLA.Field observation showed that AFWLA elicits longer calls with longer interval between calls than SPWLA (Ramzy Pers.Observ.).The resemblance in the antipredatory calls from the lapwings may stem from both species belonging to the same subfamily Vanellinae with similar behavioral and ecological requirements (del Hoyo et al., 1996).Among birds and other taxonomic groups, interspecific response to distress calls of similar structure has been observed(see Russ et al.,2004;Magrath et al.,2007).The patterns of antipredatory call response of the two lapwings as shown in our result may be influenced by the differential habitat requirement and associated levels of disturbance.Higher rate of alarm and mobbing calls was directed to dogs and humans than in cattle and other birds for the AFWLA, relative to the mobbing call which was more frequently elicited against intruders in the SPWLA.The AFWLA was mostly found around farmlands, this area experiences high disturbance from the presence of human and prowling predators.Whereas the SPWLA was mostly observed around wetland areas with probably less disturbance.AFWLA aggressively mobs predators such as the Yellow-billed Kite (Milvus migrans), Pied Crow (Corvus albus), and Domestic Dog(Canis familiaris), which are known to depredate lapwing eggs (Hart et al.,2002).Humans also destroy eggs or kill adult lapwings(Lees et al.,2013).About 73%of Masked Lapwings(Vanellus vanellus)were known to mob human pedestrians in close proximity to their territories or nesting sites (Cardillini, 2010).Furthermore, AFWLA nests are sometimes destroyed during farmland cultivation.The presence of livestock and cattle grazing leads to nest and egg destruction translating to reproductive failure.These can trigger aggressive defenses displayed towards intruders–both human and non-human(Hart et al.,2002).Our findings fit in the context of Brynychova et al.(2022) who showed how tropical Red-wattled Lapwing (Vanellus indicus aigneri) distinguish between potential predators and adjust their defense strategy accordingly in nest defense.
Latency of mobbing calls was shorter than of alarm calls in both species.Alarm calls convey information to conspecifics about the presence of a predator,but because the behavior of the predator is uncertain,the responses of conspecific prey species vary in latency.In contrast,mobbing calls are made while harassing a potential predator,which may retreat in response.Call latency did not vary by call type played during the playbacks experiment.Lapwings were seen to scan for the source of call before responding; which may imply that the lapwings lack a referential antipredator call system or are more visually oriented than acoustically.Alternatively,experience with incorrect alarm calls of other species within the same habitat may have caused the lapwings to ascertain the source of the call and/or to increase vigilance before eliciting antipredatory behavior(Magrath et al.,2007).Call latency also did not vary by species,perhaps because the two lapwings are close relatives and are sometimes found in mixed flocks.
Flock size has been reported to affect the behavior of individuals in the flock (Maruyama et al., 2009).For example, individuals in larger flocks seem less vigilant than those in smaller flocks.In this study,flock size had no effect on the lapwing call latency.Flock size also had no effect on alert distance.This result is similar to that of Fernandez-Juricic and Jimenez(2001).A plausible explanation is that the flock size of the study species is usually small (3–4 birds).Both species were mostly found in pairs and only few during feeding activities we observe them in flocks.
The distance of speaker from the birds did not affect call latency even though the amplitudes of the calls received by the lapwings varied with distance.Alert distance showed no correlation with distance to cover.Lapwings generally live in open habitats and their proximity to cover(trees or shrubs)does not dictate their response to predators or intruder(Kis et al.,2000;Salek and Cepakova,2006;Borrow and Demey,2014).The species are adapted to fleeing and do not shelter in nearby shrubs and vegetation during escape from predators.
The alert distance of the two lapwings showed no variation with the type of response directed at predators.This implies that the distance at which lapwings detect predators did not affect whether lapwings departed the area or mobbed.However, alert distance was positively correlated with start distance which suggests lapwings may be able to detect predators earlier than when predator initiates approach from a farther distance.This corroborates the report of Fernandez-Juricic and Jimenez(2001).
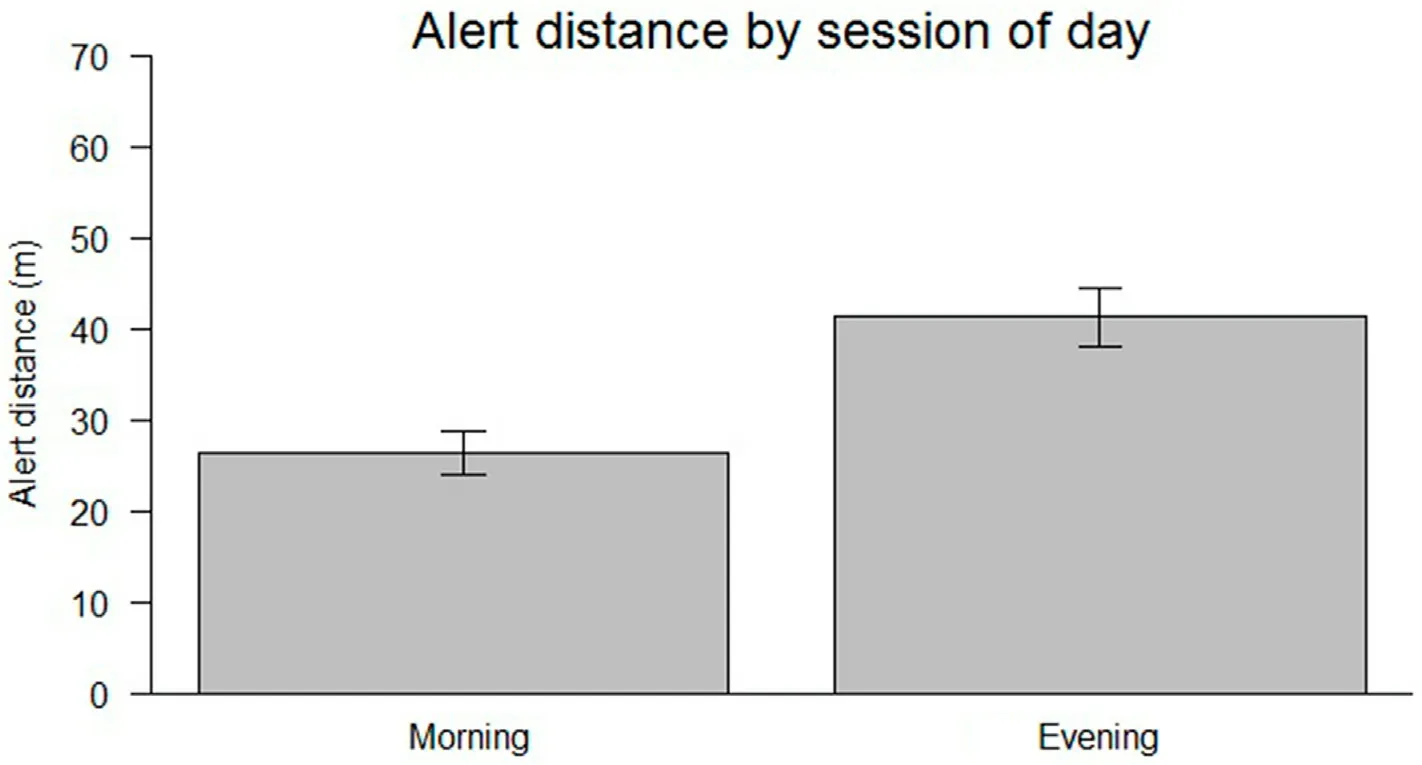
Fig.6.Difference in alert distance by period of the day;alert distance was shorter in the morning relative to longer alert distance in the evening in the lapwing species.
This study showed an effect of time of day on alert distance,shorter alert distance was observed in the morning than evening in both species.Generally, birds seem to be more active in the morning than evening.High foraging activity in the morning may decrease vigilance,hence,the shorter alert distance in the morning relative to the evening.Alternatively,birds that foraged for long hours in the morning may concentrate on being vigilant in the evening.
In summary, the mobbing and alarm calls of both species showed similarity.High frequency and maximum frequency of calls appeared to be important parameters for distinguishing calls of the two lapwing species.The rate of calls of both lapwings varied by species and by the category of intruders.Call latency was a function of the call type and was not affected by flock size,species,and type of intruder playback call.This study provides baseline data and evidence of the antipredatory vocal behavior of two species of lapwing in a tropical African environment.This study was conducted during the breeding period of the lapwings;future studies should focus on non-breeding period to test any contrast in antipredatory behavior in the lapwing species.
Ethics statement
The study did not require any ethical approval as it did not involve direct handling of the species under study at any period.The study was carried out with no form of disturbance to the well-being of the study species.
Authors’contributions
Conceptualization:F.R.J,S.T.O,S.A.M,T.C.O;Formal analysis:F.R.J,S.T.O, C.I.O and T.C.O; Investigation: F.R.J, S.T.O and T.C.O; Methodology: F.R.J, S.T.O, S.A.M and T.C.O; Writing–Original draft: F.R.J and T.C.O;Writing review&editing:all authors;Resource:S.T.O and T.C.O;supervision:S.T.O,S.A.M,T.C.O.All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
The authors wish to thank the management staff,field assistants and students of A.P.Leventis Ornithological Research Institute who provided support during the field work.Our in-depth gratitudes go to two anonymous reviewers for constructive comments on earlier drafts.This project received funding (No.217) from the A.P.Leventis Foundation Scholarship(to F.R.J).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.avrs.2023.100137.
杂志排行

Avian Research的其它文章
- Aviary measurements of dominance and affiliation between members of mixed-species birds flocks in southern China
- Home range variability and philopatry in Cinereous vultures (Aegypius monachus) breeding in Iberia
- A high level of extra-pair paternity in the Chestnut Thrush(Turdus rubrocanus)
- Comparisons of microstructure and elemental composition of eggshells among wild plover populations
- Impact of agricultural landscape structure on the patterns of bird species diversity at a regional scale
- Modelling the nesting-habitat of threatened vulture species in the caucasus:An ecosystem approach to formalising environmental factors in species distribution models